
The Impact of Dietary Curcumin on the Growth Performance, Intestinal Antibacterial Capacity, and Haemato-Biochemical Parameters of Gilthead Seabream (Sparus aurata)

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Diets
2.2. Experimental Design
2.3. Final Sampling
2.4. Blood Sampling and Dissection
2.5. Blood Analysis
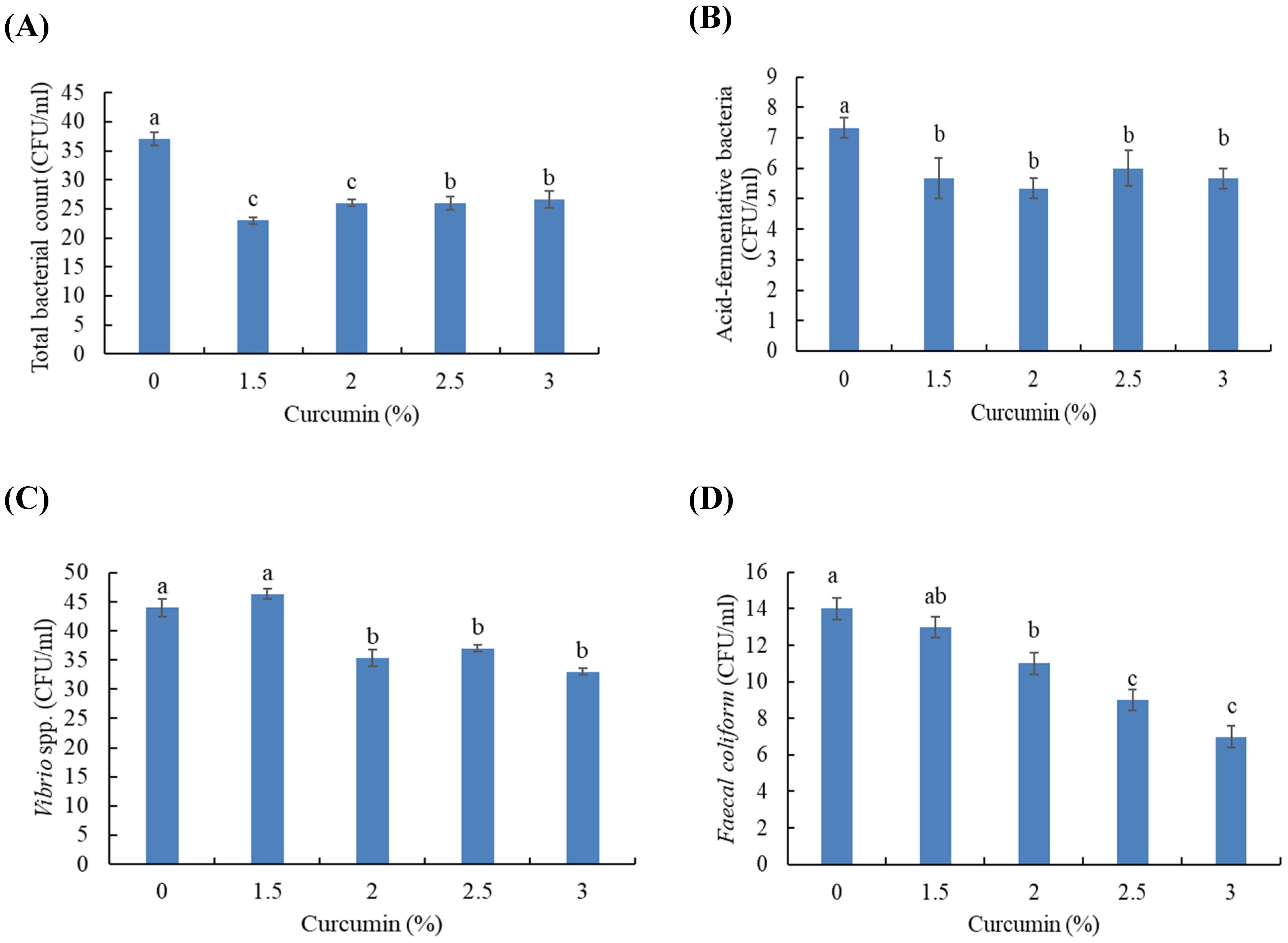
2.6. Intestinal Microbial Analysis
2.7. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Carcass Composition and Somatic Indices
3.3. Intestinal Microbial Populations
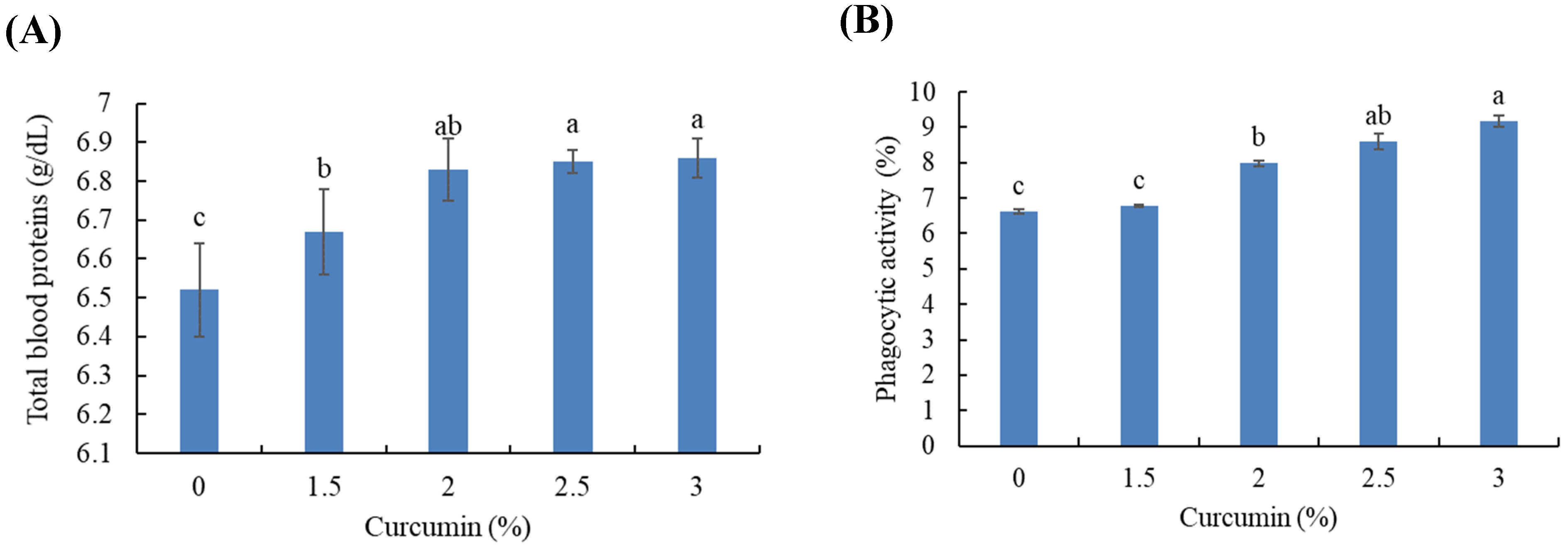
3.4. Hematological and Biochemical Indices
3.5. Blood Immunity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shafique, L.; Abdel-Latif, H.M.R.; Hassan, F.-U.; Alagawany, M.; Naiel, M.A.E.; Dawood, M.A.O.; Yilmaz, S.; Liu, Q. The feasibility of using yellow mealworms (Tenebrio molitor): Towards a sustainable aquafeed industry. Animals 2021, 11, 811. [Google Scholar] [CrossRef]
- Henares, M.N.P.; Medeiros, M.V.; Camargo, A.F.M. Overview of strategies that contribute to the environmental sustainability of pond aquaculture: Rearing systems, residue treatment, and environmental assessment tools. Rev. Aquac. 2020, 12, 453–470. [Google Scholar] [CrossRef]
- El-Hawarry, W.N.; Shourbela, R.M.; Haraz, Y.G.; Khatab, S.A.; Dawood, M.A.O. The influence of carbon source on growth, feed efficiency, and growth-related genes in Nile tilapia (Oreochromis niloticus) reared under biofloc conditions and high stocking density. Aquaculture 2021, 542, 736919. [Google Scholar] [CrossRef]
- Boyd, C.E.; D’Abramo, L.R.; Glencross, B.D.; Huyben, D.C.; Juarez, L.M.; Lockwood, G.S.; McNevin, A.A.; Tacon, A.G.J.; Teletchea, F.; Tomasso, J.R., Jr.; et al. Achieving sustainable aquaculture: Historical and current perspectives and future needs and challenges. J. World Aquac. Soc. 2020, 51, 578–633. [Google Scholar] [CrossRef]
- Mzula, A.; Wambura, P.N.; Mdegela, R.H.; Shirima, G.M. Present status of aquaculture and the challenge of bacterial diseases in freshwater farmed fish in Tanzania; a call for sustainable strategies. Aquac. Fish. 2020, 6, 247–253. [Google Scholar] [CrossRef]
- Chen, J.; Sun, R.; Pan, C.; Sun, Y.; Mai, B.; Li, Q.X. Antibiotics and food safety in aquaculture. J. Agric. Food Chem. 2020, 68, 11908–11919. [Google Scholar] [CrossRef] [PubMed]
- Adel, M.; Dawood, M.A.J.P.B.; Animal, P.M.R.I.; Health, H. Probiotics application: Implications for sustainable aquaculture. Probiotic Bact. Postbiotic Metab. Role Anim. Hum. Health 2021, 2, 191. [Google Scholar]
- EU. Ban on Antibiotics as Growth Promoters in Animal Feed Enters into Effect. Brussels. 92005. 2020. Available online: http://europa.Eu/rapid/press-release_ip-05-1687_en.Htm (accessed on 15 May 2021).
- Perry, W.B.; Lindsay, E.; Payne, C.J.; Brodie, C.; Kazlauskaite, R. The role of the gut microbiome in sustainable teleost aquaculture. Proc. R. Soc. B Biol. Sci. 2020, 287, 20200184. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.C.; Huang, Q.; St-Hilaire, S.; Liu, H.; Zheng, X.; Cheung, K.B.; Zwetsloot, I.M. Fraudulent antibiotic products on the market for aquaculture use. Prev. Vet. Med. 2020, 181, 105052. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; El Basuini, M.F.; Zaineldin, A.I.; Yilmaz, S.; Hasan, M.T.; Ahmadifar, E.; El Asely, A.M.; Abdel-Latif, H.M.R.; Alagawany, M.; Abu-Elala, N.M.; et al. Antiparasitic and antibacterial functionality of essential oils: An alternative approach for sustainable aquaculture. Pathogens 2021, 10, 185. [Google Scholar] [CrossRef]
- Mehrinakhi, Z.; Ahmadifar, E.; Sheikhzadeh, N.; Moghadam, M.S.; Dawood, M.A. Extract of grape seed enhances the growth performance, humoral and mucosal immunity, and resistance of common carp (Cyprinus carpio) against Aeromonas hydrophila. Ann. Anim. Sci. 2021, 21, 217–232. [Google Scholar] [CrossRef]
- Yeganeh, S.; Adel, M.; Nosratimovafagh, A.; Dawood, M.A.O. The effect of Lactococcus lactis subsp. Lactis ptcc 1403 on the growth performance, digestive enzymes activity, antioxidative status, immune response, and disease resistance of rainbow trout (Oncorhynchus mykiss). Probiotics Antimicrob. Proteins 2021. [Google Scholar] [CrossRef] [PubMed]
- Zorofchian, M.S.; Abdul, K.H.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. BioMed Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A review of its effects on human health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Banez, M.J.; Geluz, M.I.; Chandra, A.; Hamdan, T.; Biswas, O.S.; Bryan, N.S.; Von Schwarz, E.R. A systemic review on the antioxidant and anti-inflammatory effects of resveratrol, curcumin, and dietary nitric oxide supplementation on human cardiovascular health. Nutr. Res. 2020, 78, 11–26. [Google Scholar] [CrossRef]
- Liczbiński, P.; Michałowicz, J.; Bukowska, B. Molecular mechanism of curcumin action in signaling pathways: Review of the latest research. Phytother. Res. 2020, 34, 1992–2005. [Google Scholar] [CrossRef] [PubMed]
- Fagnon, M.S.; Thorin, C.; Calvez, S. Meta-analysis of dietary supplementation effect of turmeric and curcumin on growth performance in fish. Rev. Aquac. 2020, 12, 2268–2283. [Google Scholar] [CrossRef]
- Stohs, S.J.; Chen, O.; Ray, S.D.; Ji, J.; Bucci, L.R.; Preuss, H.G. Highly bioavailable forms of curcumin and promising avenues for curcumin-based research and application: A review. Molecules 2020, 25, 1397. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Lázaro, D.; Mielgo-Ayuso, J.; Seco Calvo, J.; Córdova Martínez, A.; Caballero, G.A.; Fernandez-Lazaro, C.I. Modulation of exercise-induced muscle damage, inflammation, and oxidative markers by curcumin supplementation in a physically active population: A systematic review. Nutrients 2020, 12, 501. [Google Scholar] [CrossRef] [Green Version]
- Nm, J.; Joseph, A.; Maliakel, B.; Im, K. Dietary addition of a standardized extract of turmeric (turmafeed(tm)) improves growth performance and carcass quality of broilers. J. Anim. Sci. Technol. 2018, 60, 8. [Google Scholar] [CrossRef] [Green Version]
- Alagawany, M.; Farag, M.R.; Abdelnour, S.A.; Dawood, M.A.O.; Elnesr, S.S.; Dhama, K. Curcumin and its different forms: A review on fish nutrition. Aquaculture 2021, 532, 736030. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Cardinaletti, G.; Cerri, R.; Giorgini, E.; Belloni, A.; Contò, M.; Tibaldi, E.; Olivotto, I. Hermetia illucens and poultry by-product meals as alternatives to plant protein sources in gilthead seabream (Sparus aurata) diet: A multidisciplinary study on fish gut status. Animals 2021, 11, 677. [Google Scholar] [CrossRef] [PubMed]
- Grigorakis, K.; Alexis, M.N.; Taylor, K.D.A.; Hole, M. Comparison of wild and cultured gilthead sea bream (Sparus aurata); composition, appearance and seasonal variations. Int. J. Food Sci. Technol. 2002, 37, 477–484. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- AOAC (Association of Official Analytical Chemists). Official Methods of Analysis of Official Analytical Chemists International, 16th ed.; AOAC: Washington, DC, USA, 1998. [Google Scholar]
- Houston, A. Blood and circulation. In Methods for Fish Biology; American Fisheries Society: Bethesda, MD, USA, 1990. [Google Scholar]
- Blaxhall, P.C.; Daisley, K.W. Routine haematological methods for use with fish blood. J. Fish Biol. 1973, 5, 771–781. [Google Scholar] [CrossRef]
- Dacie, J.V.; Lewis, S.M. Practical Hematology, 6th ed.; Churchill Livingstone: London, UK, 1999. [Google Scholar]
- Doumas, B.T.; Bayse, D.D.; Carter, R.J.; Peters, T.; Schaffer, R. A candidate reference method for determination of total protein in serum. I. Development and validation. Clin. Chem. 1981, 27, 1642–1650. [Google Scholar] [CrossRef]
- Dumas, B.T.; Biggs, H.G. Standard Methods of Clinical Chemistry; Academic Press: New York, NY, USA, 1972. [Google Scholar]
- Reitman, S.; Frankel, S. A colorimeter method for determination of serum glutamic oxaloacetic acid and glutamic pyruvic acid transferases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Kid, P.; King, E. Colorimetric determination of alkaline phosphatase activity. J. Clin. Pathol. 1954, 6, 322. [Google Scholar]
- Schettler, G.; Nussel, N.; Arrbestrred, S.; Praventive, M. Determination of cholesterol in blood. Ann. Clin. Biochem. 1975, 1, 10–25. [Google Scholar]
- Cai, W.-Q.; Li, S.-F.; Ma, J.-Y. Diseases resistance of Nile tilapia (Oreochromis niloticus), blue tilapia (Oreochromis aureus) and their hybrid (female Nile tilapia×male blue tilapia) to Aeromonas sobria. Aquaculture 2004, 229, 79–87. [Google Scholar] [CrossRef]
- Kawahara, E.; Ueda, T.; Nomura, S. In vitro phagocytic activity of white-spotted char blood cells after injection with Aeromonas salmonicida extracellular products. Fish Pathol. 1991, 26, 213–214. [Google Scholar] [CrossRef]
- Zobell, C.E. Marine Microbiology; Chronica Botanica: Waltham, MA, USA, 1946. [Google Scholar]
- Kousoulaki, K.; Sæther, B.S.; Albrektsen, S.; Noble, C. Review on european sea bass (Dicentrarchus labrax, linnaeus, 1758) nutrition and feed management: A practical guide for optimizing feed formulation and farming protocols. Aquac. Nutr. 2015, 21, 129–151. [Google Scholar] [CrossRef]
- Amiza, M.; Zakiah, J.; Ng, L.; Lai, K. Fermentation of tempoyak using isolated tempoyak culture. Res. J. Microbiol. 2010, 5, 903–914. [Google Scholar]
- Paray, B.A.; El-Basuini, M.F.; Alagawany, M.; Albeshr, M.F.; Farah, M.A.; Dawood, M.A.O. Yucca schidigera usage for healthy aquatic animals: Potential roles for sustainability. Animals 2021, 11, 93. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Gewaily, M.S.; Monier, M.N.; Younis, E.M.; Van Doan, H.; Sewilam, H. The regulatory roles of yucca extract on the growth rate, hepato-renal function, histopathological alterations, and immune-related genes in common carp exposed with acute ammonia stress. Aquaculture 2021, 534, 736287. [Google Scholar] [CrossRef]
- Shourbela, R.M.; El-Hawarry, W.N.; Elfadadny, M.R.; Dawood, M.A.O. Oregano essential oil enhanced the growth performance, immunity, and antioxidative status of Nile tilapia (Oreochromis niloticus) reared under intensive systems. Aquaculture 2021, 542, 736868. [Google Scholar] [CrossRef]
- Zhu, F. A review on the application of herbal medicines in the disease control of aquatic animals. Aquaculture 2020, 526, 735422. [Google Scholar] [CrossRef]
- Gholamhosseini, A.; Adel, M.; Dawood, M.A.O.; Banaee, M. The potential benefits of Mentha longifolia on growth performance and innate immunity parameters in caspian kutum (Rutilus frisii kutum). Aquac. Res. 2020, 51, 5212–5227. [Google Scholar] [CrossRef]
- Adel, M.; Omidi, A.H.; Dawood, M.A.O.; Karimi, B.; Shekarabi, S.P.H. Dietary Gracilaria persica mediated the growth performance, fillet colouration, and immune response of Persian sturgeon (Acipenser persicus). Aquaculture 2021, 530, 735950. [Google Scholar] [CrossRef]
- El Asely, A.; Amin, A.; El-Naby, A.S.A.; Samir, F.; El-Ashram, A.; Dawood, M.A.O. Ziziphus mauritiana supplementation of Nile tilapia (Oreochromis niloticus) diet for improvement of immune response to Aeromonas hydrophila infection. Fish Physiol. Biochem. 2020, 46, 1561–1575. [Google Scholar] [CrossRef]
- Mahmoud, H.K.; Al-Sagheer, A.A.; Reda, F.M.; Mahgoub, S.A.; Ayyat, M.S. Dietary curcumin supplement influence on growth, immunity, antioxidant status, and resistance to Aeromonas hydrophila in Oreochromis niloticus. Aquaculture 2017, 475, 16–23. [Google Scholar] [CrossRef]
- Giri, S.S.; Sukumaran, V.; Park, S.C. Effects of bioactive substance from turmeric on growth, skin mucosal immunity and antioxidant factors in common carp, Cyprinus carpio. Fish Shellfish Immunol. 2019, 92, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.; Ye, J.; Zhang, Y.; Xu, Q.; Yang, X.; Shao, X.; Qiang, J.; Xu, P. Optimal dietary curcumin improved growth performance, and modulated innate immunity, antioxidant capacity and related genes expression of nf-κb and nrf2 signaling pathways in grass carp (Ctenopharyngodon idella) after infection with Aeromonas hydrophila. Fish Shellfish Immunol. 2020, 97, 540–553. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Wu, X.-Y.; Zhou, X.-Q.; Feng, L.; Liu, Y.; Jiang, W.-D.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Akdemir, F.; Orhan, C.; Tuzcu, M.; Sahin, N.; Juturu, V.; Sahin, K. The efficacy of dietary curcumin on growth performance, lipid peroxidation and hepatic transcription factors in rainbow trout Oncorhynchus mykiss (walbaum) reared under different stocking densities. Aquac. Res. 2017, 48, 4012–4021. [Google Scholar] [CrossRef]
- Yonar, M.E.; Yonar, S.M.; İspir, Ü.; Ural, M.Ş. Effects of curcumin on haematological values, immunity, antioxidant status and resistance of rainbow trout (Oncorhynchus mykiss) against Aeromonas salmonicida subsp. Achromogenes. Fish Shellfish Immunol. 2019, 89, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.; Xiang, X.; Li, X.; Mai, K.; Ai, Q. Effects of dietary curcumin on growth, antioxidant capacity, fatty acid composition and expression of lipid metabolism-related genes of large yellow croaker fed a high-fat diet. Br. J. Nutr. 2020, 1–10. [Google Scholar] [CrossRef]
- Dawood, M.A.O. Nutritional immunity of fish intestines: Important insights for sustainable aquaculture. Rev. Aquac. 2021, 13, 642–663. [Google Scholar] [CrossRef]
- Li, G.; Zhou, X.; Jiang, W.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.; Tang, L.; Shi, H.; Feng, L. Dietary curcumin supplementation enhanced growth performance, intestinal digestion, and absorption and amino acid transportation abilities in on-growing grass carp (Ctenopharyngodon idella). Aquac. Res. 2020, 51, 4863–4873. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Angulo, C.; Sanchez, V.; Vázquez-Martínez, J.; López, M.G. Antioxidant, intestinal immune status and anti-inflammatory potential of Chenopodium ambrosioides l. in fish: In vitro and in vivo studies. Fish Shellfish Immunol. 2019, 86, 420–428. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Zarrin, V.; Moghadam, E.R.; Hashemi, F.; Makvandi, P.; Samarghandian, S.; Khan, H.; Hashemi, F.; et al. Toward regulatory effects of curcumin on transforming growth factor-beta across different diseases: A review. Front. Pharmacol. 2020, 11, 1785. [Google Scholar] [CrossRef] [PubMed]
- Moniruzzaman, M.; Min, T. Curcumin, curcumin nanoparticles and curcumin nanospheres: A review on their pharmacodynamics based on monogastric farm animal, poultry and fish nutrition. Pharmaceutics 2020, 12, 447. [Google Scholar] [CrossRef]
- Vazirzadeh, A.; Dehghan, F.; Kazemeini, R. Changes in growth, blood immune parameters and expression of immune related genes in rainbow trout (Oncorhynchus mykiss) in response to diet supplemented with Ducrosia anethifolia essential oil. Fish Shellfish Immunol. 2017, 69, 164–172. [Google Scholar] [CrossRef]
- Kundrapu, S.; Noguez, J. Chapter six—laboratory assessment of anemia. In Advances in Clinical Chemistry; Makowski, G.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 83, pp. 197–225. [Google Scholar]
- Fazio, F. Fish hematology analysis as an important tool of aquaculture: A review. Aquaculture 2019, 500, 237–242. [Google Scholar] [CrossRef]
- Enis, Y.M.; Yonar, S.M.; Ural, M.Ş.; Silici, S.; Düşükcan, M. Protective role of propolis in chlorpyrifos-induced changes in the haematological parameters and the oxidative/antioxidative status of Cyprinus carpio carpio. Food Chem. Toxicol. 2012, 50, 2703–2708. [Google Scholar] [CrossRef]
- Wu, L.; Qin, Z.; Liu, H.; Lin, L.; Ye, J.; Li, J. Recent advances on phagocytic b cells in teleost fish. Front. Immunol. 2020, 11, 824. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, R.; Balasundaram, C.; Kim, M.-C.; Kim, J.-S.; Han, Y.-J.; Heo, M.-S. Innate immune response and disease resistance in Carassius auratus by triherbal solvent extracts. Fish Shellfish Immunol. 2009, 27, 508–515. [Google Scholar] [CrossRef]
- Yonar, M.E. Chlorpyrifos-induced biochemical changes in Cyprinus carpio: Ameliorative effect of curcumin. Ecotoxicol. Environ. Saf. 2018, 151, 49–54. [Google Scholar] [CrossRef]
- Jagetia, G.C.; Aggarwal, B.B. “Spicing up” of the immune system by curcumin. J. Clin. Immunol. 2007, 27, 19–35. [Google Scholar] [CrossRef]
- Gao, X.; Kuo, J.; Jiang, H.; Deeb, D.; Liu, Y.; Divine, G.; Chapman, R.A.; Dulchavsky, S.A.; Gautam, S.C. Immunomodulatory activity of curcumin: Suppression of lymphocyte proliferation, development of cell-mediated cytotoxicity, and cytokine production in vitro. Biochem. Pharmacol. 2004, 68, 51–61. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S. Application of fermentation strategy in aquafeed for sustainable aquaculture. Rev. Aquac. 2020, 12, 987–1002. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ingredient | % | Chemical Composition | % |
|---|---|---|---|
| Fish meal | 40 | Dry matter | 90.74 |
| Shrimp meal | 5 | Crude protein | 44.20 |
| Soybean meal | 15 | Ether extract | 15.59 |
| Yellow corn | 10 | Total ash | 9.66 |
| Corn gluten | 6 | Gross energy (Kcal/100 g) | 516.70 |
| Wheat middling | 12 | ||
| Dicalcium phosphate | 1 | ||
| Mineral and vitamin mixture | 1 | ||
| Fish oil | 10 | ||
| Total | 100 |
| Item | Curcumin (%) | ||||
|---|---|---|---|---|---|
| 0 | 1.5 | 2 | 2.5 | 3 | |
| IBW (g) | 20.18 ± 0.51 | 20.15 ± 0.54 | 20.04 ± 0.26 | 19.91 ± 0.12 | 19.74 ± 0.42 |
| FBW (g) | 112.56 ± 0.65 d | 115.20 ± 0.81 cd | 117.13 ± 0.93 bc | 119.30 ± 0.87 b | 126.11 ± 0.38 a |
| WG (g) | 92.38 ± 0.88 d | 95.06 ± 1.23 cd | 97.10 ± 0.96 bc | 99.39 ± 0.76 b | 106.36 ± 0.48 a |
| SGR (%/day) | 1.15 ± 0.02 b | 1.16 ± 0.02 b | 1.18 ± 0.01 b | 1.19 ± 0.00 b | 1.24 ± 0.01 a |
| FCR | 1.61 ± 0.02 b | 1.72 ± 0.01 a | 1.65 ± 0.01 b | 1.56 ± 0.03 c | 1.35 ± 0.01 d |
| PER | 1.41 ± 0.02 bc | 1.32 ± 0.01 c | 1.37 ± 0.01 c | 1.45 ± 0.03 b | 1.67 ± 0.01 a |
| Survival (%) | 91.11 ± 5.88 | 95.56 ± 4.44 | 97.78 ± 2.22 | 95.56 ± 4.44 | 100.00 ± 0.00 |
| Item | Curcumin (%) | |||||
|---|---|---|---|---|---|---|
| Initial | 0 | 1.5 | 2 | 2.5 | 3 | |
| Dry matter (%) | 32.84 ± 1.28 | 33.17 ± 0.03 | 33.54 ± 0.44 | 32.68 ± 0.26 | 34.48 ± 0.36 | 33.23 ± 0.31 |
| Crude protein (%) | 57.28 ± 0.39 | 54.14 ± 0.14 | 53.90 ± 0.34 | 53.35 ± 0.38 | 53.55 ± 0.33 | 54.43 ± 0.24 |
| Lipids (%) | 25.78 ± 0.78 | 28.63± 0.25 | 28.45± 0.13 | 28.23 ± 0.14 | 28.39 ± 0.27 | 28.22 ± 0.55 |
| Ash (%) | 15.80 ± 0.15 | 15.94 ± 0.69 | 16.52 ± 0.20 | 17.16 ± 0.32 | 17.27 ± 0.15 | 16.74 ± 0.40 |
| HSI (%) | - | 2.18 ± 0.08 | 2.47 b ± 0.06 | 2.61 ± 0.17 | 2.70 ± 0.20 | 3.12 ± 0.07 |
| VSI (%) | - | 9.73 ± 0.39 | 10.02 ± 0.30 | 10.54 ± 0.22 | 9.78 ± 0.45 | 9.96 ± 0.31 |
| Item | Curcumin (%) | ||||
|---|---|---|---|---|---|
| 0 | 1.5 | 2 | 2.5 | 3 | |
| Hemoglobin (g/100 mL) | 8.35 ± 0.55 c | 10.29 ± 0.30 b | 11.21 ± 0.65 ab | 12.16 ± 0.35 a | 12.54 ± 0.31 a |
| RBCs (×106/mm3) | 3.56 ± 0.12 b | 3.64 ± 0.12 b | 4.20 ± 0.24 a | 4.46 ± 0.06 a | 4.63 ± 0.03 a |
| Hematocrit (%) | 31.05 ± 0.36 d | 33.20 ± 0.79 c | 37.81 ± 1.09 b | 39.78 ± 0.51 ab | 40.95 ± 0.17 a |
| MCV (µm3/cell) | 99.58 ± 0.39 | 102.96 ± 1.66 | 104.69 ± 0.44 | 105.29 ± 0.27 | 105.54 ± 0.30 |
| MCH (pg/cell) | 30.21 ± 0.39 | 31.41 ± 0.28 | 32.06 ± 0.14 | 32.26 ± 0.25 | 32.90 ± 0.24 |
| MCHC (%) | 30.17 ± 0.15 | 30.62 ± 0.17 | 31.38 ± 0.25 | 31.52 ± 0.37 | 31.80 ± 0.09 |
| WBCs (×103/mm3) | 25.85 ± 89.75 d | 26.64 ± 150.89 c | 27.78 ± 116.93 b | 27.92 ± 33.48 b | 28.57± 136.68 a |
| Lymphocyte (%) | 40.88 ± 0.06 | 41.38 ± 0.51 | 41.72 ± 0.46 | 42.22 ± 0.29 | 42.53 ± 0.28 |
| Monocyte (%) | 4.44 ± 0.31 | 4.48 ± 0.32 | 4.87 ± 0.05 | 5.11 ± 0.17 | 5.35 ± 0.07 |
| Eosinophil (%) | 0.81 ± 0.04 | 0.84 ± 0.01 | 0.93 ± 0.02 | 1.71 ± 0.07 | 1.80 ± 0.04 |
| Item | Curcumin (%) | ||||
|---|---|---|---|---|---|
| 0 | 1.5 | 2 | 2.5 | 3 | |
| ALT (U/I) | 83.76 ± 0.12 | 84.49 ± 0.11 | 84.84c ± 0.03 | 85.51 ± 0.15 | 86.08 ± 0.26 |
| AST (U/I) | 81.33 ± 0.33 | 82.00 ± 0.00 | 82.33 ± 0.33 | 82.67 ± 0.33 | 83.00 ± 0.00 |
| ALP (U/I) | 91.30 ± 1.45 | 70.27 ± 0.66 | 69.55 ± 0.94 | 72.73 ± 1.11 | 61.30 ± 1.11 |
| Albumin (g/dL) | 3.23 ± 0.07 | 3.41 ± 0.08 | 3.65 ± 0.05 | 3.75 ± 0.06 | 3.83 ± 0.06 |
| Urea (mg/dL) | 4.53 ± 0.07 | 4.74 ± 0.05 | 4.76 ± 0.10 | 4.86 ± 0.04 | 5.44 ± 0.06 |
| Total cholesterol (mg/dL) | 971.00 ± 2.31 | 953.00 ± 4.93 | 940.00 ± 3.21 | 941.33 ± 1.45 | 959.00 ± 2.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashry, A.M.; Hassan, A.M.; Habiba, M.M.; El-Zayat, A.; El-Sharnouby, M.E.; Sewilam, H.; Dawood, M.A.O. The Impact of Dietary Curcumin on the Growth Performance, Intestinal Antibacterial Capacity, and Haemato-Biochemical Parameters of Gilthead Seabream (Sparus aurata). Animals 2021, 11, 1779. https://doi.org/10.3390/ani11061779
Ashry AM, Hassan AM, Habiba MM, El-Zayat A, El-Sharnouby ME, Sewilam H, Dawood MAO. The Impact of Dietary Curcumin on the Growth Performance, Intestinal Antibacterial Capacity, and Haemato-Biochemical Parameters of Gilthead Seabream (Sparus aurata). Animals. 2021; 11(6):1779. https://doi.org/10.3390/ani11061779
Chicago/Turabian StyleAshry, Ahmed M., Aziza M. Hassan, Mahmoud M. Habiba, Ahmed El-Zayat, Mohamed E. El-Sharnouby, Hani Sewilam, and Mahmoud A.O. Dawood. 2021. "The Impact of Dietary Curcumin on the Growth Performance, Intestinal Antibacterial Capacity, and Haemato-Biochemical Parameters of Gilthead Seabream (Sparus aurata)" Animals 11, no. 6: 1779. https://doi.org/10.3390/ani11061779
APA StyleAshry, A. M., Hassan, A. M., Habiba, M. M., El-Zayat, A., El-Sharnouby, M. E., Sewilam, H., & Dawood, M. A. O. (2021). The Impact of Dietary Curcumin on the Growth Performance, Intestinal Antibacterial Capacity, and Haemato-Biochemical Parameters of Gilthead Seabream (Sparus aurata). Animals, 11(6), 1779. https://doi.org/10.3390/ani11061779