
The Effect of Egg Laying on Feather and Plasma Corticosterone Concentrations in Turkey (Meleagris gallopavo) Hens

 ,
,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
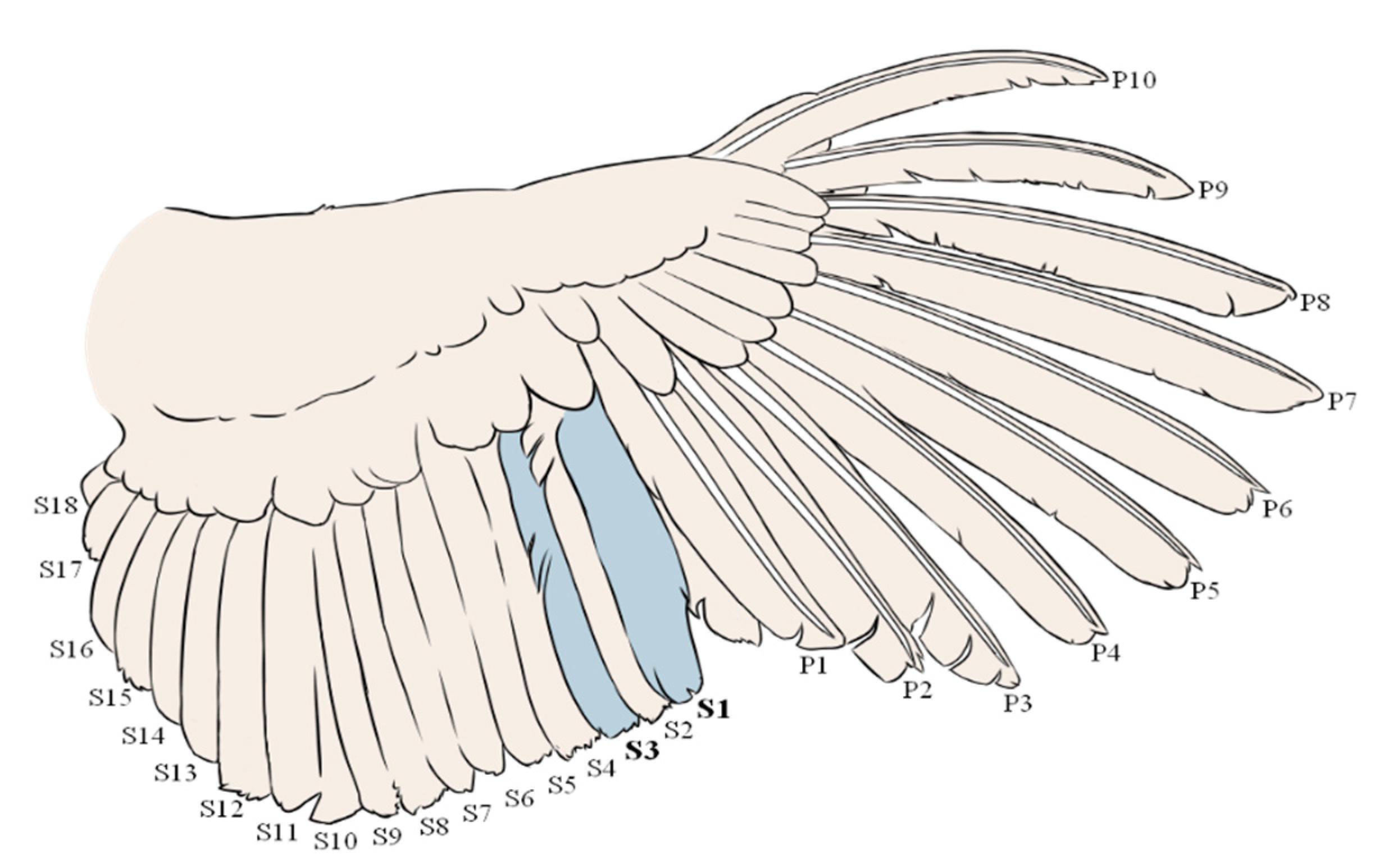
2.1. Subjects and Experimental Design
2.2. FCORT Extraction
2.3. PCORT Extraction
2.4. Assay Evaluation and Procedure
2.5. Statistical Methods
3. Results
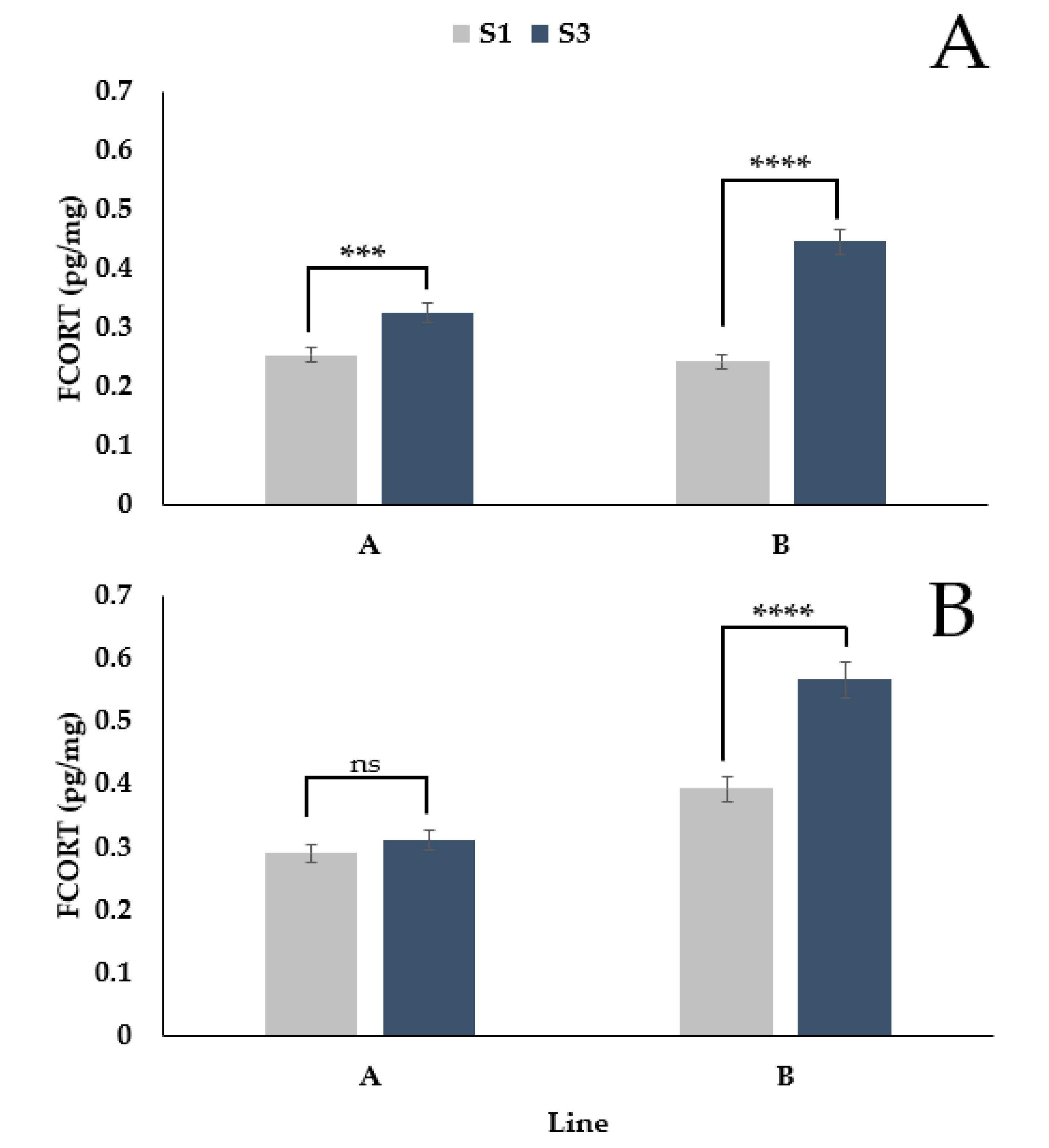
3.1. The Effect of Egg Laying on FCORT Is Dependent on Genetic Line
3.2. Differences in FCORT between Feathers
3.3. Intra- and Inter-Individual Variation in FCORT
3.4. PCORT Decreases during Egg Laying
3.5. Correlations between FCORT and PCORT
4. Discussion
4.1. Effect of Egg Laying on FCORT and PCORT
4.2. FCORT Variability
4.3. Relationship between FCORT and PCORT
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boonstra, R. Coping with Changing Northern Environments: The Role of the Stress Axis in Birds and Mammals. Integr. Comp. Biol. 2004, 44, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Blas, J. Stress in Birds. In Sturkie’s Avian Physiology; Scanes, C.G., Ed.; Elsevier: London, UK, 2015; pp. 769–810. [Google Scholar]
- MacDougall-Shackleton, S.A.; Bonier, F.; Romero, L.M.; Moore, I.T. Glucocorticoids and “Stress” Are Not Synonymous. Integr. Org. Biol. 2019, 1, obz017. [Google Scholar] [CrossRef] [Green Version]
- Scanes, C.G. Biology of stress in poultry with emphasis on glucocorticoids and the heterophil to lymphocyte ratio. Poult. Sci. 2016, 95, 2208–2215. [Google Scholar] [CrossRef]
- Dickens, M.J.; Romero, L.M. A consensus endocrine profile for chronically stressed wild animals does not exist. Gen. Comp. Endocrinol. 2013, 191, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Welcker, J.; Speakman, J.R.; Elliott, K.H.; Hatch, S.A.; Kitaysky, A.S. Resting and daily energy expenditures during reproduction are adjusted in opposite directions in free-living birds. Funct. Ecol. 2014, 29, 250–258. [Google Scholar] [CrossRef] [Green Version]
- Carsia, R.V. Adrenals. In Sturkie’s Avian Physiology; Elsevier BV: Amsterdam, The Netherlands, 2015; pp. 577–611. [Google Scholar]
- Hazard, D.; Couty, M.; Faure, J.M.; Guemene, D. Relationship between hypothalamic-pituitary-adrenal axis responsiveness and age, sexual maturity status, and sex in Japanese quail selected for long or short duration of tonic immobility. Poult. Sci. 2005, 84, 1913–1919. [Google Scholar] [CrossRef]
- Nordquist, R.E.; Zeinstra, E.C.; Dougherty, A.; Riber, A.B. Effects of Dark Brooder Rearing and Age on Hypothalamic Vasotocin and Feather Corticosterone Levels in Laying Hens. Front. Veter Sci. 2020, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lendvai, Á.Z.; Giraudeau, M.; Chastel, O. Reproduction and modulation of the stress response: An experimental test in the house sparrow. Proc. R. Soc. B Boil. Sci. 2006, 274, 391–397. [Google Scholar] [CrossRef] [Green Version]
- Holberton, R.L.; Wingfield, J.C. Modulating the Corticosterone Stress Response: A Mechanism for Balancing Individual Risk and Reproductive Success in Arctic-Breeding Sparrows? Auk 2003, 120, 1140–1150. [Google Scholar] [CrossRef]
- Schmid, B.; Tam-Dafond, L.; Jenni-Eiermann, S.; Arlettaz, R.; Schaub, M.; Jenni, L. Modulation of the adrenocortical response to acute stress with respect to brood value, reproductive success and survival in the Eurasian hoopoe. Oecologia 2013, 173, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, J.C.; O’Reilly, K.M.; Astheimer, L.B.; Wingfield, J.C.; O’Reilly, K.M.; Astheimer, L.B. Modulation of the Adrenocortical Responses to Acute Stress in Arctic Birds: A Possible Ecological Basis. Am. Zool. 1995, 35, 285–294. [Google Scholar] [CrossRef]
- Bortolotti, G.R.; Marchant, T.A.; Blas, J.; German, T. Corticosterone in feathers is a long-term, integrated measure of avian stress physiology. Funct. Ecol. 2008, 22, 494–500. [Google Scholar] [CrossRef]
- Jenni-Eiermann, S.; Helfenstein, F.; Vallat, A.; Glauser, G.; Jenni, L. Corticosterone: Effects on feather quality and deposition into feathers. Methods Ecol. Evol. 2015, 6, 237–246. [Google Scholar] [CrossRef]
- Aharon-Rotman, Y.; Buttemer, W.A.; Koren, L.; Wynne-Edwards, K. Experimental corticosterone manipulation increases mature feather corticosterone content: Implications for inferring avian stress history from feather analyses. bioRxiv 2021. [Google Scholar] [CrossRef]
- Fairhurst, G.; Marchant, T.A.; Soos, C.; Machin, K.L.; Clark, R.G. Experimental relationships between levels of corticosterone in plasma and feathers in a free-living bird. J. Exp. Biol. 2013, 216, 4071–4081. [Google Scholar] [CrossRef] [Green Version]
- Harris, C.M.; Madliger, C.L.; Love, O.P. An evaluation of feather corticosterone as a biomarker of fitness and an ecologically relevant stressor during breeding in the wild. Oecologia 2017, 183, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Romero, L.M.; Fairhurst, G. Measuring corticosterone in feathers: Strengths, limitations, and suggestions for the future. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 202, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattin, C.R.; Reed, J.M.; DesRochers, D.W.; Romero, L.M. Elevated corticosterone in feathers correlates with corticosterone-induced decreased feather quality: A validation study. J. Avian Biol. 2011, 42, 247–252. [Google Scholar] [CrossRef]
- Fusani, L. Endocrinology in field studies: Problems and solutions for the experimental design. Gen. Comp. Endocrinol. 2008, 157, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Leishman, E.M.; Freeman, N.E.; Newman, A.E.; van Staaveren, N.; Wood, B.J.; Harlander-Matauschek, A.; Baes, C.F. Research Note: Quantifying corticosterone in turkey (Meleagris gallopavo) feathers using ELISA. Poult. Sci. 2020, 99, 5261–5264. [Google Scholar] [CrossRef] [PubMed]
- Häffelin, K.E.; Kaufmann, F.; Lindenwald, R.; Döhring, S.; Spindler, B.; Preisinger, R.; Rautenschlein, S.; Kemper, N.; Andersson, R. Corticosterone in feathers: Inter- and intraindividual variation in pullets and the importance of the feather type. Veter Anim. Sci. 2021, 11, 100155. [Google Scholar] [CrossRef] [PubMed]
- Häffelin, K.; Lindenwald, R.; Kaufmann, F.; Döhring, S.; Spindler, B.; Preisinger, R.; Rautenschlein, S.; Kemper, N.; Andersson, R. Corticosterone in feathers of laying hens: An assay validation for evidence-based assessment of animal welfare. Poult. Sci. 2020, 99, 4685–4694. [Google Scholar] [CrossRef]
- Hybrid Turkeys Technical guide for Hybrid Turkeys Commercial Products; Hybrid Turkeys: Kitchener, ON, Canada, 2020.
- Hybrid Turkeys Nutrient Guidelines; Hybrid Turkeys: Kitchener, ON, Canada, 2016.
- Hybrid Turkeys Female Parent Stock Lighting Guidelines; Hybrid Turkeys: Kitchener, ON, Canada, 2015.
- Freeman, N.E.; Newman, A.E.M. Quantifying corticosterone in feathers: Validations for an emerging technique. Conserv. Physiol. 2018, 6, coy051. [Google Scholar] [CrossRef]
- Von Eugen, K.; Nordquist, R.E.; Zeinstra, E.; Van Der Staay, F.J. Stocking Density Affects Stress and Anxious Behavior in the Laying Hen Chick During Rearing. Animals 2019, 9, 53. [Google Scholar] [CrossRef] [Green Version]
- Sapolsky, R.M.; Romero, L.M.; Munck, A.U. How Do Glucocorticoids Influence Stress Responses? Integrating Permissive, Suppressive, Stimulatory, and Preparative Actions. Endocr. Rev. 2000, 21, 55–89. [Google Scholar] [CrossRef] [Green Version]
- Melnychuk, V.L.; Robinson, F.E.; Renema, R.A.; Hardin, R.T.; Emmerson, D.A.; Bagley, L.G. Carcass traits and reproductive development at the onset of lay in two lines of female turkeys. Poult. Sci. 1997, 76, 1197–1204. [Google Scholar] [CrossRef]
- Wang, X.; Liu, L.; Zhao, J.; Jiao, H.; Lin, H. Stress impairs the reproduction of laying hens: An involvement of energy. World’s Poult. Sci. J. 2017, 73, 845–856. [Google Scholar] [CrossRef]
- Hau, M.; Ricklefs, R.E.; Wikelski, M.; Lee, K.A.; Brawn, J.D. Corticosterone, testosterone and life-history strategies of birds. Proc. R. Soc. B Boil. Sci. 2010, 277, 3203–3212. [Google Scholar] [CrossRef]
- Romero, L.M.; Dickens, M.; Cyr, N.E. The reactive scope model—A new model integrating homeostasis, allostasis, and stress. Horm. Behav. 2009, 55, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Sahin, A.; Forbes, J.M. Effect of corticosterone on diet selection for protein in relation to egg production by laying hens. Br. Poult. Sci. 1999, 40, 54–55. [Google Scholar] [CrossRef] [PubMed]
- Shini, S.; Shini, A.; Huff, G. Effects of chronic and repeated corticosterone administration in rearing chickens on physiology, the onset of lay and egg production of hens. Physiol. Behav. 2009, 98, 73–77. [Google Scholar] [CrossRef]
- Wingfield, J.C.; Sapolsky, R.M. Reproduction and Resistance to Stress: When and How. J. Neuroendocr. 2003, 15, 711–724. [Google Scholar] [CrossRef]
- Pereyra, M.E.; Wingfield, J.C. Changes in plasma corticosterone and adrenocortical response to stress during the breeding cycle in high altitude flycatchers. Gen. Comp. Endocrinol. 2003, 130, 222–231. [Google Scholar] [CrossRef]
- Proudman, J. Daily rhythm of prolactin and corticosterone in unrestrained, incubating turkey hens. Domest. Anim. Endocrinol. 1991, 8, 265–270. [Google Scholar] [CrossRef]
- Harris, C.M.; Madliger, C.L.; Love, O.P. Temporal overlap and repeatability of feather corticosterone levels: Practical considerations for use as a biomarker. Conserv. Physiol. 2016, 4, cow051. [Google Scholar] [CrossRef] [Green Version]
- Grunst, M.L.; Grunst, A.S.; Parker, C.E.; Romero, L.M.; Rotenberry, J.T. Pigment-specific relationships between feather corticosterone concentrations and sexual coloration. Behav. Ecol. 2014, 26, 706–715. [Google Scholar] [CrossRef] [Green Version]
- Leishman, E.M.; Van Staaveren, N.; McIntyre, D.R.; Mohr, J.; Wood, B.J.; Baes, C.F.; Harlander-Matauschek, A. Describing the growth and molt of modern domestic turkey (Meleagris gallopavo) primary wing feathers. J. Anim. Sci. 2020, 98, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Leopold, A.S. The Molts of Young Wild and Domestic Turkeys. Condor 1943, 45, 133–145. [Google Scholar] [CrossRef]
- Dalton, H.; Wood, B.; Torrey, S. Injurious pecking in domestic turkeys: Development, causes, and potential solutions. World’s Poult. Sci. J. 2013, 69, 865–876. [Google Scholar] [CrossRef]
- Arrazola, A. The Effect of Alternative Feeding Strategies for Broiler Breeder Pullets throughout the Production Cycle; University of Guelph: Guelph, ON, Canada, 2018. [Google Scholar]
- Hõrak, P.; Männiste, M.; Meitern, R.; Sild, E.; Saks, L.; Sepp, T. Dexamethasone inhibits corticosterone deposition in feathers of greenfinches. Gen. Comp. Endocrinol. 2013, 191, 210–214. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | Total | Line A | Line B |
|---|---|---|---|
| Secondary 1 | 73 | 37 | 36 |
| Secondary 3 | 72 | 34 | 38 |
| Plasma | 73 | 37 | 36 |
| Period | Intra-Individual | Inter-Individual (S1) | Inter-Individual (S3) | |||
|---|---|---|---|---|---|---|
| SD | CV | SD | CV | SD | CV | |
| PRE | 0.11 | 32.58 | 0.07 | 25.41 | 0.12 | 29.96 |
| LAY | 0.14 | 26.15 | 0.22 | 55.22 | 0.33 | 64.91 |
| BOTH | 0.12 | 29.37 | 0.17 | 53.48 | 0.25 | 55.75 |
| Variable | FCORT_S1 | FCORT_S3 | PCORT |
|---|---|---|---|
| FCORT_S1 | 0.15 | 0.13 | |
| FCORT_S3 | 0.57 1 | −0.15 | |
| PCORT | 0.50 1 | 0.43 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leishman, E.M.; van Staaveren, N.; Mohr, J.; Wood, B.J.; Freeman, N.E.; Newman, A.E.M.; Harlander-Matauschek, A.; Baes, C.F. The Effect of Egg Laying on Feather and Plasma Corticosterone Concentrations in Turkey (Meleagris gallopavo) Hens. Animals 2021, 11, 1892. https://doi.org/10.3390/ani11071892
Leishman EM, van Staaveren N, Mohr J, Wood BJ, Freeman NE, Newman AEM, Harlander-Matauschek A, Baes CF. The Effect of Egg Laying on Feather and Plasma Corticosterone Concentrations in Turkey (Meleagris gallopavo) Hens. Animals. 2021; 11(7):1892. https://doi.org/10.3390/ani11071892
Chicago/Turabian StyleLeishman, Emily M., Nienke van Staaveren, Jeff Mohr, Benjamin J. Wood, Nikole E. Freeman, Amy E. M. Newman, Alexandra Harlander-Matauschek, and Christine F. Baes. 2021. "The Effect of Egg Laying on Feather and Plasma Corticosterone Concentrations in Turkey (Meleagris gallopavo) Hens" Animals 11, no. 7: 1892. https://doi.org/10.3390/ani11071892
APA StyleLeishman, E. M., van Staaveren, N., Mohr, J., Wood, B. J., Freeman, N. E., Newman, A. E. M., Harlander-Matauschek, A., & Baes, C. F. (2021). The Effect of Egg Laying on Feather and Plasma Corticosterone Concentrations in Turkey (Meleagris gallopavo) Hens. Animals, 11(7), 1892. https://doi.org/10.3390/ani11071892