
Preference and Motivation Tests for Body Tactile Stimulation in Fish


Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Fish Housing
2.2. Tactile Stimulation
2.3. Experimental Design
2.4. Exposure to Tactile Stimulation
2.5. Preference Test
2.6. Motivation Test
2.7. Social Stress Test
2.8. Experimental Details
2.9. Data Analysis
3. Ethical Note
4. Results
4.1. Previous Exposure to Tactile Stimulation
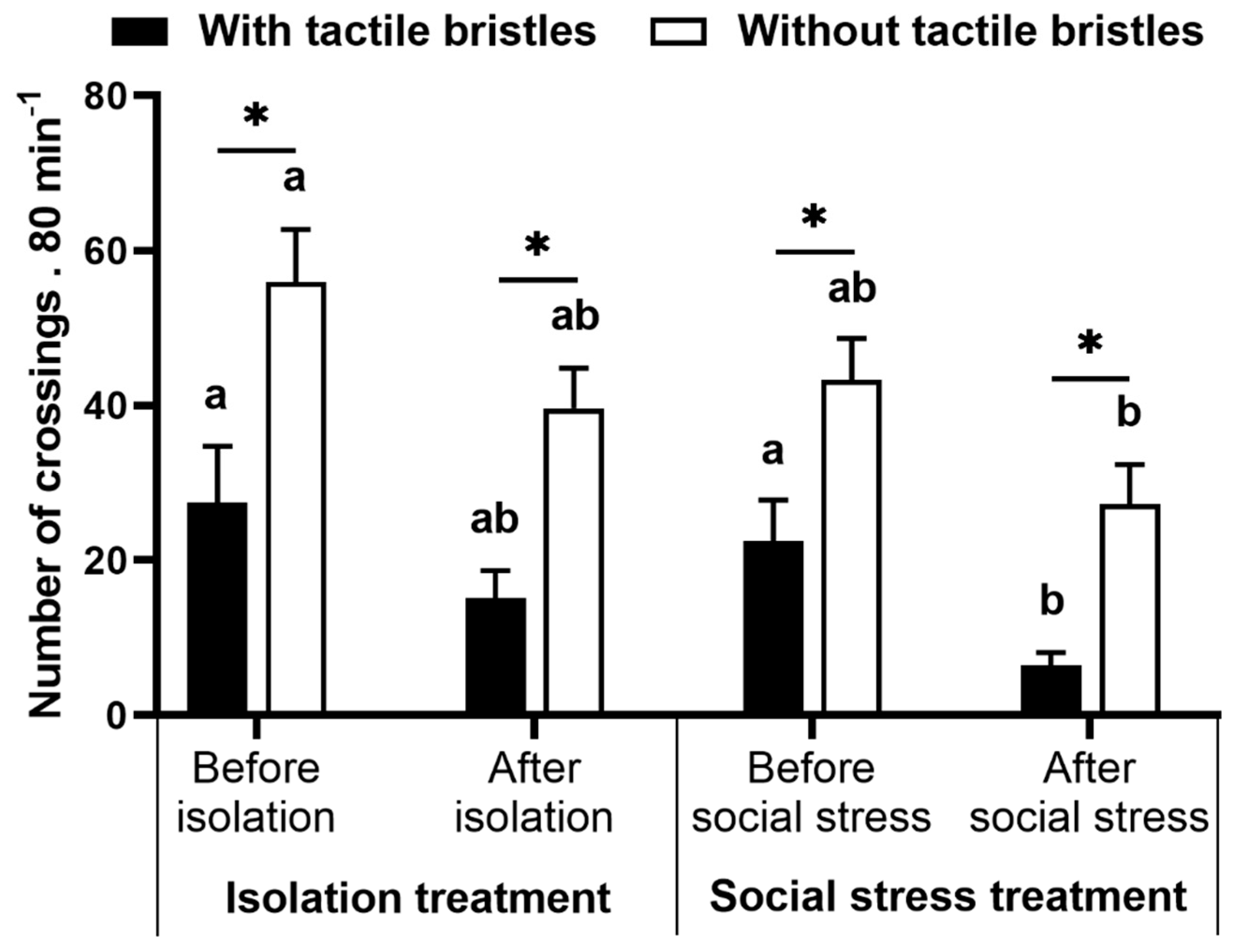
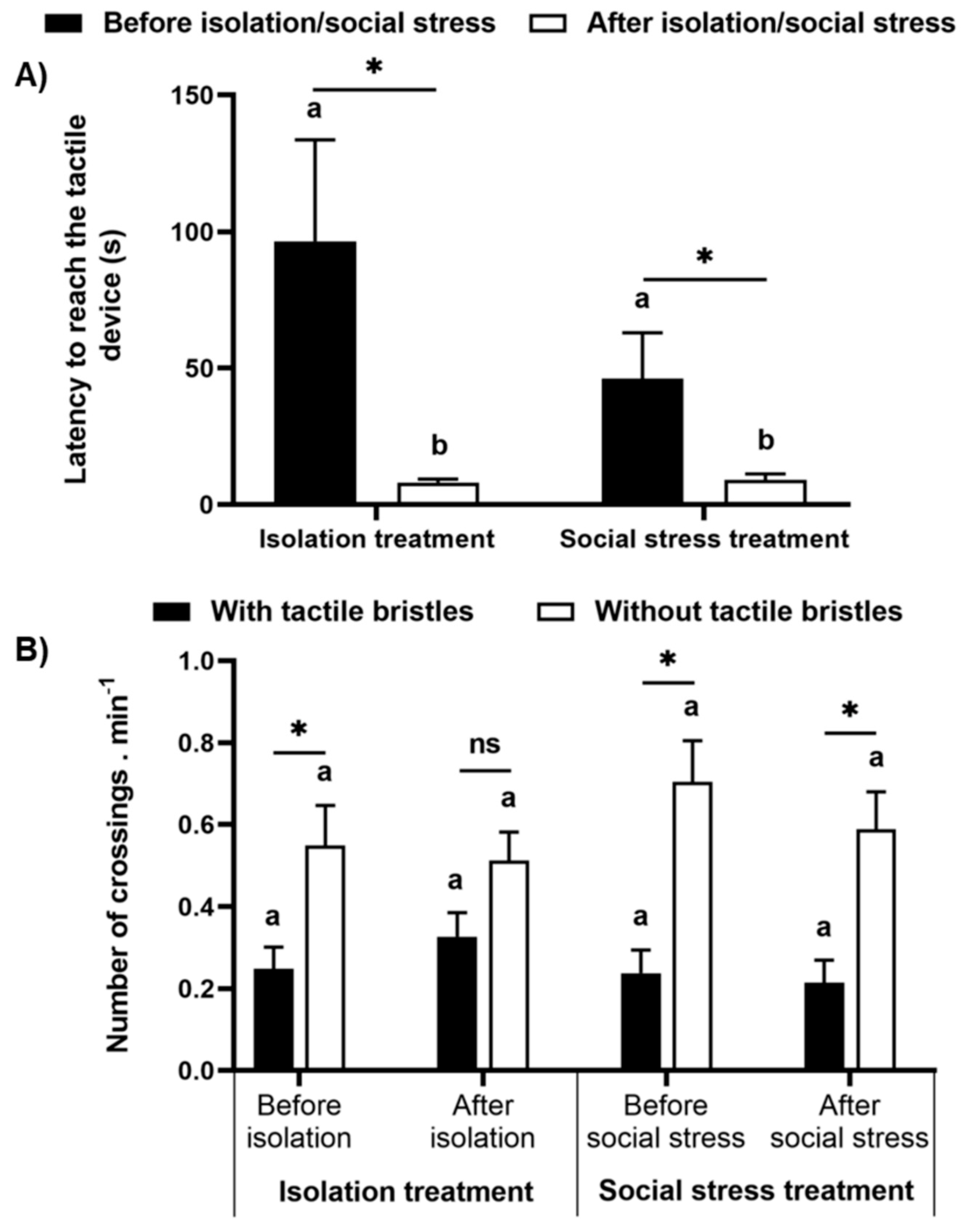
4.2. Preference Test
4.3. Preference Index
4.4. Motivation Test
4.5. Social Rank
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Broom, D.M. A history of animal welfare science. Acta Biotheor. 2011, 59, 121–137. [Google Scholar] [CrossRef]
- Ashley, P.J. Fish welfare: Current issues in aquaculture. Appl. Anim. Behav. Sci. 2007, 104, 199–235. [Google Scholar] [CrossRef]
- Volpato, G.L. Challenges in assessing fish welfare. ILAR J. 2009, 50, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Brown, C. Fish intelligence, sentience and ethics. Anim. Cogn. 2015, 18, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huntingford, F.A.; Kadri, S.; Jobling, M. Introduction: Aquaculture and Behaviour. In Aquaculture and Behavior, 1st ed.; Huntingford, F.A., Jobling, M., Kadri, S., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2012; pp. 1–35. [Google Scholar]
- Huntingford, F.A.; Adams, C.; Braithwaite, V.A.; Kadri, S.; Pottinger, T.G.; Sandøe, P.; Turnbull, J.F. Current issues in fsh welfare. J. Fish. Biol. 2006, 68, 332–372. [Google Scholar] [CrossRef] [Green Version]
- Almazán-Rueda, P.; Schrama, J.W.; Verreth, J.A. Behavioural responses under different feeding methods and light regimes of the African catfish (Clarias gariepinus) juveniles. Aquaculture 2004, 231, 347–359. [Google Scholar] [CrossRef]
- Boscolo, C.N.P.; Morais, R.N.; Gonçalves-de-Freitas, E. Same-sized fish groups increase aggressive interaction of sex-reversed males Nile tilapia GIFT strain. Appl. Anim. Behav. Sci. 2011, 135, 154–159. [Google Scholar] [CrossRef]
- Brandão, M.L.; Braithwaite, V.A.; Gonçalves-de-Freitas, E. Isolation impairs cognition in a social fish. Appl. Anim. Behav. Sci. 2015, 171, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Dawkins, M.S. The science of animal suffering. Ethology 2008, 114, 937–945. [Google Scholar] [CrossRef]
- Volpato, G.L.; Gonçalves-de-Freitas, E.; Fernandes-de-Castilho, M. Insights into the concept of fish welfare. Dis. Aquat. Organ. 2007, 75, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, I.J.H. The changing concept of animal sentience. Appl. Anim. Behav. Sci. 2006, 100, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Kirkden, R.D.; Pajor, E.A. Using preference, motivation and aversion tests to ask scientific questions about animals’ feelings. Appl. Anim. Behav. Sci. 2006, 100, 29–47. [Google Scholar] [CrossRef]
- Nicol, C.J.; Caplen, G.; Edgar, J.; Browne, W.J. Associations between welfare indicators and environmental choice in laying hens. Anim. Behav. 2009, 78, 413–424. [Google Scholar] [CrossRef]
- Galhardo, L.; Almeida, O.; Oliveira, R.F. Measuring motivation in a cichlid fish: An adaption of the push-door paradigm. Appl. Anim. Behav. Sci. 2011, 130, 60–70. [Google Scholar] [CrossRef]
- Franks, B. What do animals want? Anim. Welf. 2019, 28, 1–10. [Google Scholar] [CrossRef]
- Duncan, I.J.H. Measuring preferences and the strength of preferences. Poult. Sci. 1992, 71, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Maia, C.M.; Volpato, G.L. A history-based method to estimate animal preference. Sci. Rep. 2016, 6, 28328. [Google Scholar] [CrossRef] [PubMed]
- Dawkins, M.S. Through animal eyes: What behaviour tell us. Appl. Anim. Behav. Sci. 2006, 100, 4–10. [Google Scholar] [CrossRef]
- Boissy, A.; Arnould, C.; Chaillou, E.; Désiré, L.; Duvaux-Ponter, C.; Greiveldeinger, L.; Leterrier, C.; Richard, S.; Roussel, S.; Saint-Dizier, H.; et al. Emotions and cognition: A new approach to animal welfare. Anim. Welf. 2007, 16, 37–43. [Google Scholar]
- Duncan, I.J.H. Poultry welfare: Science or subjectivity? Poult. Sci. 2002, 43, 643–652. [Google Scholar] [CrossRef]
- Kistler, C.; Heggling, D.; Wurbel, H.; Konig, B. Preference for structured environment in zebrafish (Danio rerio) and checker barbs (Puntius oligolepis). Appl. Anim. Behav. Sci. 2011, 135, 318–327. [Google Scholar] [CrossRef] [Green Version]
- Galhardo, L.; Almeida, O.; Oliveira, R.F. Preference for the presence of substrate in male cichlid fish: Effects of social dominance and context. Appl. Anim. Behav. Sci. 2009, 120, 224–230. [Google Scholar] [CrossRef]
- Mendonça, F.Z.; Volpato, G.L.; Costa-Ferreira, R.S.; Gonçalves-de-Freitas, E. Substratum choice for nesting in male Nile tilapia Oreochromis niloticus. J. Fish. Biol. 2010, 77, 1439–1445. [Google Scholar] [CrossRef]
- Maia, C.M.; Volpato, G.L. Individuality matters for substrate-size preference in the Nile tilapia juveniles. J. Appl. Anim. Welf. Sci. 2018, 21, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Millot, S.; Cerqueira, M.; Castanheira, M.F.; Øverli, Ø.; Martins, C.I.M.; Oliveira, R.F. Use of conditioned place preference/avoidance tests to assess affective states in fish. Appl. Anim. Behav. Sci. 2014, 154, 104–111. [Google Scholar] [CrossRef]
- Maia, C.M.; Ferguson, B.; Volpato, G.L.; Braithwaite, V.A. Physical and psychological motivation tests of individual preferences in rainbow trout. J. Zool. 2017, 302, 108–118. [Google Scholar] [CrossRef]
- Fraser, D.; Nicol, C.J. Preference and motivation research. In Animal Welfare, 3rd ed.; Appleby, M.C., Olsson, I.A.S., Galindo, F., Eds.; CAB International: Wallingford, UK, 2018; pp. 213–231. [Google Scholar]
- Jensen, M.B.; Pedersen, L.J. Using motivation tests to assess ethological needs and preferences. Appl. Anim. Behav. Sci. 2008, 113, 340–356. [Google Scholar] [CrossRef]
- Olsson, I.A.S.; Keeling, L.J.C. The push-door for measuring motivation in hens: Laying hens are motivated to perch at night. Anim. Welf. 2002, 9, 11–19. [Google Scholar]
- Sullivan, M.; Lawrence, C.; Blache, D. Why did the fish cross the tank? Objectively measuring the value of enrichment for captive fish. Appl. Anim. Behav. Sci. 2016, 174, 181–188. [Google Scholar] [CrossRef]
- Gonçalves-de-Freitas, E.; Bolognesi, M.C.; Gauy, A.C.S.; Brandão, M.L.; Giaquinto, P.C.; Fernandes-Castilho, M. Social behavior and welfare in Nile tilapia. Fishes 2019, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Field, T.; Hernandez-Reif, M.; Diego, M.; Schanberg, S.; Kuhn, C. Cortisol decreases and serotonin and dopamine increase following massage therapy. Int. J. Neurosci. 2005, 115, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Reif, M.; Diego, M.; Field, T. Preterm infants show reduced stress behaviors and activity after 5 days of massage therapy. Inf. Behav. Develop. 2007, 30, 557–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Probst, J.K.; Spengler-Nef, A.; Leiber, F.; Kreuzer, M.; Hillmann, E. Gentle touching in early life reduces avoidance distance and slaughter stress in beef cattle. Appl. Anim. Behav. Sci. 2012, 139, 42–49. [Google Scholar] [CrossRef]
- Field, T. Massage therapy effects. Am. Psychol. 1998, 53, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Jane, S.W.; Wilkie, D.J.; Galucci, B.B.; Beaton, R.D.; Huang, H.Y. Effects of a full-body massage on pain intensity, anxiety, and physiological relaxation in taiwanese patients with metastatic bone pain: A pilot study. J. Pain Symptom Manag. 2009, 37, 754–763. [Google Scholar] [CrossRef]
- Freitas, D.; Antoniazzi, C.T.D.; Segat, H.J.; Metz, V.G.; Vey, L.T.; Barcelos, R.C.S.; Duarte, T.; Duarte, M.M.M.F.; Burger, M.E. Neonatal tactile stimulation decreases depression-like and anxiety-like behaviors and potentiates sertraline action in young rats. Int. J. Dev. Neurosci. 2015, 47, 192–197. [Google Scholar] [CrossRef]
- Antoniazzi, C.T.; Metz, V.G.; Roversi, K.; Freitas, D.L.; Vey, L.T.; Dias, V.T.; Segat, H.J.; Duarte, M.M.; Burger, M.E. Tactile stimulation during different developmental periods modifies hippocampal BDNF and GR, affecting memory and behavior in adult rats. Hippocampus 2017, 27, 210–220. [Google Scholar] [CrossRef]
- De Los Angeles, G.A.M.; Del Carmen, R.O.; Wendy, P.M.; Socorro, R.M. Tactile stimulation effects on hippocampal neurogenesis and spatial learning and memory in prenatally stressed rats. Brain Res. Bull. 2016, 124, 1–11. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Soares, M.C.; Oliveira, R.F.; Ros, A.F.H.; Grutter, A.S.; Bshary, R. Tactile stimulation lowers stress in fish. Nat. Commun. 2011, 2, 534. [Google Scholar] [CrossRef] [Green Version]
- Bolognesi, M.C.; Gauy, A.C.S.; Gonçalves-deFreitas, E. Effect of tactile stimulation in a territorial fish. Sci. Rep. 2019, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Alvarenga, C.M.D.; Volpato, G.L. Agonistic profile and metabolism in alevins of the Nile tilapia. Physiol. Behav. 1995, 57, 75–80. [Google Scholar] [CrossRef]
- Carvalho, T.B.; Gonçalves-de-Freitas, E. Sex group composition, social interaction, and metabolism in the fish Nile tilapia. Braz. J. Biol. 2008, 68, 807–812. [Google Scholar] [CrossRef] [Green Version]
- Barreto, R.E.; Carvalho, G.G.A.; Volpato, G.L. The aggressive behavior of Nile tilapia introduced into novel environments with variation in enrichment. Zoology 2011, 114, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Mendl, M.; Burman, O.H.P.; Paul, E.S. An integrative and functional framework for the study of animal emotion and mood. Proc. R. Soc. B 2010, 277, 2895–2904. [Google Scholar] [CrossRef] [Green Version]
- Cerqueira, M.; Millot, S.; Castanheira, M.F.; Félix, A.S.; Silva, T.; Oliveira, G.A.; Oliveira, C.C.; Martins, C.I.M.; Oliveira, R.F. Cognitive appraisal of environmental stimuli induces emotion-like states in fish. Sci. Rep. 2017, 7, 13181. [Google Scholar] [CrossRef]
- Damsgård, B.; Huntingford, F. Fighting and aggression. In Aquaculture and Behavior, 1st ed.; Huntingford, F., Jobling, M., Kadri, S., Eds.; WilleyBlackwell: Oxford, UK, 2012; pp. 248–285. [Google Scholar]
- FAO. The state of world fisheries and aquaculture 2020. In Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2020; p. 206. [Google Scholar] [CrossRef]
- FishEthoBase Project. Available online: http://www.fishethobase.net/ (accessed on 9 May 2021).
- Saraiva, J.L.; Arechavala-Lopez, P.; Castanheira, M.F.; Volstorf, J.; Studer, B.H. A global assessment of welfare in farmed fishes: The FishEthoBase. Fishes 2019, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Conte, F.S. Stress and the welfare of cultured fish. Appl. Anim. Behav. Sci. 2004, 86, 205–223. [Google Scholar] [CrossRef]
- Volpato, G.L.; Giaquinto, P.C.; Castilho, M.F.; Barreto, R.E.; Gonçalves-de-Freitas, E. Animal welfare: From concepts to reality. Oecol. Bras. 2009, 13, 5–15. [Google Scholar] [CrossRef] [Green Version]
- Marchesan, M.; Spoto, M.; Verginella, L.; Ferrero, E.A. Behavioural effects of artificial light on fish species of commercial interest. Fish. Res. 2005, 73, 171–185. [Google Scholar] [CrossRef]
- Mesquita, F.D.; Godinho, H.P.; de Azevedo, P.G.; Young, R.J. A preliminary study into the effectiveness of stroboscopic light as an aversive stimulus for fish. Appl. Anim. Behav. Sci. 2008, 111, 402–407. [Google Scholar] [CrossRef]
- Maximino, C.; Marques, T.; Dias, F.; Cortes, F.V.; Taccolini, I.B.; Pereira, P.M.; Colmanetti, R.; Gazolla, R.A.; Tavares, R.I.; Rodrigues, S.T.K.; et al. A comparative analysis of the preference for dark environments in five teleosts. Int. J. Comp. Psychol. 2007, 20, 351–367. [Google Scholar]
- Ejike, C.; Schreck, C.B. Stress and social hierarchy rank in coho salmon. Trans. Am. Fish. Soc. 1980, 109, 423–426. [Google Scholar] [CrossRef]
- Corrêa, S.A.; Fernandes, M.O.; Iseki, K.K.; Negrao, J.A. Effect of the establishment of dominance relationships on cortisol and other metabolic parameters in Nile tilapia (Oreochromis niloticus). Braz. J. Med. Biol. Res. 2003, 36, 1725–1731. [Google Scholar] [CrossRef] [Green Version]
- Foo, J.T.W.; Lam, T.J. Serum cortisol response to handling stress and the effect of cortisol implantation on testosterone level in the tilapia, Oreochromis mossambicus. Aquaculture 1993, 115, 145–158. [Google Scholar] [CrossRef]
- Barcellos, L.J.G.; Volpato, G.L.; Barreto, R.E.; Coldebella, I.; Ferreira, D. Chemical communication of handling stress in fish. Physiol. Behav. 2011, 103, 372–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves-de-Freitas, E.; Teresa, F.B.; Gomes, F.S.; Giaquinto, P.C. Effect of water renewal on dominance hierarchy of juvenile Nile tilapia. Appl. Anim. Behav. Sci. 2008, 112, 187–195. [Google Scholar] [CrossRef]
- Ros, A.F.H.; Becker, K.; Oliveira, R.F. Aggressive behaviour and energy metabolism in a cichlid fish, Oreochromis mossambicus. Physiol. Behav. 2006, 89, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Barreto, T.N.; Boscolo, C.N.P.; Gonçalves-de-Freitas, E. Homogeneously sized groups increase aggressive interaction and affect social stress in Thai strain Nile tilapia (Oreochromis niloticus). Mar. Freshw. Behav. Physiol. 2015, 48, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Maia, C.M.; Volpato, G.L. Environmental light color affects the stress response of Nile tilapia. Zoology 2013, 116, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Lehner, P.N. Handbook of Ethological Methods, 2nd ed.; Cambridge University Press: Cambridge, MA, USA, 1996; p. 672. [Google Scholar]
- Grutter, A.S. Relationship between changing rates and ectoparasite loads in coral reef fshes. Mar. Ecol. Prog. Ser. 1995, 118, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Huth, A.; Wissel, C. The simulation of the movement of fish schools. J. Theor. Biol. 1992, 156, 365–385. [Google Scholar] [CrossRef]
- Carleton, K.L. Cichlid fish visual systems: Mechanisms of spectral tuning. Integr. Zool. 2009, 4, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.L.S.; Gonçalves-de-Freitas, E.; Volpato, G.L.; Oliveira, C. Visual communication stimulates reproduction in Nile tilapia, Oreochromis niloticus (L.). Braz. J. Med. Biol. Res. 2009, 42, 368–374. [Google Scholar] [CrossRef] [Green Version]
- Douglas, R.H.; Hawryshyn, C.W. Behavioural studies on fish vision: An analysis of visual capabilities. In The Visual System of Fish, 1st ed.; Douglas, R.H., Djamgoz, M.B.A., Eds.; Chapman and Hall: London, UK, 1990; Volume 1, pp. 373–418. [Google Scholar]
- Amaral, S.D.; Branco, P.; Katopodis, C.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. To swim or to jump? Passage behaviour of a potamodromous cyprinid over an experimental broad-crested weir. River Res. Applic. 2017, 34, 174–182. [Google Scholar] [CrossRef]
- Maia, C.M.; Volpato, G.L. Preference index supported by motivation tests in Nile tilapia. PLoS ONE 2017, 12, e0175821. [Google Scholar] [CrossRef] [Green Version]
- Maia, C.M.; Alves, N.P.C.; Tatemoto, P. Juvenile Nile tilapia fish avoid red shelters. J. Appl. Anim. Welf. Sci. 2020, 24, 98–106. [Google Scholar] [CrossRef]
- Carvalho, T.B.; Mendonça, F.Z.; Costa-Ferreira, R.S.; Gonçalves-de-Freitas, E. The effect of increased light intensity on the aggressive behavior of the Nile tilapia, Oreochromis niloticus (Teleostei: Cichlidae). Zoologia 2013, 30, 125–129. [Google Scholar] [CrossRef] [Green Version]
- Brandão, M.L.; Fernandes, A.M.T.A.; Goncalves-de-Freitas, E. Male and female cichlid fish show cognitive inhibitory control ability. Sci. Rep. 2019, 9, 15795. [Google Scholar] [CrossRef] [PubMed]
- Schreck, C.B. Physiological, behavioral, and performance indicators of stress. Am. Fish. Soc. Symp. 1990, 8, 29–37. [Google Scholar]
- Kutty, M.N.; Sukumaran, N. Influence of upper and lower temperature extremes on the swimming performance of Tilapia mossambica. Trans. Am. Fish. Soc. 1975, 104, 755–761. [Google Scholar] [CrossRef]
- Lankford, S.E.; Adams, T.E.; Miller, R.A.; Cech, J.J., Jr. The cost of chronic stress: Impacts of a nonhabituating stress response on metabolic variables and swimming performance in sturgeon. Physiol. Biochem. Zool. 2005, 78, 599–609. [Google Scholar] [CrossRef]
- Gaikwad, S.; Stewart, A.; Hart, P.; Wong, K.; Piet, V.; Cachat, J.; Kalueff, A.V. Acute stress disrupts performance of zebrafish in the cued and spatial memory tests: The utility of fish models to study stress–memory interplay. Behav. Process. 2011, 87, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.S.; Desjardins, J.K.; Fernand, R.D. Effects of stress and motivation on performing a spatial task. Neurobiol. Learn. Mem. 2011, 95, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galhardo, L.; Oliveira, R.F. The effects of social isolation on steroid hormone levels are modulated by previous social status and context in a cichlid fish. Horm. Behav. 2014, 65, 1–5. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Session | Row Frequency | Cumulative Frequency | Area | Cumulative Area | Expected Area | Area Variation | Preference Index (PI) |
|---|---|---|---|---|---|---|---|
| 1 | 2 | 2 | 1 | 1 | 5.25 | −4.25 | −4.25 |
| 2 | 6 | 8 | 9 | 10 | 30.75 | −20.75 | −25 |
| 3 | 8 | 16 | 20 | 30 | 50.75 | −20.75 | −45.75 |
| 4 | 5 | 21 | 17.5 | 47.5 | 80.5 | −33 | −78.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gauy, A.C.d.S.; Bolognesi, M.C.; Martins, G.D.; Gonçalves-de-Freitas, E. Preference and Motivation Tests for Body Tactile Stimulation in Fish. Animals 2021, 11, 2042. https://doi.org/10.3390/ani11072042
Gauy ACdS, Bolognesi MC, Martins GD, Gonçalves-de-Freitas E. Preference and Motivation Tests for Body Tactile Stimulation in Fish. Animals. 2021; 11(7):2042. https://doi.org/10.3390/ani11072042
Chicago/Turabian StyleGauy, Ana Carolina dos Santos, Marcela Cesar Bolognesi, Guilherme Delgado Martins, and Eliane Gonçalves-de-Freitas. 2021. "Preference and Motivation Tests for Body Tactile Stimulation in Fish" Animals 11, no. 7: 2042. https://doi.org/10.3390/ani11072042
APA StyleGauy, A. C. d. S., Bolognesi, M. C., Martins, G. D., & Gonçalves-de-Freitas, E. (2021). Preference and Motivation Tests for Body Tactile Stimulation in Fish. Animals, 11(7), 2042. https://doi.org/10.3390/ani11072042