
Update on Distribution and Conservation Status of Amphibians in the Democratic People’s Republic of Korea: Conclusions Based on Field Surveys, Environmental Modelling, Molecular Analyses and Call Properties

 ,
,  , ,
, ,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxonomic Sampling
2.2. Genetic Identification
2.3. Pictures Identification
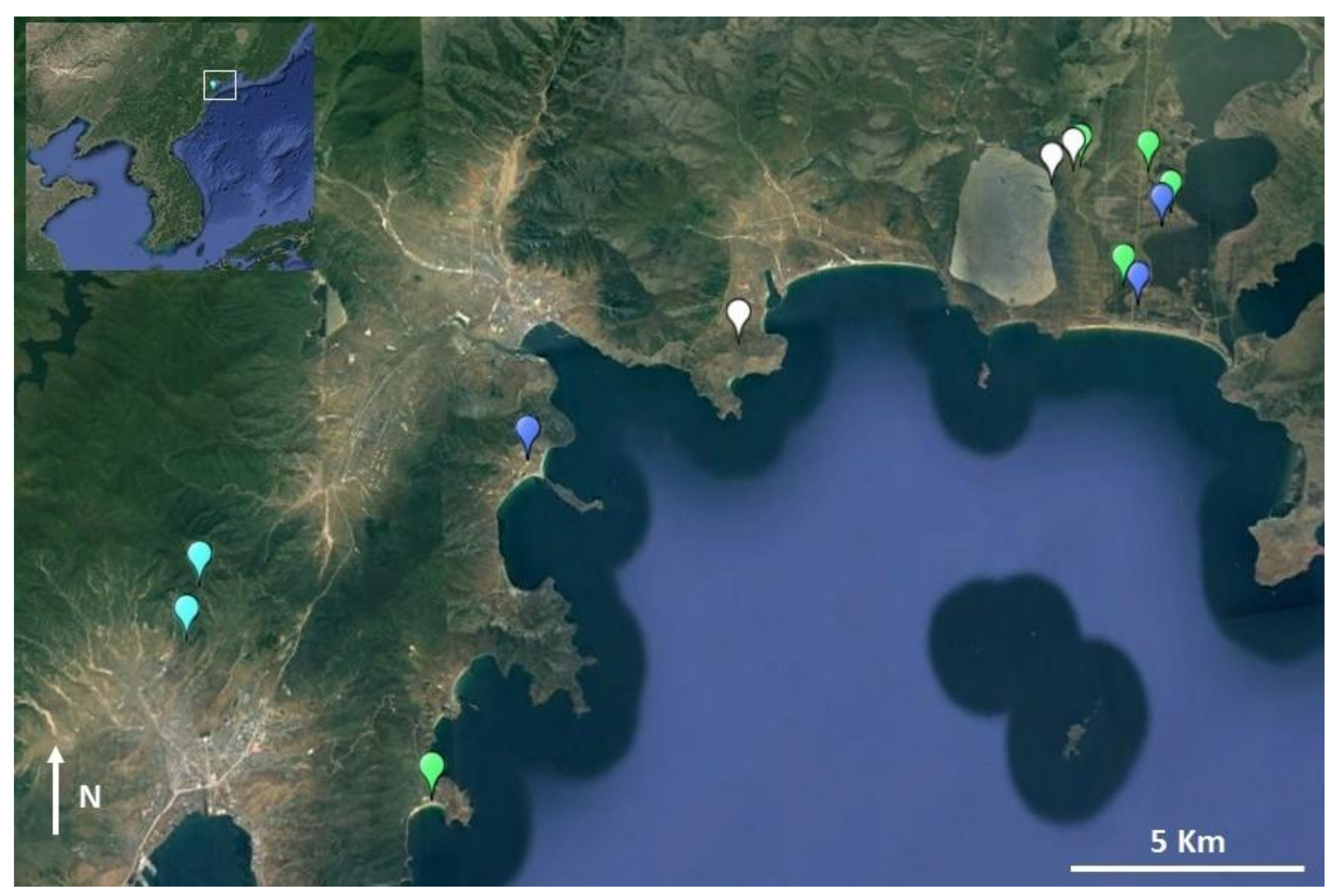
2.4. Field Surveys
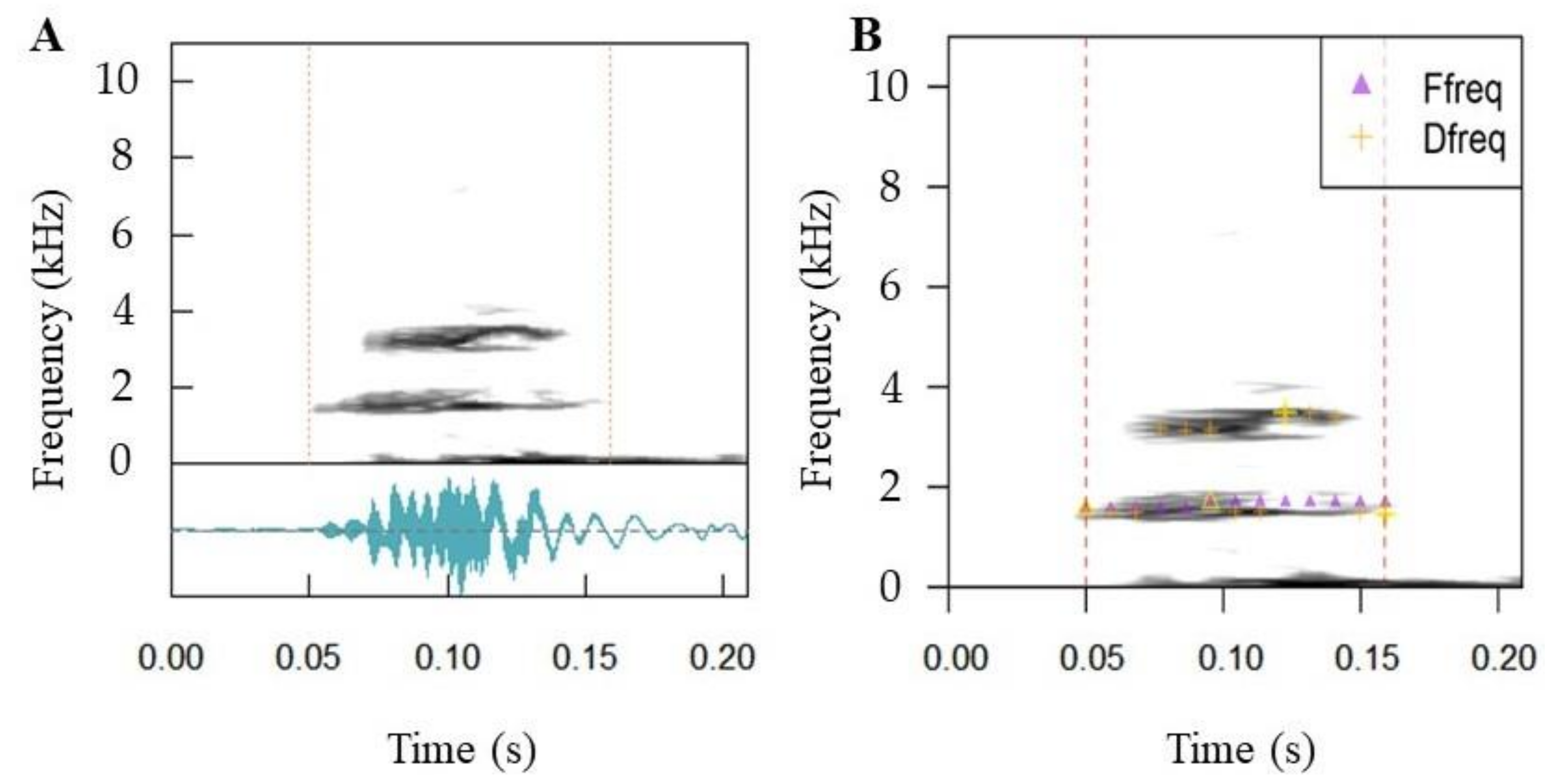
2.5. Call Properties
2.6. Distribution Estimates and Species Richness
2.6.1. Anura
2.6.2. Caudata
2.7. Extinction Risk Assessment
3. Results
3.1. Genetic Identification
3.2. Picture Identification
3.3. Field Surveys
3.4. Call Properties
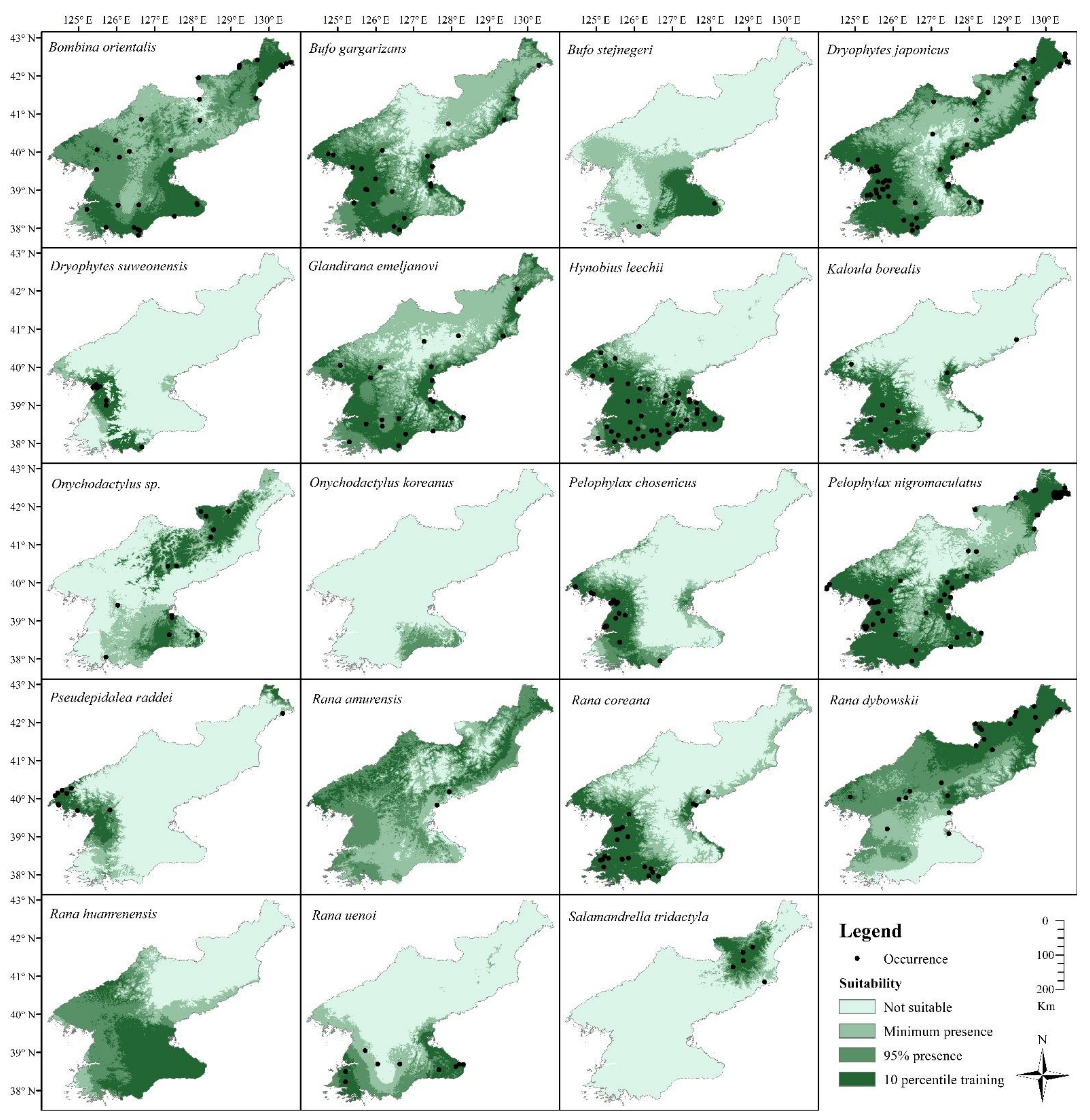
3.5. Distribution Estimates and Species Richness
3.6. Extinction Risk Assessment
3.6.1. Bombina orientalis
3.6.2. Bufo gargarizans
3.6.3. Bufo stejnegeri
3.6.4. Dryophytes japonicus
3.6.5. Dryophytes suweonensis
3.6.6. Glandirana emeljanovi
3.6.7. Hynobius leechii
3.6.8. Kaloula borealis
3.6.9. Onychodactylus sp.
3.6.10. Onychodactylus fischeri
3.6.11. Onychodactylus zhaoermii
3.6.12. Onychodactylus zhangyapingi
3.6.13. Onychodactylus koreanus
3.6.14. Pelophylax chosenicus
3.6.15. Pelophylax nigromaculatus
3.6.16. Strauchbufo raddei
3.6.17. Rana amurensis
3.6.18. Rana coreana
3.6.19. Rana dybowskii
3.6.20. Rana huanrenensis
3.6.21. Rana uenoi
3.6.22. Salamandrella tridactyla
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IUCN. The IUCN Red List of Threatened Species; IUCN, Ed.; Version 2020-1; IUCN: Gland, Switzerland, 2020; Available online: https://www.iucnredlist.org (accessed on 19 March 2020).
- Tomek, T. The birds of North Korea. Non-Passeriformes. Acta Zool. Crac. 1999, 42, 1–21. [Google Scholar]
- Tomek, T. The birds of North Korea. Passeriformes. Acta Zool. Crac. 2002, 45, 1–235. [Google Scholar]
- Moores, N. Birds and their conservation in Rason Special Economic Zone, Democratic People’s Republic of Korea. Forktail 2018, 33, 124–133. [Google Scholar]
- MAB National Committee of DPR Korea. Biodiversity of Mt. Kumgang; UNESCO-MAB Programme: Pyongang, Democratic People’s Republic of Korea, 2003; Volume 92. [Google Scholar]
- Li, M. Research on the organic components of toads. Biol. Sci. 2014, 103, 57. [Google Scholar]
- MAB National Committee of DPR Korea. Red Data Book of DPRK; National Committee of DPR Korea, Ed.; Science and Technology Publisher: Pyongyang, Democratic People’s Republic of Korea, 2016; Volume Animal. [Google Scholar]
- Chong, J.-R.; Hong, Y.J.; Hong, Y.G.; Ie, D.S. A Directory of Wetlands of D. P. R. Korea; Wild Bird Society of Japan and Korea University: Pyongyang, Democratic People’s Republic of Korea, 2016. [Google Scholar]
- Kim, L.; Han, G. Chosun Animal Encyclopedia, Herpetology Volume; Roh, H., Ed.; Science and Technology Publisher: Pyongyang, Democratic People’s Republic of Korea, 2009; p. 138. [Google Scholar]
- Ri, J. Systematics of the frogs (Anura) of our country. Biol. Sci. 2018, 107, 64. [Google Scholar]
- Song, J.-Y. Current status and distribution of Amphibian in North Korea. J. Natl. Park Res. 2016, 7, 211–217. [Google Scholar]
- Werner, F. Über Reptilien und Batrachier aus Guatemala und China in der zoologischen Staats-Sammlung in München nebst einem Abhang über seltene Formen aus Anderer gebieten. Abh. Der Math. Phys. Cl. Königlich Bayer. Akad. Wiss. 1903, 22, 342–384. [Google Scholar]
- Kaneko, Y.; Matsui, M. The IUCN Red List of Threatened Species; IUCN, Ed.; IUCN: Gland, Switzerland, 2004; Volume e.T58688A11811732. [Google Scholar]
- Kuzmin, S.; Ishchenko, V.; Tuniyev, B.; Beebee, T.; Andreone, F.; Nyström, P.; Anthony, B.P.; Schmidt, B.; Ogrodowczyk, A.; Ogielska, M.; et al. The IUCN Red List of Threatened Species; IUCN, Ed.; IUCN: Gland, Switzerland, 2009; Volume e.T58734A86470817. [Google Scholar]
- Xie, F.; Ye, C.; Fei, L.; Jiang, J.; Zeng, X.; Matsui, M. Taxonomical studies on Brown frogs (Rana) from Noertheastern China (amphibian: Ranidae). Acta Zootaxonomica Sin. 1999, 24, 224–231. [Google Scholar]
- Lu, Y.-Y.; Li, P.-P. A brief review on advance of Wood frogs research in China and discussing the importance of studying the Wood frog biodiversity around Bohai. Sichuan J. Zool. 2004, 24, 271–275. [Google Scholar]
- Yang, J.-X.; Zhou, W.-W.; Rao, D.-Q.; Poyarkov, A.N.; Kuzmin, S.L.; Che, J. Validity and systematic position of Rana altaica (Rana: Ranidae): Results of a phylogenetic analysis. Zool. Res. 2010, 31, 353–360. [Google Scholar]
- Schmidt, K.P. A new toad from Korea. Copeia 1931, 1931, 93–94. [Google Scholar] [CrossRef]
- Lim, C.H. Abour a new speice Bufo sambangi sp. n. Sci. Bull. 1996, 258, 39–41. [Google Scholar]
- Duellman, W.E.; Marion, A.B.; Hedges, S.B. Phylogenetics, classification, and biogeography of the treefrogs (Amphibia: Anura: Arboranae). Zootaxa 2016, 4104, 1–109. [Google Scholar] [CrossRef] [Green Version]
- Kuramoto, M. Mating calls of treefrogs (genus Hyla) in the far east, with description of a new species from Korea. Copeia 1980, 1, 100–108. [Google Scholar] [CrossRef]
- Chun, S.; Chung, E.; Voloshina, I.; Chong, J.R.; Lee, H.; Min, M.-S. Genetic diversity of korean tree frogs (Hyla suweonensis and Hyla japonica): Assessed by mitochondrial cytochrome b gene and cytochrome oxidase subunit I gene. Korean J. Herpetol. 2012, 4, 31–41. [Google Scholar]
- Borzée, A.; Seliger, B. Dryophytes suweonensis (Suweon Treefrog). Herpetol. Rev. 2018, 49, 707. [Google Scholar]
- Poyarkov, J.N.; Che, J.; Min, M.-S.; Kuro-o, M.; Yan, F.; Li, C.; Lizuka, K.; Vieites, D. Review of the systematics, morphology and distribution of Asian Clawed Salamanders, genus Onychodactylus (Amphibia, Caudata: Hynobiidae), with the description of four new species. Zootoxa 2012, 3465, 1–106. [Google Scholar] [CrossRef]
- Maslova, I.V.; Portnyagina, E.Y.; Sokolova, D.A.; Vorobieva, P.A.; Akulenko, M.V.; Portnyagin, A.S.; Somov, A.A. Distribution of rare and endangered amphibians and reptiles in Primorsky Krai (far east, Russia). Nat. Conserv. Res. 2018, 3, 61–72. [Google Scholar] [CrossRef]
- Borzée, A.; Andersen, D.; Groffen, J.; Kim, H.-T.; Bae, Y.; Jang, Y. Climate change-based models predict range shifts in the distribution of the only Asian plethodontid salamander: Karsenia koreana. Sci. Rep. 2019, 9, 11838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, P.J.; Angulo, A.; Lewis, J.P.; Moore, R.D.; Rabb, G.B.; Garcia Moreno, J. The amphibian extinction crisis-what will it take to put the action into the Amphibian Conservation Action Plan? Surv. Perspect. Integr. Environ. Soc. 2012, 5, 97–111. [Google Scholar]
- Lannoo, M.J. A perspective on amphibian conservation in the United States. Alytes 2012, 29, 133–144. [Google Scholar]
- Okada, Y. The tailless bactracians of the Japanese Empire. J. Imp. Agric. Exp. Stn. Nishigahara Tokyo 1931, 29, 1–215. [Google Scholar]
- Borzée, A.; Kyong, C.N.; Kil, H.K.; Jang, Y. Impact of water quality on the occurrence of two endangered Korean anurans: Dryophytes suweonensis and Pelophylax chosenicus. Herpetologica 2018, 74, 1–7. [Google Scholar] [CrossRef]
- Yeum, J.-H.; Lee, J.-H.; Lee, H.-J. Designation Properties of Ramsar Site for 20 Years in Korea. J. Environ. Sci. Int. 2018, 27, 319–332. [Google Scholar] [CrossRef]
- Son, E.; Borzée, A. Pelophylax chosenicus (Gold-spotted Pond Frog), Geographic distribution note. Herpetol. Rev. 2020, 51, 134. [Google Scholar]
- Borzée, A.; Bae, Y.H.; Seliger, B. Pelophylax chosenicus (Gold-spotted Pond Frog). Herpetol. Rev. 2018, 49, 709. [Google Scholar]
- Gunther, A. Catalogue of the Batrachia Salientia in the Collection of the British Museum; Collection of the British Museum; British Museum: London, UK, 1958. [Google Scholar]
- Kuzmin, S.; Maslova, I.; Matsui, M.; Liang, F.; Kaneko, Y. Dryophytes Japonicus (Amended Version of 2014 Assessment); A. S. Group, Ed.; The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2017. [Google Scholar]
- Roh, G.; Borzée, A.; Jang, Y. Spatiotemporal distributions and habitat characteristics of the endangered treefrog, Hyla suweonensis, in relation to sympatric H. japonica. Ecol. Inform. 2014, 24, 78–84. [Google Scholar] [CrossRef]
- Dufresnes, C.; Litvinchuk, S.N.; Borzée, A.; Jang, Y.; Li, J.-T.; Miura, I.; Perrin, N.; Stöck, M. Phylogeography reveals an ancient cryptic radiation in East-Asian tree frogs (Hyla japonica group) and complex relationships between continental and island lineages. BMC Evol. Biol. 2016, 16, 253. [Google Scholar]
- Fei, L.; Ye, C.-Y.; Huang, Y.-Z.; Liu, M. Atlas of amphibians of China; Henan Science and Technology Press: Zhengzhou, China, 1999. [Google Scholar]
- Liu, P.; Zhang, Z.; Zhao, S.; Zhao, W. Effect of female reproduction and mate choice on sexual size dimorphism in the northeast treefrog Hyla ussuriensis (Anura: Hylidae) in China. Asian Herpetol. Res. 2012, 3, 273–279. [Google Scholar]
- Matsui, M. Description of a New Brown Frog from Tsushima Island, Japan (Anura: Ranidae: Rana). Zool. Sci. 2014, 31, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Günther, A.C.L.G. Description of a new frog from north-eastern Asia. Ann. Mag. Nat. Hist. 1876, 4, 387. [Google Scholar] [CrossRef]
- Yang, B.-T.; Zhou, Y.; Min, M.-S.; Matsui, M.; Dong, B.-J.; Li, P.-P.; Fong, J.J. Diversity and phylogeography of Northeast Asian brown frogs allied to Rana dybowskii (Anura, Ranidae). Mol. Phylogenet. Evol. 2017, 112, 148–157. [Google Scholar] [CrossRef]
- Boulenger, G.A. Notes sur les grenouilles rousses d’Asie. Bull. Société Zool. Fr. 1886, 11, 595–600. [Google Scholar]
- Song, J.-Y.; Matsui, M.; Chung, K.-H.; Oh, H.-S.; Zhao, W. Distinct specific status of the Korean brown frog, Rana amurensis coreana (Amphibia: Ranidae). Zool. Sci. 2006, 23, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-B.; Min, M.-S.; Yang, S.-Y.; Matsui, M. Genetic relationships among Korean brown frog species (Anura, Ranidae), with special reference to evolutionary divergences between two allied species Rana dybowskii and R. huanrenensis. Zool. Sci. 2002, 19, 369–382. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.-Y.; Zhang, S.-Q.; Liu, M. A new species of Ranidae fron Liaoning, China (Anura). Acta Zootaxonomica Sin. 1993, 18, 493–497. [Google Scholar]
- Yang, S.Y.; Kim, J.B.; Min, M.S.; Suh, J.H.; Kang, Y.J.; Matsui, M.; Fei, L. First record of a brown frog Rana huanrenensis (Family Ranidae) from Korea. Korean J. Biol. Sci. 2000, 4, 45–50. [Google Scholar] [CrossRef]
- Che, J.; Pang, J.; Zhao, E.-M.; Matsui, M.; Zhang, Y.-P. Phylogenetic relationships of the Chinese brown frogs (genus Rana) inferred from partial mitochondrial 12S and 16S rRNA gene sequences. Zool. Sci. 2007, 24, 71–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Yang, B.-T.; Li, P.-P.; Min, M.-S.; Fong, J.J.; Dong, B.-J.; Zhou, Z.-Y.; Lu, Y.-Y. Molecular and morphological evidence for Rana kunyuensis as a junior synonym of Rana coreana (Anura: Ranidae). J. Herpetol. 2015, 49, 302–307. [Google Scholar] [CrossRef]
- Shannon, F.A. The Reptiles and Amphibians of Korea. Herpetologica 1956, 12, 22–49. [Google Scholar]
- Szyndlar, Z. A description of a small collection of amphibians and reptiles from the Democratic People’s Republic of Korea with notes on the distribution of the herpetofauna in that country. Acta Zool. Crac. 1984, 17, 1–11. [Google Scholar]
- Borzée, A.; Groffen, J.; Seliger, B. Bombina orientalis (Oriental Fire-bellied Toad). Herpetol. Rev. 2018, 49, 69. [Google Scholar]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wake, D.B. Facing extinction in real time. Science 2012, 335, 1052–1053. [Google Scholar] [CrossRef] [PubMed]
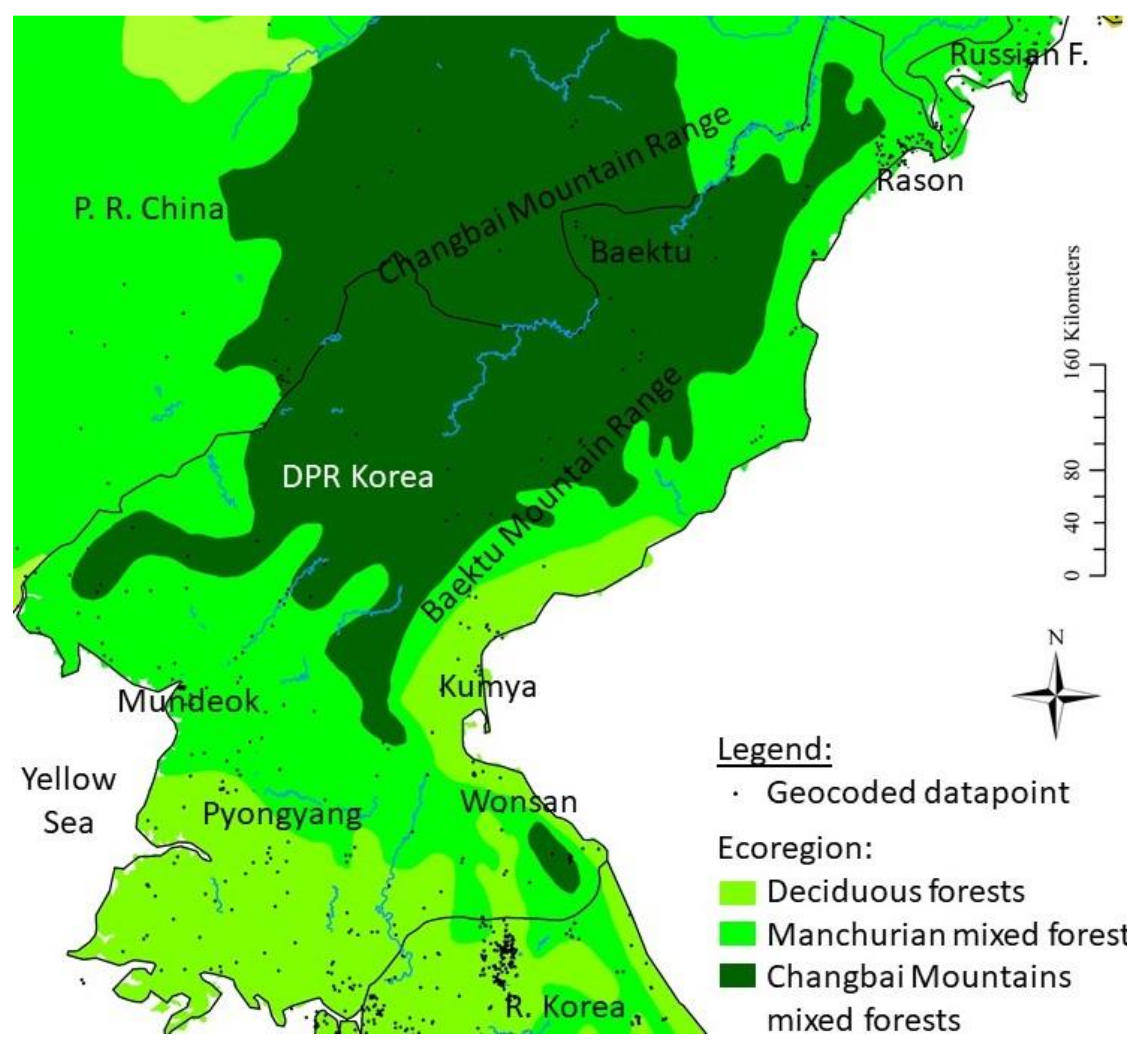
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’Amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. BioScience 2013, 51, 933–938. [Google Scholar] [CrossRef]
- de Magalhães, R.F.; Rocha, P.C.; Santos, F.R.; Strüssmann, C.; Giaretta, A.A. Integrative taxonomy helps to assess the extinction risk of anuran species. J. Nat. Conserv. 2018, 45, 1–10. [Google Scholar] [CrossRef]
- Rohland, N.; Hofreiter, M. Ancient DNA extraction from bones and teeth. Nat. Protoc. 2007, 2, 1756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, H.Q.; Kim, Y.I.; Borzée, A.; Jang, Y. Efficient isolation method for high-quality genomic DNA from cicada exuviae. Ecol. Evol. 2017, 7, 8161–8169. [Google Scholar] [CrossRef] [Green Version]
- Campos, P.F.; Gilbert, T.M. Ancient DNA; Shapiro, B., Hofreiter, M., Eds.; Humana Press: New York, NY, USA, 2012; Volume 840, pp. 81–85. [Google Scholar]
- Gilbert, M.T.P.; Haselkorn, T.; Bunce, M.; Sanchez, J.J.; Lucas, S.B.; Jewell, L.D.; Van Marck, E.; Worobey, M. The isolation of nucleic acids from fixed, paraffin-embedded tissues—Which methods are useful when? PLoS ONE 2007, 2, e537. [Google Scholar] [CrossRef]
- Qiagen. Sample & Assay Technologies. In QIAamp DNA FFPE Tissue Handbook; Qiagen: Venlo, The Netherlands, 2012. [Google Scholar]
- Hua, X.; Fu, C.; Li, J.; Oca, A.N.M.D.; Wiens, J.J. A revised phylogeny of holarctic treefrogs (genus Hyla) based on nuclear and mitochondrial DNA sequences. Herpetologica 2009, 65, 246–259. [Google Scholar] [CrossRef]
- Jeong, T.J.; Jun, J.; Han, S.; Kim, H.T.; Oh, K.; Kwak, M. DNA barcode reference data for the Korean herpetofauna and their applications. Mol. Ecol. Resour. 2013, 13, 1019–1032. [Google Scholar] [CrossRef]
- Igawa, T.; Kurabayashi, A.; Usuki, C.; Fujii, T.; Sumida, M. Complete mitochondrial genomes of three neobatrachian anurans: A case study of divergence time estimation using different data and calibration settings. Gene 2008, 407, 116–129. [Google Scholar] [CrossRef]
- Li, J.-T.; Wang, J.-S.; Nian, H.-H.; Litvinchuk, S.N.; Wang, J.; Li, Y.; Rao, D.-Q.; Klaus, S. Amphibians crossing the bering land bridge: Evidence from holarctic treefrogs (Hyla, Hylidae, Anura). Mol. Phylogenet. Evol. 2015, 87, 80–90. [Google Scholar] [CrossRef]
- Borzée, A.; Didinger, C.; Jang, Y. The Complete Mitochondrial Genome of Dryophytes suweonensis (Anura Hylidae). Mitochondrial DNA Part B Resour. 2017, 1, 5–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.-Y.; Jeon, H.S.; Min, M.-S.; An, J. Sequencing and analysis of the complete mitochondrial genome of Hyla suweonensis (Anura: Hylidae). Mitochondrial DNA Part B 2017, 2, 126–127. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Xie, Y.; Zhao, W.; Liu, P. Sequencing and analysis of the complete mitochondrial genome of Hyla ussuriensis (Anura: Hylidae). Mitochondrial DNA Part A 2017, 28, 328–329. [Google Scholar] [CrossRef] [PubMed]
- Sumida, M.; Ueda, H.; Nishioka, M. Reproductive isolating mechanisms and molecular phylogenetic relationships among Palearctic and Oriental brown frogs. Zool. Sci. 2003, 20, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, S.; Zhu, H.; Li, P.; Yang, B.; Ma, J. Phylogeny and biogeography of South Chinese brown frogs (Ranidae, Anura). PLoS ONE 2017, 12, e0175113. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Okada, Y. Frogs in Korea. J. Chosen Biol. Soc. 1928, 6, 15–46. [Google Scholar]
- Milto, K.D.; Barabanov, A.V. An annotated catalogue of the amphibian types in the collection of the Zoological Institute, Russian Academy of Sciences, St. Petersburg. Russ. J. Herpetol. 2011, 18, 137–153. [Google Scholar]
- Berman, D.B.N.; Balan, I. The Siberian frog. Priroda 2017, 8. n.a.. [Google Scholar]
- Stejneger, L. Herpetology of Japan and Adjacent Territory; Smithsonian Institute: Washington, DC, USA, 1907. [Google Scholar]
- Borzée, A.; Park, S.; Kim, A.; Kim, H.-T.; Jang, Y. Morphometrics of two sympatric species of tree frogs in Korea: A morphological key for the critically endangered Hyla suweonensis in relation to H. japonica. Anim. Cells Syst. 2013, 17, 348–356. [Google Scholar] [CrossRef]
- Borzée, A.; Messenger, K.R.; Chae, S.; Andersen, D.; Groffen, J.; Kim, Y.I.; An, J.; Othman, S.; Ri, K.; Nam, T.Y.; et al. Yellow sea mediated segregation between North East Asian Dryophytes species. PLoS ONE 2020, 15, e0234299. [Google Scholar]
- Borzée, A.; Kong, S.; Didinger, C.; Nguyen, H.; Jang, Y. A ring-species or a ring of species? Phylogenetic relationship between two treefrog species, Dryophytes suweonensis and D. immaculatus, around the Yellow Sea. Herpetol. J. 2018, 28, 160–170. [Google Scholar]
- Borzée, A.; Kim, K.; Heo, K.; Jablonski, P.G.; Jang, Y. Impact of land reclamation and agricultural water regime on the distribution and conservation status of the endangered Dryophytes suweonensis. PeerJ 2017, 5, e3872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borzée, A.; Andersen, D.; Jang, Y. Population trend inferred from aural surveys for calling anurans in Korea. PeerJ 2018, 6, e5568. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.W.; Nydegger, N.C. A spotlight, line-transect method for surveying jack rabbits. J. Wildl. Manag. 1985, 49, 699–702. [Google Scholar] [CrossRef]
- R Core Team. R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Araya-Salas, M.; Smith-Vidaurre, G. WarbleR: An R package to streamline analysis of animal acoustic signals. Methods Ecol. Evol. 2017, 8, 184–191. [Google Scholar] [CrossRef]
- Sueur, J.; Aubin, T.; Simonis, C.; Lellouch, L.; Brown, E.C.; Depraetere, M.; Desjonqueres, C.; Fabianek, F.; Gasc, A.; Kasten, E.; et al. Package “Seewave”. 2018. Available online: https://cran.r-project.org/web/packages/seewave/index.html (accessed on 1 December 2020).
- Ligges, U.; Krey, S.; Mersmann, O.; Schnackenberg, S.; Guenard, G.; Preusser, A.; Thieler, A.; Mielke, J.; Weihs, C. Package “TtuneR”. 2008. Available online: https://cran.r-project.org/web/packages/tuneR/index.html (accessed on 1 December 2020).
- Park, S.; Jeong, G.; Jang, Y. No reproductive character displacement in male advertisement signals of Hyla japonica in relation to the sympatric H. suweonensis. Behav. Ecol. Sociobiol. 2013, 67, 1345–1355. [Google Scholar] [CrossRef]
- Won, H.K.; Choy, Y.G. Checklist of Korean Chordata: Amphibia, Reptilia; Publication of the Korean Academy of Science: Pyongyang, Democratic People’s Republic of Korea, 1967; pp. 167–245. [Google Scholar]
- Won, H.K. Amphibian and Reptilian Fauna of Korea; Publication of the Korean Academy of Science: Pyongyang, Democratic People’s Republic of Korea, 1971; p. 170. [Google Scholar]
- Nikolsky, A.M. Faune de la Russie et des Pays limitrophes; Izd. Ros. Akad. Nauk: Petrograd, Russia, 1918; p. 309. [Google Scholar]
- Slevin, J. Contributions to oriental herpetology II. Korea or Chosen. Calif. Acad. Sci. 1925, 115, 89–100. [Google Scholar]
- Okada, Y. A study on the distribution of tailess batrachians of Japan. Annot. Zool. Jpn. 1926, 11, 137–144. [Google Scholar]
- Slevin, J.R. Contributions to Oriental herpetology. V. Honshu Hondo Neighboring Isl. Sado Awaji Seven Isl. Idzu. Ibid 1937, 23, 175–190. [Google Scholar]
- Kang, Y.S.; Yoon, I.B. Amphibia and Reptilia; Illustrated Encyclopedia of Fauna and Flora of Korea; Ministry of Education: Seoul, Republic of Korea, 1975; Volume 17, p. 191. [Google Scholar]
- Lee, S.; Lee, K. The Yearbook of Natural Monument in D. P. R. Korea; Agriculture Press: Pyeongyang, Democratic People’s Republic of Korea, 1994. [Google Scholar]
- Pawlowski, J.; Tomek, T. Zoological expeditions to the North Korea organized in the years 1971–1992 by the Cracow Institute of Systematics and Evolutions of Animals of the Polish Academy of Science. Fragm. Faun. 1997, 40, 231–246. [Google Scholar] [CrossRef] [Green Version]
- Cantor, T. General features of Chusan, with remarks on the flora and fauna of that island. Ann. Mag. Nat. Hist. 1842, 1, 481–493. [Google Scholar] [CrossRef] [Green Version]
- Mészáros, F.; Zombori, L. Zoological collectings by the Hungarian Natural History Museum in Korea. 107. A report of the collecting of the fourteenth expedition. Folia Entomol. Hung. 1992, 52, 73–80. [Google Scholar]
- Lee, S.; Gil, J. A study of mammal and amphibians-reptile species around Shin-Po area, Hamkyungbuk-do, North Korea. Korean Wetl. Soc. 2007, 9, 13–20. [Google Scholar]
- Schmidt, P.Y. News from the Korean-Sakhalin expedition [Vesti is Koreysko-Sakhalinskoy ekspeditsii]. Izv. Imp. Rus. Geogr. Obs. St. Petersburg. 1900, 36, 463–501. [Google Scholar]
- Borkin, L. On rare Far Eastern toad species Bufo stejnegeri Schmidt, 1931. Proc. Zool. Inst. Leningr. 1986, 157, 192–196. [Google Scholar]
- Strauch, A. Mongolia and the Tangut Country: A Three Years’ Journey in the Eastern Highlands of Asia; Izd. Imp. Russkago Ob-va: Sankpeterburg, Russia, 1876; Volume 2. [Google Scholar]
- Boulenger, G. Annals and Magazine of Natural History (5), XIX; not of Audouin; Taill. Batr. Europe, Pt.: Gensan, Korea, 1887; p. 67. [Google Scholar]
- Borzée, A. First amphibian behavioural observation from the Democratic People’s Republic of Korea: Predation of a Dryophytes japonicus tadpole by Hydaticus sp. larvae. Nat. Conserv. Res. 2019, 4, 106–108. [Google Scholar] [CrossRef]
- Boulenger, G.A. A list of the reptiles and batrachians of Amoorland. Ann. Mag. Nat. Hist. 1890, 6, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Zvegintsov, A. Travel in North Korea [Poezdka v severnuyu Koreyu]. Izv. Imp. Rus. Geogr. Obs. St. Petersburg 1900, 36, 502–518. [Google Scholar]
- Suk, J. Collection record of animal in Mt. Baekdusan area. J. Chosen Biol. Soc. 1934, 18, 129–136. [Google Scholar]
- California Academy of Sciences. Herpetology 32497; California Academy of Sciences, Ed.; California Academy of Sciences: San Francisco, CA, USA, 2017; Volume 2019. [Google Scholar]
- California Academy of Sciences. Herpetology 32483; California Academy of Sciences, Ed.; California Academy of Sciences: San Francisco, CA, USA, 2017; Volume 2019. [Google Scholar]
- California Academy of Sciences. Herpetology 32519; California Academy of Sciences, Ed.; California Academy of Sciences: San Francisco, CA, USA, 2017; Volume 2019. [Google Scholar]
- Barbour, T. Some new reptiles and amphibians. Bull. Mus. Comp. Zool. 1908, 51, 315–325. [Google Scholar]
- Kim, R.; Ju, J.; Sin, G.; Choi, G.; Park, H.; Go, Y.; Seok, S. The Illustrated Book of the Korean Natural Monuments 3: Animals Part; Seong, J., Kim, S., Eds.; Korean Culture Preservation Company: Pyongyang, Democratic People’s Republic of Korea, 2007; Volume 96. [Google Scholar]
- Choi, D.; Park, C.; Jang, U.; Jeong, S.; Go, I.; Park, Y.; Gang, C. Gwangmyeong Encyclopedia: Agriculture, Forestry, Fishery; Gwangmyeong Encyclopedia; Pyongyang Printing Press: Pyongyang, Democratic People’s Republic of Korea, 2009; Volume 18, p. 888. [Google Scholar]
- California Academy of Sciences. Herpetology 36028; California Academy of Sciences, Ed.; California Academy of Sciences: San Francisco, CA, USA, 2017; Volume 2019. [Google Scholar]
- California Academy of Sciences. Herpetology 32735; California Academy of Sciences, Ed.; California Academy of Sciences: San Francisco, CA, USA, 2017; Volume 2019. [Google Scholar]
- Bannikov, A.; Darevsky, I.; Ishchenko, V.; Rustamov, A.; Szczerbak, N. Guide to Amphibians and Reptiles of the USSR Fauna; Prosveshchenie: Moscow, Russia, 1977; p. 414. [Google Scholar]
- California Academy of Sciences. Herpetology 32757; California Academy of Sciences, Ed.; California Academy of Sciences: San Francisco, CA, USA, 2019; Volume 2019. [Google Scholar]
- Nikolskii, A.M. Rana emeljanovi sp. n. Annu. Musée Zool. L’academie Impériale Sci. St. Pétersbourg 1913, 18, 148–150. [Google Scholar]
- Hallowell, E. Report upon the Reptilia of the North Pacific Exploring Expedition, under command of Capt. John Rogers. Proc. Acad. Nat. Sci. Phila. 1861, 12, 480–510. [Google Scholar]
- California Academy of Sciences. Herpetology 32875; California Academy of Sciences, Ed.; California Academy of Sciences: San Francisco, CA, USA, 2017; Volume 2019. [Google Scholar]
- Boulenger, G. Description of a new tailed batrachian from Corea. J. Nat. Hist. 1887, 14, 67. [Google Scholar] [CrossRef]
- Kim, J. Natural monuments of our country. Chosun Nat. Conserv. Prot. Group Cent. Branch 2017, 4, 50–51. [Google Scholar]
- Kim, K.; Kim, M.; Kim, R.; Kim, K.; Kim, T.; Rim, C.; Pak, U.; Han, K. Red Data Book of Democratic People’s Republic of Korea; MAB national committee of Democratic People’s Republic of Korea: Pyongyang, Democratic People’s Republic of Korea, 2002. [Google Scholar]
- Mori, T. On a new Hynobius from Quelpaert Isl. J. Chosen Nat. Hist. Soc. 1928, 6, 47–52. [Google Scholar]
- Nadachowski, A.; Szyndlar, Z.; Tomek, T. Fauna kregowcow ladowych polwyspu Koreanskiego. Prz. Zool. 1982, 26, 79–92. [Google Scholar]
- Shin, Y.; Jang, Y.; Allain, S.J.R.; Borzée, A. Catalogue of herpetological specimens of the Ewha Womans University Natural History Museum (EWNHM), Republic of Korea. ZooKeys 2020, 965, 103–139. [Google Scholar] [CrossRef]
- Lu, Y.Y.; Wang, J.Q.; Dong, B.J.; Li, P.P. A New Record of Rana huanrenensis in Jilin Province, China. Sichuan J. Zool. 2005, 3, 3. [Google Scholar]
- Jiang, X. Thoughts on the development of ecotourism in Laoguiding Nature Reserve of Huanren. For. Prod. Spec. China 2015, 6, 89–91. [Google Scholar]
- Malyarchuk, B.; Derenko, M.; Berman, D.; Perkova, M.; Grzybowski, T.; Lejrikh, A.; Bulakhova, N. Phylogeography and molecular adaptation of Siberian salamander Salamandrella keyserlingii based on mitochondrial DNA variation. Mol. Phylogenet. Evol. 2010, 56, 562–571. [Google Scholar] [CrossRef]
- Min, M.S.; Yang, S.-Y.; Bonett, R.; Vieites, D.; Brandon, R.; Wake, D. Discovery of the first Asian plethodontid salamander. Nature 2005, 435, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Tytar, V.; Nekrasova, O.; Pupina, A.; Pupins, M.; Oskyrko, O. Long-term bioclimatic modelling the distribution of the fire-bellied toad, Bombina bombina (Anura, Bombinatoridae), under the influence of global climate change. Vestn. Zool. 2018, 52, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Maes, D.; Isaac, N.J.; Harrower, C.A.; Collen, B.; Van Strien, A.J.; Roy, D.B. The use of opportunistic data for IUCN Red List assessments. Biol. J. Linn. Soc. 2015, 115, 690–706. [Google Scholar] [CrossRef] [Green Version]
- Lamoreux, J.; Akçakaya, H.R.; Bennun, L.; Collar, N.J.; Boitani, L.; Brackett, D.; Bräutigam, A.; Brooks, T.M.; da Fonseca, G.A.; Mittermeier, R.A. Value of the IUCN red list. Trends Ecol. Evol. 2003, 18, 214–215. [Google Scholar]
- Rodrigues, A.S.; Pilgrim, J.D.; Lamoreux, J.F.; Hoffmann, M.; Brooks, T.M. The value of the IUCN Red List for conservation. Trends Ecol. Evol. 2006, 21, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Keller, V.; Bollmann, K. From red lists to species of conservation concern. Conserv. Biol. 2004, 18, 1636–1644. [Google Scholar] [CrossRef]
- Fitzpatrick, U.; Murray, T.E.; Paxton, R.J.; Brown, M.J. Building on IUCN regional red lists to produce lists of species of conservation priority: A model with Irish bees. Conserv. Biol. 2007, 21, 1324–1332. [Google Scholar] [CrossRef] [Green Version]
- Simaika, J.P.; Samways, M.J. Reserve selection using Red Listed taxa in three global biodiversity hotspots: Dragonflies in South Africa. Biol. Conserv. 2009, 142, 638–651. [Google Scholar] [CrossRef]
- Butchart, S.H.; Akcakaya, H.R.; Kennedy, E.; Hilton-Taylor, C. Biodiversity indicators based on trends in conservation status: Strengths of the IUCN Red List Index. Conserv. Biol. 2006, 20, 579–581. [Google Scholar] [CrossRef]
- Mace, G.M.; Collar, N.J.; Gaston, K.J.; Hilton-Taylor, C.; Akçakaya, H.R.; Leader-Williams, N.; Milner-Gulland, E.J.; Stuart, S.N. Quantification of extinction risk: IUCN’s system for classifying threatened species. Conserv. Biol. 2008, 22, 1424–1442. [Google Scholar] [CrossRef]
- Gärdenfors, U.; Hilton-Taylor, C.; Mace, G.M.; Rodríguez, J.P. The application of IUCN Red List criteria at regional levels. Conserv. Biol. 2001, 15, 1206–1212. [Google Scholar] [CrossRef]
- Miller, R.M.; Rodríguez, J.P.; Fowler, T.A.; Bambaradeniya, C.; Boles, R.; Eaton, M.A.; Gärdenfors, U.; Keller, V.; Molur, S.; Walker, S. National threatened species listing based on IUCN criteria and regional guidelines: Current status and future perspectives. Conserv. Biol. 2007, 21, 684–696. [Google Scholar] [CrossRef]
- IUCN. Guidelines for Application of IUCN Red List Criteria at Regional and National Levels: Version 4.0; IUCN Species Survival Commission (SSC): Gland, Switzerland, 2012. [Google Scholar]
- Mace, G.M. Classifying threatened species: Means and ends. Phil. Trans. R. Soc. Lond. B 1994, 344, 91–97. [Google Scholar]
- Hermoso, V.; Kennard, M.J.; Linke, S. Evaluating the costs and benefits of systematic data acquisition for conservation assessments. Ecography 2015, 38, 283–292. [Google Scholar] [CrossRef]
- IUCN. International Union for Conservation of Nature Red List; IUCN, Ed.; IUCN: Gland, Switzerland, 2018. [Google Scholar]
- Kim, Y.I. Intraspecific Latitudinal Variation in Critical Thermal Maximum of Tadpoles in Two Anuran Species; Ewha Womans University: Seoul, Korea, 2018. [Google Scholar]
- Heo, K.; Kim, Y.I.; Bae, Y.; Jang, Y.; Amaël, B. First report of Dryophytes japonicus tadpoles in saline environment. Russ. J. Herpetol. 2019, 26, 87–90. [Google Scholar] [CrossRef]
- Borzée, A.; Purevdorj, Z.; Kim, Y.I.; Kong, S.; Choe, M.; Yi, Y.; Kim, K.; Kim, A.; Jang, Y. Breeding preferences in the Treefrogs Dryophytes japonicus (Hylidae) in Mongolia. J. Nat. Hist. 2019, 53, 43–44. [Google Scholar]
- Gosner, K.L. A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica 1960, 16, 183–190. [Google Scholar]
- Matsui, M. Pelophylax chosenicus. In The IUCN Red List Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T58577A11806007. [Google Scholar] [CrossRef]
- Lee, S.-D.; Miller-Rushing, A.J. Degradation, urbanization, and restoration: A review of the challenges and future of conservation on the Korean Peninsula. Biol Cons 2014, 176, 262–276. [Google Scholar] [CrossRef]
- Borzée, A.; Struecker, M.-Y.; Yi, Y.; Kim, D.; Kim, H. Time for Korean wildlife conservation. Science 2019, 363, 1161–1162. [Google Scholar]
- The World Bank. World Development Indicators; The World Bank: Washington, DC, USA, 2013. [Google Scholar]
- Stone, R. Seeking cures for North Korea’s environmental ills. Science 2012, 335, 1425–1426. [Google Scholar] [CrossRef] [Green Version]
- UNEP. United Nations Environment Programme; Piddington, K., Ed.; UNEP: Klong Luang, Thailand, 2003. [Google Scholar]
- Democratic People’s Republic of Korea. National Biodiversity Strategy and Action Plan of DPR Korea; Democratic People’s Republic of Korea: Pyongyang, Democratic People’s Republic of Korea, 2007. [Google Scholar]
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 987–997. [Google Scholar] [CrossRef]
- IUCN SSC Amphibian Specialist Group. Dryophytes Suweonensis. In The IUCN Red List of Threatened Species; IUCN SSC Amphibian Specialist Group; IUCN: Gland, Switzerland, 2017. [Google Scholar] [CrossRef]
- Kuzmin, S.; Pipeng, L.; Matsui, M.; Ishchenko, V.; Maslova, I. Bombina orientalis. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T54449A11146991. [Google Scholar] [CrossRef]
- Kuzmin, S.; Yuezhao, W.; Matsui, M.; Kaneko, Y.; Maslova, I. Bufo gargarizans. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T54647A11180910. [Google Scholar]
- Matsui, M.; Wenge, Z. Bufo stejnegeri. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T54766A11200818. [Google Scholar] [CrossRef]
- Koo, K.-S. Distribution Characteristics and Body Size Variation of Japanese Tree Frog (Hyla japonica) in South Korea; Gangwon National University: Chuncheon, Korea, 2018. [Google Scholar]
- Borzée, A. Why Are Anurans Threatened? The Case of Dryophytes suweonensis. Ph.D Thesis, Seoul National University, Seoul, Korea, 2018. [Google Scholar]
- Borzée, A.; Fong, J.J.; Nguyen, H.; Jang, Y. Large-scale hybridisation as an extinction threat to the Suweon treefrog (Hylidae: Dryophytes suweonensis). Animals 2020, 10, 764. [Google Scholar] [CrossRef] [PubMed]
- Groffen, J.; Kong, S.; Jang, Y.; Borzée, A. The invasive American bullfrog (Lithobates catesbeianus) in the Republic of Korea: History and recommendation for population control. Manag. Biol. Invasions 2019, 10, 517–535. [Google Scholar] [CrossRef] [Green Version]
- Borzée, A.; Kosch, T.A.; Kim, M.; Jang, Y. Introduced bullfrogs are associated with increased Batrachochytrium dendrobatidis prevalence and reduced occurrence of Korean treefrogs. PLoS ONE 2017, 12, e0177860. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Dryophytes suweonensis (amended version of 2014 assessment). In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2017; Volume e.T55670A112715252. [Google Scholar] [CrossRef]
- Kuzmin, S.; Matsui, M.; Wenge, Z.; Kaneko, Y. Glandirana emeljanovi. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T58591A11793116. [Google Scholar] [CrossRef]
- Matsui, M.; Wenge, Z. Hynobius leechii. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2008; Volume e.T59095A11870831. [Google Scholar]
- Matsui, M.; Wenge, Z. The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T57849A11693398. [Google Scholar]
- Kuzmin, S.; Matsui, M.; Wenge, Z.; Maslova, I.; Tuniyev, B. Onychodactylus fischeri. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T39419A10234504. [Google Scholar] [CrossRef]
- Kuzmin, S.; Maslova, I.; Tuniyev, B.; Matsui, M.; Pipeng, L.; Kaneko, Y. Pelophylax nigromaculatus. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T58679A11809026. [Google Scholar]
- Litvinchuk, S.N.; Schepina, N.A.; Borzée, A. Reconstruction of past distribution for the Mongolian toad, Strauchbufo raddei (Anura: Bufonidae) using environmental modeling. PeerJ 2020, 8, e9216. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, S.; Matsui, M.; Gang, L.; Maslova, I. Strauchbufo raddei. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T54744A11198066. [Google Scholar]
- Kuzmin, S.; Maslova, I.; Matsui, M.; Wenge, Z. Rana amurensis. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2008; Volume e.T58542A11799162. [Google Scholar] [CrossRef]
- Song, J.-Y. Rana coreana. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2008; Volume e.T135860A4212618. [Google Scholar] [CrossRef]
- Kuzmin, S.; Ishchenko, V.; Maslova, I.; Ananjeva, N.; Orlov, N.; Matsui, M.; Feng, X.; Kaneko, Y. Rana dybowskii. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T58589A11792510. [Google Scholar] [CrossRef]
- Matsui, M.; Feng, X. Rana huanrensis. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; Volume e.T58619A11812585. [Google Scholar] [CrossRef]
- Fei, L.; Changyuan, Y.; Jianping, J. Colored Atlas of Chinese Amphibians and Their Distributions; Sichuan science and technology Press: Chendu, China, 2012. [Google Scholar]
- Korotkov, Y.M. To the Biology of Bombina Orientalis, Agkistrodon Blomhoffii and A. halis in Primorye Region; Zoologicheskie Problemy Sibiri: Novosibirsk, Russia, 1972; Volume 302. [Google Scholar]
- Korotkov, Y.M.; Korotkova, E.B. Some patterns of reproduction of Rana chensinensis in the Far East marine territory. Zool. Zhurnal 1977, 56, 1057–1061. [Google Scholar]
- Piao, R.Z. List of Wildlife in Heilongjiang China; The institute of wildlife in Heilongjiang province China: Harbin, China, 1990. [Google Scholar]
- Fok, K.W.; Wade, C.M.; Parkin, D.T. Inferring the phylogeny of disjunct populations of the azure–winged magpie Cyanopica cyanus from mitochondrial control region sequences. Proc. R. Soc. Lond. B Biol. Sci. 2002, 269, 1671–1679. [Google Scholar] [CrossRef] [Green Version]
- Zanini, F.; Klingemann, A.; Schlaepfer, R.; Schmidt, B.R. Landscape effects on anuran pond occupancy in an agricultural countryside: Barrier-based buffers predict distributions better than circular buffers. Can. J. Zool. 2008, 86, 692–699. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, G.M.; Hines, A.H.; Jivoff, P. Biotic resistance to invasion: Native predator limits abundance and distribution of an introduced crab. Ecology 2005, 86, 3364–3376. [Google Scholar]
- Marco, A.; Quilchano, C.; Blaustein, A.R. Sensitivity to nitrate and nitrite in pond-breeding amphibians from the Pacific Northwest, USA. Environ. Toxicol. Chem. 1999, 18, 2836–2839. [Google Scholar] [CrossRef]
- Macias, D.A.; Groffen, J.; Jang, Y.; Borzée, A. Rana coreana (Korean Brown Frog) and R. uenoi (Ueno’s Brown Frog). Hibernaculum. Herpetol. Rev. 2018, 49, 121–122. [Google Scholar]
- Borzée, A.; Choi, Y.; Kim, Y.E.; Jablonski, P.G.; Jang, Y. Interspecific variation in seasonal migration and brumation behavior in two closely related species of treefrogs. Front. Ecol. Evol. 2019, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.J. Speciation and ecology revisited: Phylogenetic niche conservatism and the origin of species. Evol. Int. J. Org. Evol. 2004, 58, 193–197. [Google Scholar] [CrossRef]
- Vences, M.; Wake, D.B. Amphibian Biology; Heatwole, H., Tyler-Michael, J., Eds.; Surrey Beatty and Sons: Norton, Australia, 2007; Volume 7, pp. 2613–2671. [Google Scholar]
- Hendry, A.P. Ecological speciation! Or the lack thereof? Can. J. Fish. Aquat. Sci. 2009, 66, 1383–1398. [Google Scholar] [CrossRef] [Green Version]
- Hua, X.; Wiens, J.J. How does climate influence speciation? Am. Nat. 2013, 182, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jo, Y.-S.; Baccus, J.T.; Koprowski, J. Mammals of Korea; National Institute of Biological Resources: Incheon, Korea, 2018. [Google Scholar]
- Brazil, M. Birds of East Asia: China, Taiwan, Korea, Japan, and Russia; Princeton University Press: Trenton, NJ, USA, 2009; p. 528. [Google Scholar]
- Hewitt, G. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef]
- Veith, M.; Kosuch, J.; Vences, M. Climatic oscillations triggered post-Messinian speciation of Western Palearctic brown frogs (Amphibia, Ranidae). Mol. Phylogenet. Evol. 2003, 26, 310–327. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography of Southern European Refugia; Weiss, S.F.N.E., Ed.; Phylogeography of southern European refugia; Springer: New York, NY, USA, 2007; p. 377. [Google Scholar]
- Knowles, L.L. Did the Pleistocene glaciations promote divergence? Tests of explicit refugial models in montane grasshopprers. Mol. Ecol. 2001, 10, 691–701. [Google Scholar] [CrossRef]
- Kent, M. Advanced Biology; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Nosil, P. Ecological Speciation; Oxford University Press: New York, NY, USA, 2012. [Google Scholar]
- Aizawa, M.; Kim, Z.-S.; Yoshimaru, H. Phylogeography of the Korean pine (Pinus koraiensis) in northeast Asia: Inferences from organelle gene sequences. J. Plant Res. 2012, 125, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Park, S.K.; Lee, H.; Oshida, T.; Kimura, J.; Kim, Y.J.; Nguyen, S.; Sashika, M.; Min, M.S. Phylogeography of Korean raccoon dogs: Implications of peripheral isolation of a forest mammal in East Asia. J. Zool. 2013, 290, 225–235. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to Quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Provan, J.; Bennett, K. Phylogeographic insights into cryptic glacial refugia. Trends Ecol. Evol. 2008, 23, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Stöck, M.; Dufresnes, C.; Litvinchuk, S.N.; Lymberakis, P.; Biollay, S.; Berroneau, M.; Borzée, A.; Ghali, K.; Ogielska, M.; Perrin, N. Cryptic diversity among Western Palearctic tree frogs: Postglacial range expansion, range limits, and secondary contacts of three European tree frog lineages (Hyla arborea group). Mol. Phylogenet. Evol. 2012, 65, 1–9. [Google Scholar]
- Zhang, H.; Yan, J.; Zhang, G.; Zhou, K. Phylogeography and demographic history of Chinese black-spotted frog populations (Pelophylax nigromaculata): Evidence for independent refugia expansion and secondary contact. BMC Evol. Biol. 2008, 8, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borzée, A.; Santos, J.L.; Sanchez-Ramirez, S.; Bae, Y.; Heo, K.; Jang, Y.; Jowers, M.J. Phylogeographic and population insights of the Asian common toad (Bufo gargarizans) in Korea and China: Population isolation and expansions as response to the ice ages. PeerJ 2017, 5, e4044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.E.; Yang, S.Y.; Lee, H.Y. Genetic relationships among six Korean Rana species (Amphibia; Ranidae) based on the mitochondrial cytochrome b gene. Korean J. Biol. Sci. 2000, 4, 117–121. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E. Large-scale processes and the Asian bias in species diversity of temperate plants. Nature 2000, 407, 180. [Google Scholar] [CrossRef] [PubMed]
- Serizawa, K.; Suzuki, H.; Iwasa, M.A.; Tsuchiya, K.; Pavlenko, M.V.; Kartavtseva, I.V.; Chelomina, G.N.; Dokuchaev, N.E.; Han, S.-H. A spatial aspect on mitochondrial DNA genealogy in Apodemus peninsulae from East Asia. Biochem. Genet. 2002, 40, 149–161. [Google Scholar] [CrossRef]
- Aizawa, M.; Yoshimaru, H.; Saito, H.; Katsuki, T.; Kawahara, T.; Kitamura, K.; Shi, F.; Kaji, M. Phylogeography of a northeast Asian spruce, Picea jezoensis, inferred from genetic variation observed in organelle DNA markers. Mol. Ecol. 2007, 16, 3393–3405. [Google Scholar] [CrossRef]
- Bai, W.N.; Liao, W.J.; Zhang, D.Y. Nuclear and chloroplast DNA phylogeography reveal two refuge areas with asymmetrical gene flow in a temperate walnut tree from East Asia. New Phytol. 2010, 188, 892–901. [Google Scholar] [CrossRef]
- Ding, L.; Gan, X.N.; He, S.P.; Zhao, E.M. A phylogeographic, demographic and historical analysis of the short-tailed pit viper (Gloydius brevicaudus): Evidence for early divergence and late expansion during the Pleistocene. Mol. Ecol. 2011, 20, 1905–1922. [Google Scholar] [CrossRef]
- Othman, S.N.; Putri, E.T.; Messenger, K.R.; Bae, Y.; Yang, Y.; Bova, T.; Reed, T.; Amin, H.; Chuang, M.-F.; Jang, Y.; et al. Impact of south-central Asian orogenesis on Kaloula spp. (Anura: Microhylidae) radiation and implication of Pleistocene glaciation on population extension to northern latitudes. Integr. Zool. 2021, (in press). [Google Scholar]
- Liu, K.; Wang, F.; Chen, W.; Tu, L.; Min, M.-S.; Bi, K.; Fu, J. Rampant historical mitochondrial genome introgression between two species of green pond frogs, Pelophylax nigromaculatus and P. plancyi. BMC Evol. Biol. 2010, 10, 201. [Google Scholar]
- Komaki, S.; Igawa, T.; Lin, S.M.; Tojo, K.; Min, M.S.; Sumida, M. Robust molecular phylogeny and palaeodistribution modelling resolve a complex evolutionary history: Glacial cycling drove recurrent mtDNA introgression among Pelophylax frogs in East Asia. J. Biogeogr. 2015, 42, 2159–2171. [Google Scholar] [CrossRef]
- Fairbanks, R.G. A 17,000-year glacio-eustatic sea level record: Influence of glacial melting rates on the Younger Dryas event and deep-ocean circulation. Nature 1989, 342, 637–642. [Google Scholar] [CrossRef]
- Park, Y.; Khim, B.; Zhao, S. Sea level fluctuation in the Yellow Sea Basin. J. Korean Soc. Oceanogr. 1994, 29, 42–49. [Google Scholar]
- Kim, J.-M.; Kennett, J.P. Paleoenvironmental changes associated with the Holocene marine transgression, Yellow Sea (Hwanghae). Mar. Micropaleontol. 1998, 34, 71–89. [Google Scholar] [CrossRef]
- Ijiri, A.; Wang, L.; Oba, T.; Kawahata, H.; Huang, C.-Y.; Huang, C.-Y. Paleoenvironmental changes in the northern area of the East China Sea during the past 42,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 239–261. [Google Scholar] [CrossRef]
- Nguyen, T.Q.; Bernardes, M.; Nguyen, T.T.; Ziegler, T. First records of Bufo gargarizans Cantor, 1842 and Odorrana lipuensis Mo, Chen, Wu, Zhang et Zhou, 2015 (Anura: Bufonidae, Ranidae) from Vietnam. Russ. J. Herpetol. 2016, 23, 103–107. [Google Scholar]
- Liu, W.; Lathrop, A.; Fu, J.; Yang, D.; Murphy, R.W. Phylogeny of East Asian bufonids inferred from mitochondrial DNA sequences (Anura: Amphibia). Mol. Phylogenet. Evol. 2000, 14, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Weadick, C.J.; Zeng, X.; Wang, Y.; Liu, Z.; Zheng, Y.; Li, C.; Hu, Y. Phylogeographic analysis of the Bufo gargarizans species complex: A revisit. Mol. Phylogenet. Evol. 2005, 37, 202–213. [Google Scholar] [CrossRef]
- Zhan, A.; Fu, J. Past and present: Phylogeography of the Bufo gargarizans species complex inferred from multi-loci allele sequence and frequency data. Mol Phylogenet. Evol 2011, 61, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Porta, J.; Litvinchuk, S.; Crochet, P.-A.; Romano, A.; Geniez, P.; Lo-Valvo, M.; Lymberakis, P.; Carranza, S. Molecular phylogenetics and historical biogeography of the west-palearctic common toads (Bufo bufo species complex). Mol. Phylogenet. Evol. 2012, 63, 113–130. [Google Scholar] [CrossRef]
- Tong, H.; Wo, Y.; Lioa, P.; Jin, Y. Phylogenetic, demographic and dating analyses of Bufo gargarizans populations from the Zhoushan archipelago and mainland China. Asian Herpetol. Res. 2017, 8, 165–173. [Google Scholar]
- Kong, W.S. Vegetational history of the Korean Peninsula. Glob. Ecol. Biogeogr. 2000, 9, 391–402. [Google Scholar] [CrossRef]
- Yan, F.; Zhou, W.; Zhao, H.; Yuan, Z.; Wang, Y.; Jiang, K.; Jin, J.; Murphy, R.W.; Che, J.; Zhang, Y. Geological events play a larger role than Pleistocene climatic fluctuations in driving the genetic structure of Quasipaa boulengeri (Anura: Dicroglossidae). Mol. Ecol. 2013, 22, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Ryu, E.; Lee, S.-J.; Yang, D.-Y.; Kim, J.-Y. Paleoenvironmental studies of the Korean peninsula inferred from diatom assemblages. Quat. Int. 2008, 176, 36–45. [Google Scholar] [CrossRef]
- He, L.; Mukai, T.; Chu, K.; Ma, Q.; Zhang, J. Biogeographical role of the Kuroshio Current in the amphibious mudskipper Periophthalmus modestus indicated by mitochondrial DNA data. Sci. Rep. 2015, 5, 15645. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Lin, H. Rainy season of the asian-pacific summer monsoon. J. Clim. 2002, 15, 386–398. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.J.; Jhun, J.G.; Park, C.K. Remote connection of the northeast Asian summer rainfall variation revealed by a newly defined monsoon index. J. Clim. 2005, 18, 4381–4393. [Google Scholar] [CrossRef]
- Beals, E.W. Vegetational change along altitudinal gradients. Science 1969, 165, 981–985. [Google Scholar] [CrossRef]
- Stevens, G.C. The elevational gradient in altitudinal range: An extension of Rapoport’s latitudinal rule to altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Blaschke, M.; Bässler, C. Altitudinal gradients in biodiversity research: The state of the art and future perspectives under climate change aspects. For. Ecol. Landsc. Res. Nat. Conserv. 2011, 11, 35–47. [Google Scholar]
- Willig, M.R.; Presley, S.J. Biodiversity and metacommunity structure of animals along altitudinal gradients in tropical montane forests. J. Trop. Ecol. 2016, 32, 421–436. [Google Scholar] [CrossRef]
- Loveridge, A. Scientific Results of an Expedition to Rain Forest Regions in Eastern Africa: Zoogeography and Itinerary. Bull. Mus. Comp. Zool. 1937, 79, 481–541. [Google Scholar]
- Daniels, R.R. Geographical distribution patterns of amphibians in the Western Ghats, India. J. Biogeogr. 1992, 19, 521–529. [Google Scholar] [CrossRef]
- Poynton, J.; Loader, S.; Sherratt, E.; Clarke, B. Amphibian diversity in East African biodiversity hotspots: Altitudinal and latitudinal patterns. Biodivers. Conserv. 2006, 16, 277–292. [Google Scholar]
- Cogălniceanu, D.; Székely, P.; Samoilă, C.; Ruben, I.; Tudor, M.; Plăiaşu, R.; Stănescu, F.; Rozylowicz, L. Diversity and distribution of amphibians in Romania. ZooKeys 2013, 296, 35. [Google Scholar]
- Gaston, K.J.; Fuller, R.A. The sizes of species’ geographic ranges. J. Appl. Ecol. 2009, 46, 1–9. [Google Scholar] [CrossRef]
- Dodd, C.K., Jr. Biological diversity of a temporary pond herpetofauna in north Florida sandhills. Biodivers. Conserv. 1992, 1, 125–142. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Biological Delineation of Terrestrial Buffer Zones for Pond-Breeding Salamanders. Conserv. Biol. 1998, 12, 1113–1119. [Google Scholar] [CrossRef]
- Natuhara, Y. Ecosystem services by paddy fields as substitutes of natural wetlands in Japan. Ecol. Eng. 2013, 56, 97–106. [Google Scholar] [CrossRef]
- Holzer, K.A. Amphibian use of constructed and remnant wetlands in an urban landscape. Urban Ecosyst. 2014, 17, 955–968. [Google Scholar] [CrossRef]
- Quesnelle, P.E.; Lindsay, K.E.; Fahrig, L. Relative effects of landscape-scale wetland amount and landscape matrix quality on wetland vertebrates: A meta-analysis. Ecol. Appl. 2015, 25, 812–825. [Google Scholar] [CrossRef]
- Bennett, A.F. Linkages in the Landscape: The Role of Corridors and Connectivity in Wildlife Conservation; IUCN: Gland, Switzerland, 1998. [Google Scholar]
- Forman, R.T.; Alexander, L.E. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef] [Green Version]
- Geneletti, D. Using spatial indicators and value functions to assess ecosystem fragmentation caused by linear infrastructures. Int. J. Appl. Earth Obs. Geoinf. 2004, 5, 1–15. [Google Scholar] [CrossRef]
- Moriyama, H. What Does it Mean to Protect Paddy Fields; Rural Culture Association Japan: Tokyo, Japan, 1997. [Google Scholar]
- Machado, I.F.; Maltchik, L. Can management practices in rice fields contribute to amphibian conservation in southern Brazilian wetlands? Aquat. Conserv. Mar. Freshw. Ecosyst. 2010, 20, 39–46. [Google Scholar] [CrossRef]
- Naito, R. Perspectives of conservation of pond-breeding frogs (focusing on the Nagoya Daruma pond frog) in rice paddy areas in Japan. Landsc. Ecol. 2012, 17, 57–73. [Google Scholar]
- Naito, R.; Sakai, M.; Morimoto, Y. Negative effects of deep roadside ditches on Pelophylax porosa brevipoda dispersal and migration in comparison with Hyla japonica in a rice paddy area in Japan. Zool. Sci. 2012, 29, 599–603. [Google Scholar] [CrossRef]
- Naito, R.; Sakai, M.; Natuhara, Y.; Morimoto, Y.; Shibata, S. Microhabitat use by Hyla japonica and Pelophylax porosa brevipoda at levees in rice paddy areas of Japan. Zool. Sci. 2013, 30, 386–391. [Google Scholar] [CrossRef]
- Borzée, A.; Jang, Y. Impact of rice and bean harvests on the Suweon Treefrog (Dryophytes suweonensis). Int. J. Curr. Res. 2017, 9, 59620–59623. [Google Scholar]
- Borzée, A.; Heo, K.; Jang, Y. Relationship between agroenvironmental variables and breeding Hylids in rice paddies. Sci. Rep. 2018, 8, 8049. [Google Scholar] [CrossRef] [PubMed]
- Groffen, J.; Borzée, A.; Jang, Y. Positioning of two treefrog species within rice paddies in relation to different habitat borders. Anim. Cells Syst. 2018, 22, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Borzée, A.; Jang, Y. Policy recommendation for the conservation of the Suweon Treefrog (Dryophytes suweonensis) in the Republic of Korea. Front. Environ. Sci. 2019, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Orchard, D.; Tessa, G.; Jehle, R. Age and growth in a European flagship amphibian: Equal performance at agricultural ponds and favourably managed aquatic sites. Aquat. Ecol. 2019, 53, 37–48. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Voucher ID | Species | Description |
|---|---|---|
| ZISP.2150 | Bombinator orientalis | Korea, 1898, 2 sp., Imperial Russian Geographical Society (perhaps, A. Zvegintsov; description of the trip in Zvegintsov (1900); his route—Tumangan River, Purenga, Musan, Peyktusan volcano, Amnoka River, Kange, Togurion-Miti pass, Chin’-Chan’-Gan’ River, Tsynampo). |
| ZISP.2151 | “Rana sp.” | Korea, 1898, 1sp. (lost), Imperial Russian Geographical Society (the same). |
| ZISP.2835 | Pelophylax nigromaculatus | Wonsan (=Genzan), Korea, 1900, 4 sp., P.Yu. Schmidt. |
| ZISP.2875 | Pelophylax nigromaculatus | Wonsan (=Genzan), Korea, 1900, 5 sp., P.Yu. Schmidt. |
| ZISP.2876 | Pelophylax nigromaculatus | Wonsan (=Genzan), Korea, 5 July 1900, 8 sp., P.Yu. Schmidt. |
| ZISP.2912 | Bombina orientalis | between Tonchen and Chogu-Chyen-Dogu Gulf, Korea, 1900, 4 sp., P.Yu. Schmidt |
| ZISP.2914 | Bufo stejnegeri | Korea, 1900, 1 sp., P.Yu. Schmidt (description of the trip in Schmidt (1900); his route—03–24 July, Wonsan—25 July, Anbion—Suchumde Lake—27 July, Tonchen—28 July, Chogu-Chyen-Dogu Gulf—30 July, Singesa monastery, Diamond Mountains—01 August, Uonjon termal springs—02 August, Uonjon’on Pass—03 August, Chaansa monastery—04 August, Peansa monastery—05 August, Ammudo-Koge pass—05–06 August, Yuchomsa monastery—07–09 August, Kosyon Town—10–12 August, Kansen—13 August, Oridin village—14 August, Naksans monastery—15 August, Yak-Yan Mount—16 August, Kannyn Town—17 August, Kusan village—18 August, Tegulien Pass—19–20 August, Ol’chansa monastery—21 August, Kvanmul’ River—22 August, Pkhen-Khan Town—23 August, Pkhen-Khan River—24 August, Chkhe-Chkhen Mount—25 August, Iyonmusa village—26 August, Changnim village—27–28 August, Shuakori village—29 August, Chu-Yon-Che pass—30 August, Iyochen’ Town—01 August, Andon Town—02 August, Chingpo Town—04 August, Iondo Town—Khynkhe Town—Kyonju Town—12 August, Fuzan City). |
| ZISP.2915 | Hynobius leechii | Korea (according to date—Ammudo-Koge pass), 05 August 1900, 2 sp., P.Yu. Schmidt. |
| ZISP.2925 | Bombina orientalis | Korea, 1900, 5 sp., P.Yu. Schmidt. |
| ZISP.3044 | Pelophylax nigromaculatus | rice fields in Wonsan (Genzan), Korea, 05 July 1900, 4 sp., P.Yu. Schmidt. |
| ZISP.13968–13981 | Rana uenoi | Korea, 14 sp., Chosun Academy of Science. |
| ZISP.13982–13990 | Pelophylax nigromaculatus | Korea, 9 sp., Chosun Academy of Science. |
| ZISP.13991–13995 | Pelophylax chosenicus | Korea, 5 sp., Chosun Academy of Science. |
| ZISP.13996–14000 | Glandirana emeljanovi | Korea, 5 sp., Chosun Academy of Science. |
| ZISP.14001–14009 | Dryophytes japonicus | Korea, 9 sp., Chosun Academy of Science. |
| ZISP.14010–14011 | Hynobius leechii | Korea, 2 sp., Chosun Academy of Science. |
| ZISP.14012 | Onychodactylus koreanus | Korea, 1 sp., Chosun Academy of Science. |
| ZISP.14013–14016 | Bufo gargarizans | Korea, 4 sp., Chosun Academy of Science. |
| ZISP.14017–14018 | Kaloula borealis | Korea, 2 sp., Chosun Academy of Science. |
| ZISP.14019–14021 | Dryophytessuweonensis | Pyongyang, Korea, 25–31 July 1947, 3 sp., V. Gnezdintsev. |
| ZISP.14022–14033 | Pelophylax nigromaculatus | Pyongyang, Korea, 25–31 July 1947, 12 sp., V. Gnezdintsev. |
| ZISP.14034–14035 | Rana uenoi | Pyongyang, Korea, 25–31 July 1947, 2 sp., V. Gnezdintsev. |
| ZISP.14036–14037 | Rana coreana | Pyongyang, Korea, 25–31 July 1947, 2 sp., V. Gnezdintsev. |
| ZISP.14038–14044 | Kaloula borealis | Pyongyang, Korea, 25–31 July 1947, 7 sp., V. Gnezdintsev. |
| ZISP.14045–14047 | Bombina orientalis | Eiko, Korea, 29 August 1947, 3 sp., V. Gnezdintsev. |
| ZMMGU.871 | Pelophylax nigromaculatus | Kheyduzuo [=Haeju], 6 October 1947, D. I. Bibikov, 1 sp. |
| ZMMGU.877 | Rana dybowskii | 8 October 1947, Kheyduzuo, D. I. Bibikov, 3 sp. |
| ZMMGU.878 | Strauchbufo raddei | 6 October 1947, Kheyduzuo, D. I. Bibikov, 6 sp. |
| ZMMGU.1141 | Pelophylax nigromaculatus | Hamhung, 25 September 1970, R. Bielawski, 1 sp. |
| Accession Number | Species | Locality | References |
|---|---|---|---|
| Dryophytes | |||
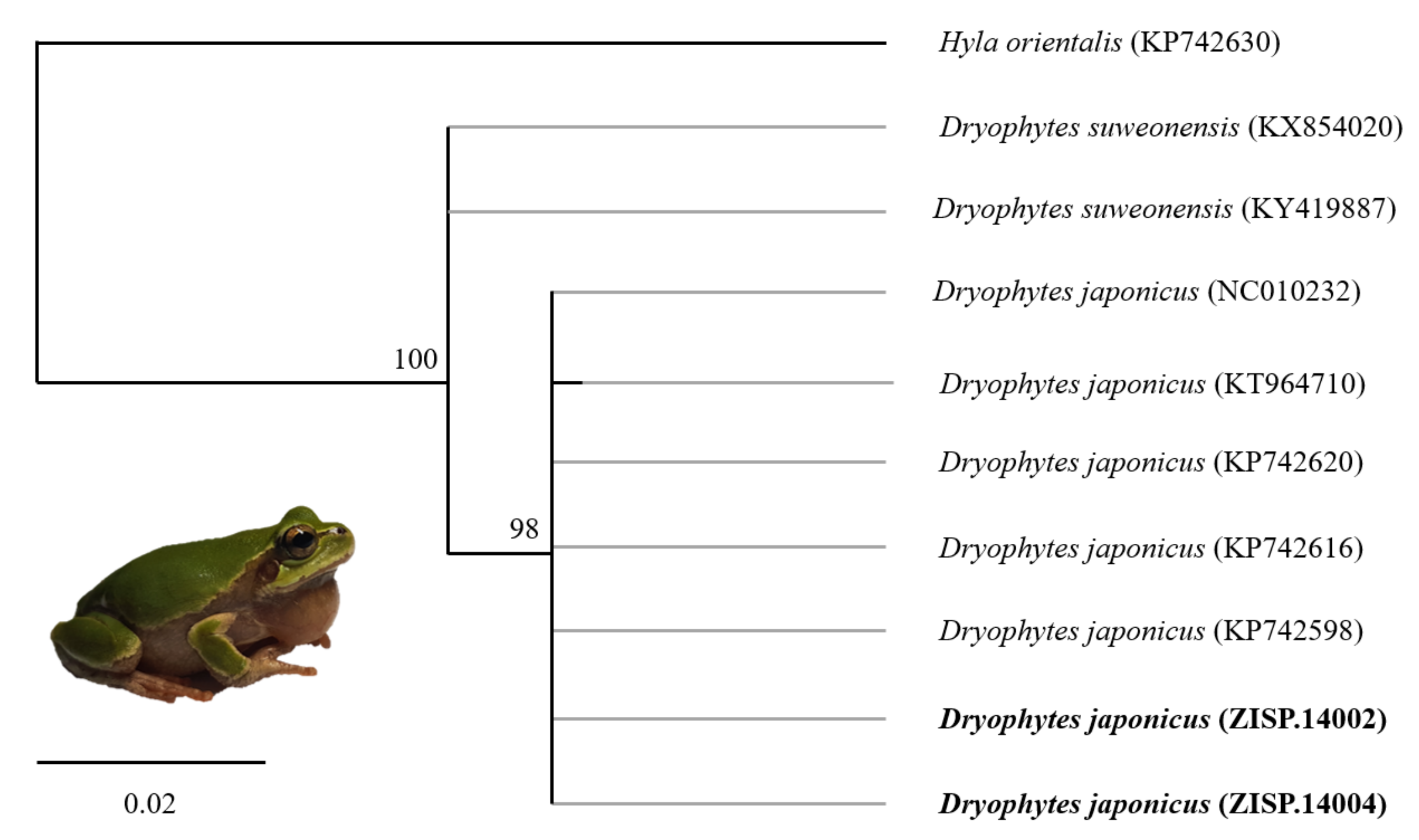
| KY419887 | Dryophytes suweonensis | Pyeongtaek, R Korea | [67] |
| KX854020 | Dryophytes suweonensis | Pyeongtaek, R Korea | [66] |
| KP742630 | Hyla orientalis | F. Russia | [65] |
| KP742620 | Dryophytes japonicus | Tsushima, Japan | |
| KP742616 | Dryophytes japonicus | Taksimo, F. Russia | |
| KP742598 | Dryophytes japonicus | Shenyang, PR China | |
| NC010232 | Dryophytes japonicus | Hiroshima, Japan | [64] |
| KT964710 | Dryophytes japonicus | Heilongjiang, PR China | [68] |
| Rana | |||
| KX024858 | Rana dybowskii | Huanran, PR China | [42] |
| KX024857 | Rana dybowskii | Fushun, PR China | [42] |
| KX024869 | Rana huanrenensis | Huanran, PR China | [42] |
| KU310893 | Rana kukunoris | PR China | |
| DQ289092 | Rana chensinensis | Huixian, PR China | [48] |
| KX024861 | Rana uenoi | Tsushima, Japan | [42] |
| KX024859 | Rana uenoi | Tsushima, Japan | [42] |
| KX024860 | Rana uenoi | Tsushima, Japan | [42] |
| KM876980 | Rana uenoi * | Boeun, R Korea | [42] |
| KM876979 | Rana uenoi * | Boeun, R Korea | [42] |
| AB058855 | Rana uenoi | Tsushima, Japan | [42] |
| KX139719 | Rana maoershanensis | Guangxi, PR China | [70] |
| KX024866 | Rana pirica | Hokkaido, Japan | [42] |
| KX024865 | Rana pirica | Hokkaido, Japan | [42] |
| AB058857 | Rana ornativentris | Aomori, Japan | [69] |
| AB058856 | Rana ornativentris | Hiroshima, Japan | [69] |
| Species | AUC ± SD | TSS ± SD |
|---|---|---|
| Bombina orientalis | 0.8651 ± 0.0116 | 0.4626 ± 0.0805 |
| Bufo gargarizans | 0.8617 ± 0.0199 | 0.4570 ± 0.0568 |
| Bufo stejnegeri | 0.9387 ± 0.0078 | 0.6905 ± 0.1167 |
| Dryophytes japonicus | 0.8459 ± 0.0155 | 0.4958 ± 0.0525 |
| Dryophytes suweonensis | 0.9637 ± 0.0064 | 0.8148 ± 0.0767 |
| Glandirana emeljanovi | 0.8877 ± 0.0367 | 0.5080 ± 0.1041 |
| Hynobius leechii | 0.8459 ± 0.0134 | 0.4538 ± 0.0544 |
| Kaloula borealis | 0.9158 ± 0.0142 | 0.6230 ± 0.1133 |
| Onychodactylus sp. | 0.9185 ± 0.0147 | 0.5924 ± 0.1078 |
| Onychodactylus koreanus | 0.9568 ± 0.0095 | 0.6962 ± 0.1314 |
| Pelophylax chosenicus | 0.9451 ± 0.0054 | 0.7137 ± 0.0810 |
| Pelophylax nigromaculatus | 0.8324 ± 0.0105 | 0.4085 ± 0.0518 |
| Strauchbufo raddei | 0.9881 ± 0.0052 | 0.9211 ± 0.0382 |
| Rana amurensis | 0.9104 ± 0.0091 | 0.6929 ± 0.0161 |
| Rana coreana | 0.8697 ± 0.0119 | 0.4652 ± 0.1158 |
| Rana dybowskii | 0.9268 ± 0.0280 | 0.5747 ± 0.1993 |
| Rana huanrenensis | 0.9913 ± 0.0030 | 0.8411 ± 0.1978 |
| Rana uenoi | 0.8869 ± 0.0079 | 0.5874 ± 0.0315 |
| Salamandrella tridactyla | 0.9716 ± 0.0143 | 0.4488 ± 0.0909 |
| Species | Population Dynamics | Reference |
|---|---|---|
| Bufo gargarizans | Common, reported from urban districts | [51,89] |
| Strauchbufo raddei | Uncommon or only locally abundant | [51,89] |
| Dryophytes japonicus | Present all over the nation | [51,89] |
| Bombina orientalis | Present all over the nation | [51,89] |
| Glandirana emeljanovi | One of the most common species in Central Korea | [50] |
| Hynobius leechii | Common in mountain forests | [51] |
| Onychodactylus | Common in mountain forests | [51] |
| Salamandrella tridactyla | Only in the high regions of the mountains | [51] |
| Pelophylax nigromaculatus | Abundant in rice paddies | [113] |
| Rana dybowskii-R. uenoi | Locally abundant | [113] |
| Bombina orientalis | Abundant in the lowlands | [113] |
| Bufo gargarizans | Abundant in the lowlands | [113] |
| Dryophytes japonicus | Abundant in the lowlands | [113] |
| Pelophylax chosenicus | Decline in abundance. Decline in North Pyongan province and Seoncheon because of droughts likely linked to climate change | W * |
| Rana coreana | Decline in abundance | W * |
| Bufo gargarizans: | High density on the west coast | W * |
| Glandirana emeljanovi | Not abundant but widespread | W * |
| Kaloula borealis | Continuous presence but not common, all the way to North Hamgyong province | W * |
| Pelophylax nigromaculatus | Most abundant amphibian species | W * |
| Strauchbufo raddei | Isolated populations in Shinuiju, Amrok River, Chilbeok mountain, North Pyongan province | W * |
| Rana dybowskii | Abundant at high altitude, populations >2100 m at Baekdu lake | W * |
| Hynobius leechii | Abundant <1000 m asl throughout its range | W * |
| Lithobates catesbeianus | Isolated large populations originating from escaped individuals from farms | W * |
| Rana sp. | Isolated large populations originating from escaped individuals from farms | W * |
| Species | Minimum Suitable Area (km2) | 10-Percentile Omission Area (km2) | IUCN Status | Recommendation at National Level | Local Name |
|---|---|---|---|---|---|
| Bombina orientalis | 119,531 | 30,729 | LC | LC | 비단개구리 |
| Bufo gargarizans | 101,627 | 29,954 | LC | LC | 두꺼비 |
| Bufo stejnegeri | 45,301 | 11,805 | LC | VU B1ab(i,ii,iii) | 금강두꺼비 |
| Dryophytes japonicus | 114,924 | 51,290 | LC | LC | 청개구리 |
| Dryophytes suweonensis | 25,214 | 9785 | EN | EN B1ab(i,ii,iii) | 수원청개구리 |
| Glandirana emeljanovi | 107,085 | 30,201 | LC | LC | 옴개구리 |
| Hynobius leechii | 60,541 | 41,443 | LC | LC | 도롱룡 |
| Kaloula borealis | 40,571 | 27,098 | LC | NT | 맹꽁이 |
| Onychodactylus sp. | 57,960 | 20,119 | |||
| Onychodactylus koreanus | 8637 | 6 | DD | VU B1ab(i,ii,iii) | 발톱도롱룡 |
| Pelophylax chosenicus | 35,908 | 13,810 | VU | VU B1ab(i,ii,iii) | 금개구리 |
| Pelophylax nigromaculatus | 102,328 | 51,779 | NT | NT | 참개구리 |
| Strauchbufo raddei | 22,026 | 7,775 | LC | VU B1ab(i,ii,iii) | 작은두꺼비 |
| Rana amurensis | 110,687 | 17,271 | LC | LC | 양용개구리 |
| Rana coreana | 54,740 | 26,249 | LC | NT | 애기개구리 |
| Rana dybowskii | 101,430 | 30,360 | LC | LC | 북방산개구리/기름개구리 |
| Rana huanrenensis | 78,533 | 34,021 | LC | LC | 계곡산개구리 |
| Rana uenoi | 36,066 | 9579 | NE | VU B1ab(i,ii,iii) | |
| Salamandrella tridactyla | 15,986 | 6294 | NE | NT | 합수도롱룡 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borzée, A.; Litvinchuk, S.N.; Ri, K.; Andersen, D.; Nam, T.Y.; Jon, G.H.; Man, H.S.; Choe, J.S.; Kwon, S.; Othman, S.N.; et al. Update on Distribution and Conservation Status of Amphibians in the Democratic People’s Republic of Korea: Conclusions Based on Field Surveys, Environmental Modelling, Molecular Analyses and Call Properties. Animals 2021, 11, 2057. https://doi.org/10.3390/ani11072057
Borzée A, Litvinchuk SN, Ri K, Andersen D, Nam TY, Jon GH, Man HS, Choe JS, Kwon S, Othman SN, et al. Update on Distribution and Conservation Status of Amphibians in the Democratic People’s Republic of Korea: Conclusions Based on Field Surveys, Environmental Modelling, Molecular Analyses and Call Properties. Animals. 2021; 11(7):2057. https://doi.org/10.3390/ani11072057
Chicago/Turabian StyleBorzée, Amaël, Spartak N. Litvinchuk, Kyongsim Ri, Desiree Andersen, Tu Yong Nam, Gwang Hyok Jon, Ho Song Man, Jong Sik Choe, Sera Kwon, Siti N. Othman, and et al. 2021. "Update on Distribution and Conservation Status of Amphibians in the Democratic People’s Republic of Korea: Conclusions Based on Field Surveys, Environmental Modelling, Molecular Analyses and Call Properties" Animals 11, no. 7: 2057. https://doi.org/10.3390/ani11072057
APA StyleBorzée, A., Litvinchuk, S. N., Ri, K., Andersen, D., Nam, T. Y., Jon, G. H., Man, H. S., Choe, J. S., Kwon, S., Othman, S. N., Messenger, K., Bae, Y., Shin, Y., Kim, A., Maslova, I., Luedtke, J., Hobin, L., Moores, N., Seliger, B., ... Jang, Y. (2021). Update on Distribution and Conservation Status of Amphibians in the Democratic People’s Republic of Korea: Conclusions Based on Field Surveys, Environmental Modelling, Molecular Analyses and Call Properties. Animals, 11(7), 2057. https://doi.org/10.3390/ani11072057