
High Granulocyte-Macrophage Colony Stimulating Factor to Interleukin 10 Ratio and Marked Antioxidant Enzyme Activities Predominate in Symptomatic Cattle Naturally Infected with Candidatus Mycoplasma haemobos, Theileria orientalis, Theileria sinensis and Trypanosoma evansi

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Blood Sample Collection
2.3. DNA Extraction
2.4. Detection of Blood Parasites by Conventional PCR
2.5. Quantification of Serum Inflammatory Cytokines in Cattle
2.6. Oxidant and Antioxidant Assays
2.7. Statistical Analysis
3. Results
3.1. Molecular Detection Rate of Blood Parasites in the Symptomatic and Asymptomatic Cattle
3.2. Serum Proinflammatory, Antiinflammatory Cytokine and Oxidant/Antioxidant Levels in Symptomatic and Asymptomatic Cattle
3.2.1. Serum Cytokine and Oxidant/Antioxidant Levels in Symptomatic and Asymptomatic Cattle Naturally Infected with Candidatus Mycoplasma haemobos
3.2.2. Serum Cytokine Levels and Oxidant/Antioxidant Levels in Symptomatic and Asymptomatic Cattle Naturally Infected by Theileria orientalis
3.2.3. Serum Cytokine and Oxidant/Antioxidant Levels in Symptomatic and Asymptomatic Cattle Naturally Infected with Anaplasma marginale
3.2.4. Serum Cytokine Levels in Symptomatic and Asymptomatic Cattle Naturally Infected with Theileria sinensis
3.2.5. Serum Cytokine and Oxidant/Antioxidant Levels in Symptomatic Cattle Naturally Infected with Trypanosoma evansi
3.2.6. Serum Cytokine Levels and Oxidant/Antioxidant Levels in Symptomatic Cattle Naturally Infected with Anaplasma platys
3.3. Pro-Inflammatory and Anti-Inflammatory Cytokine Ratios in Symptomatic and Asymptomatic Cattle Naturally Infected with Blood Parasites
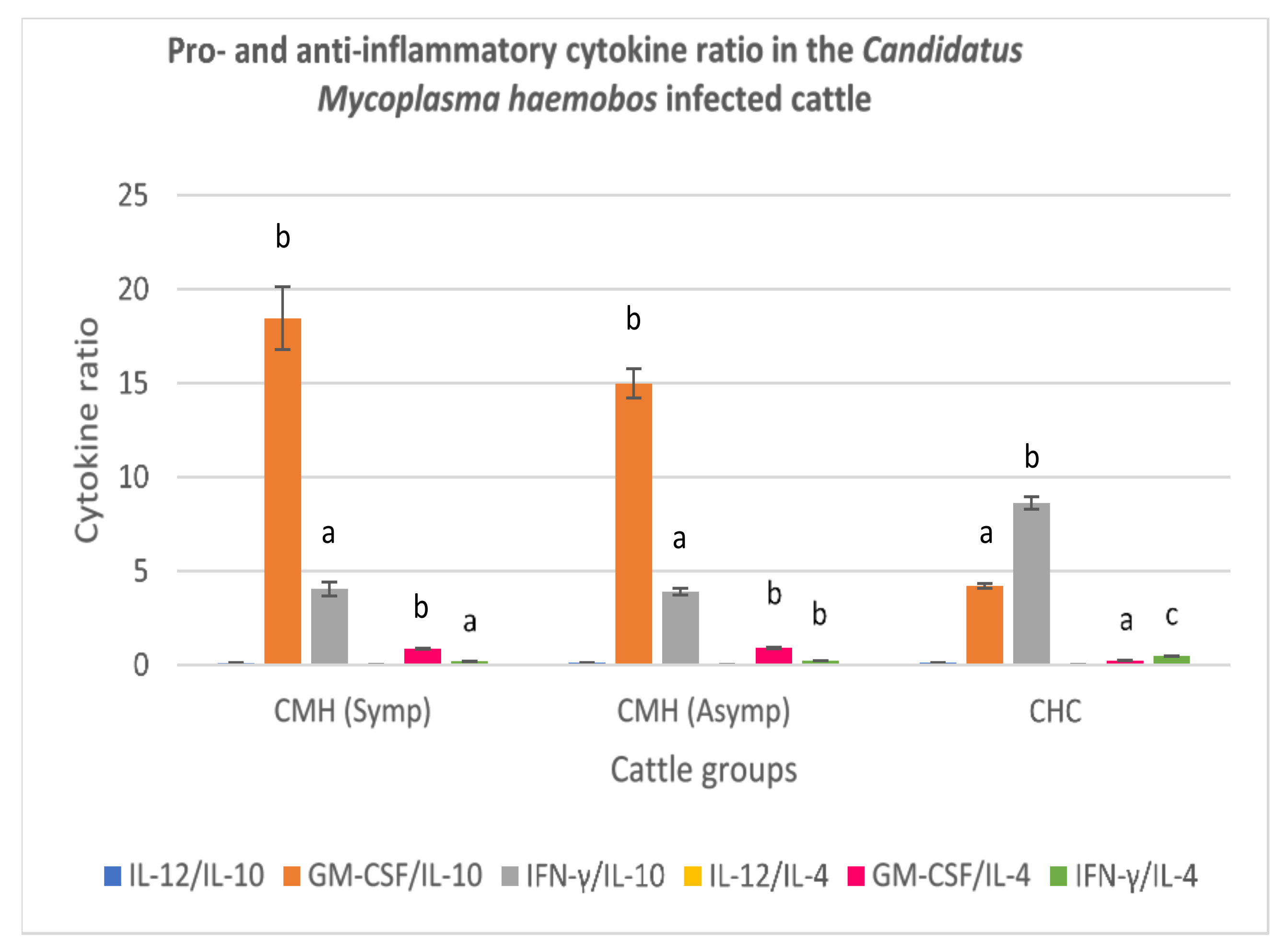
3.3.1. Candidatus Mycoplasma haemobos-Infected Cattle
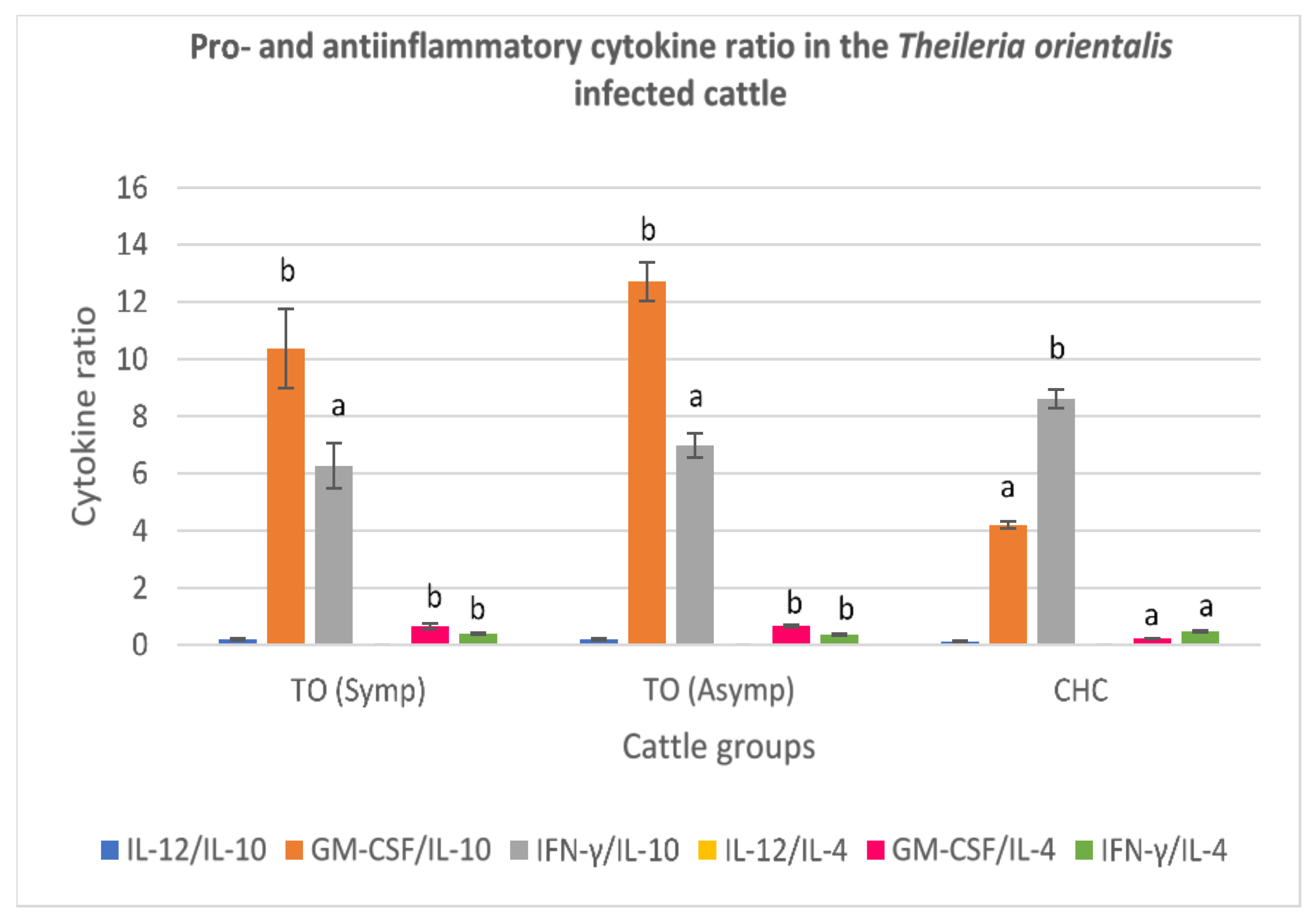
3.3.2. Theileria orientalis-Infected Cattle
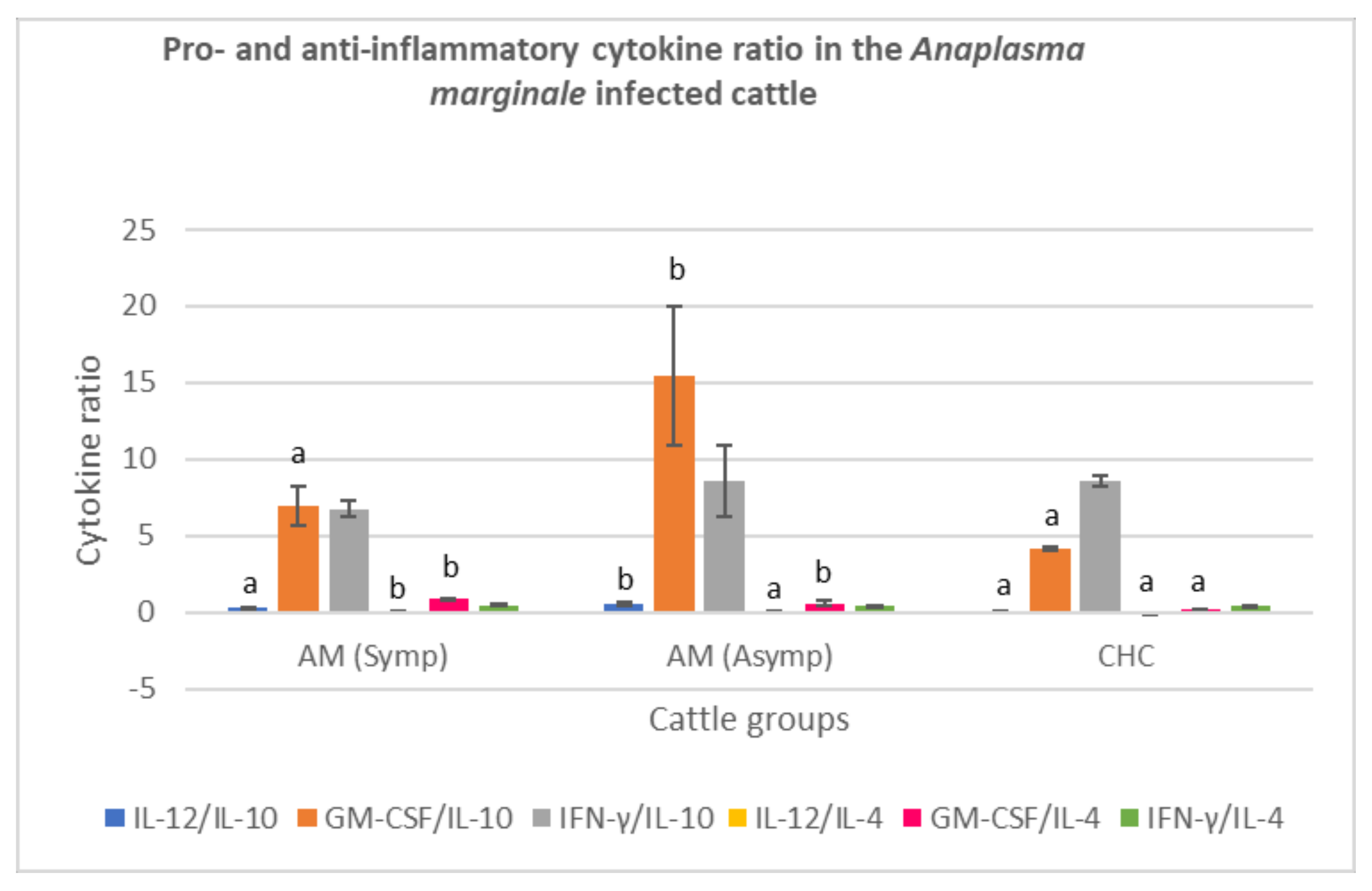
3.3.3. Anaplasma marginale-Infected Cattle
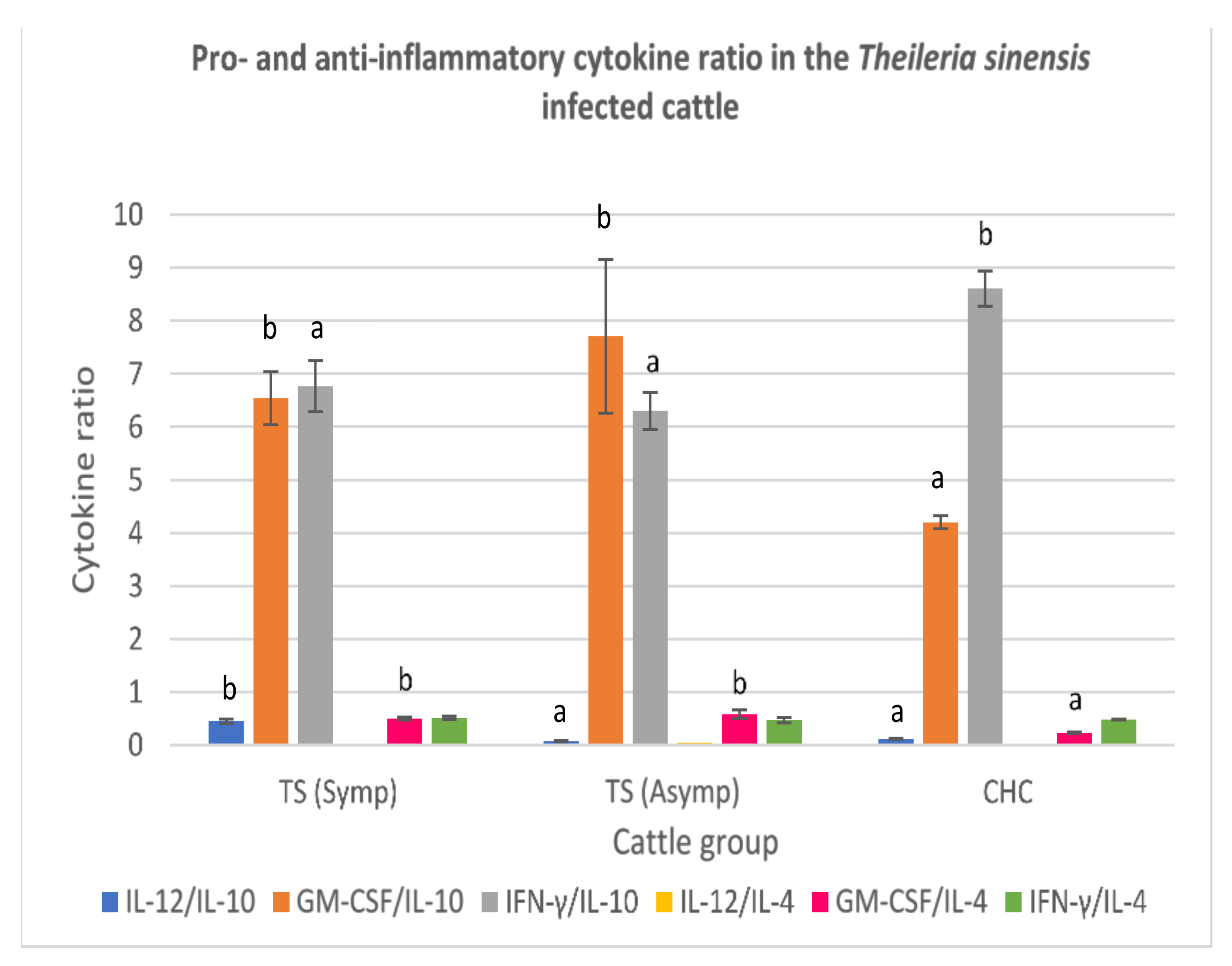
3.3.4. Theileria sinensis-Infected Cattle
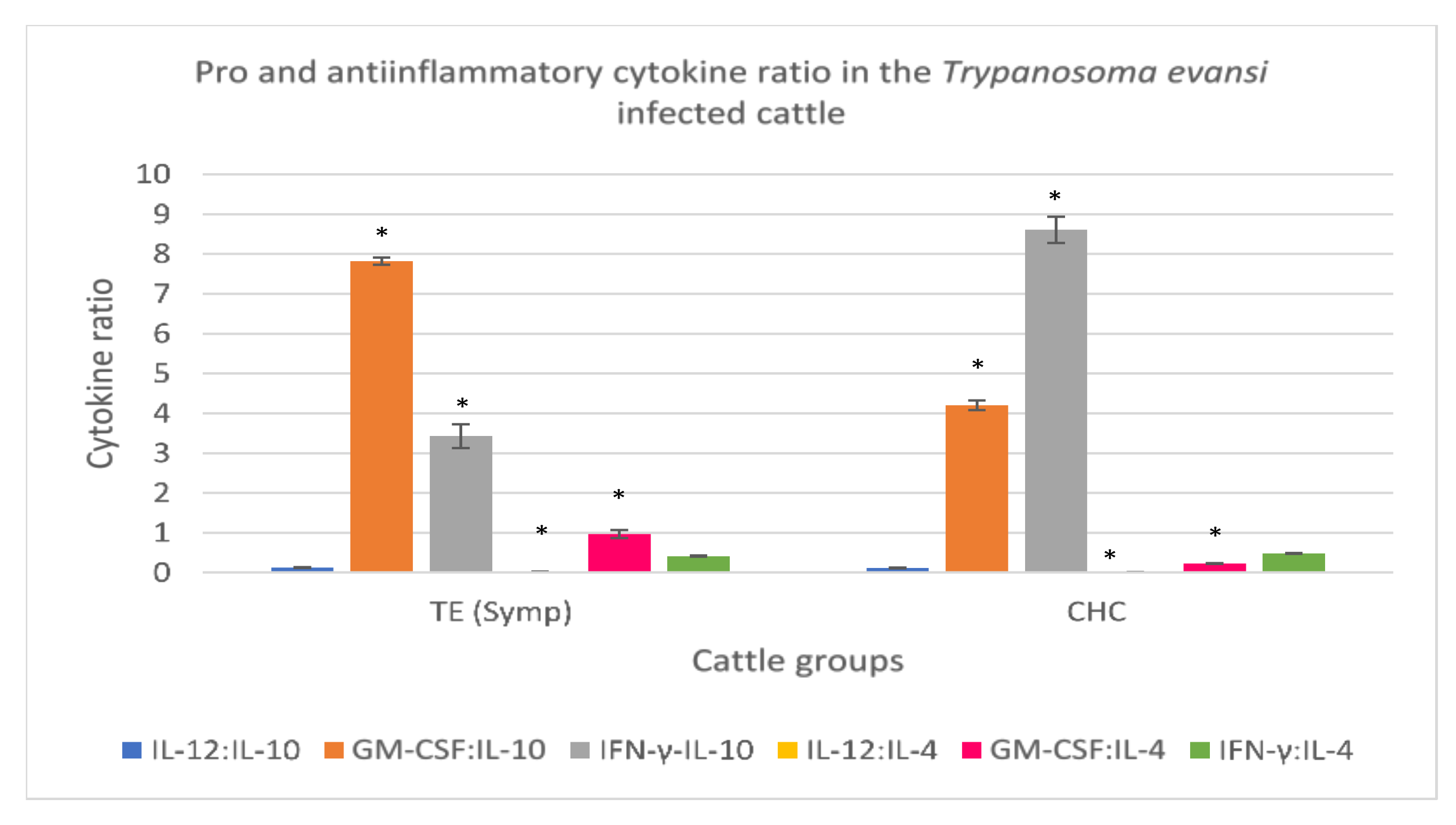
3.3.5. Trypanosoma evansi-Infected Cattle
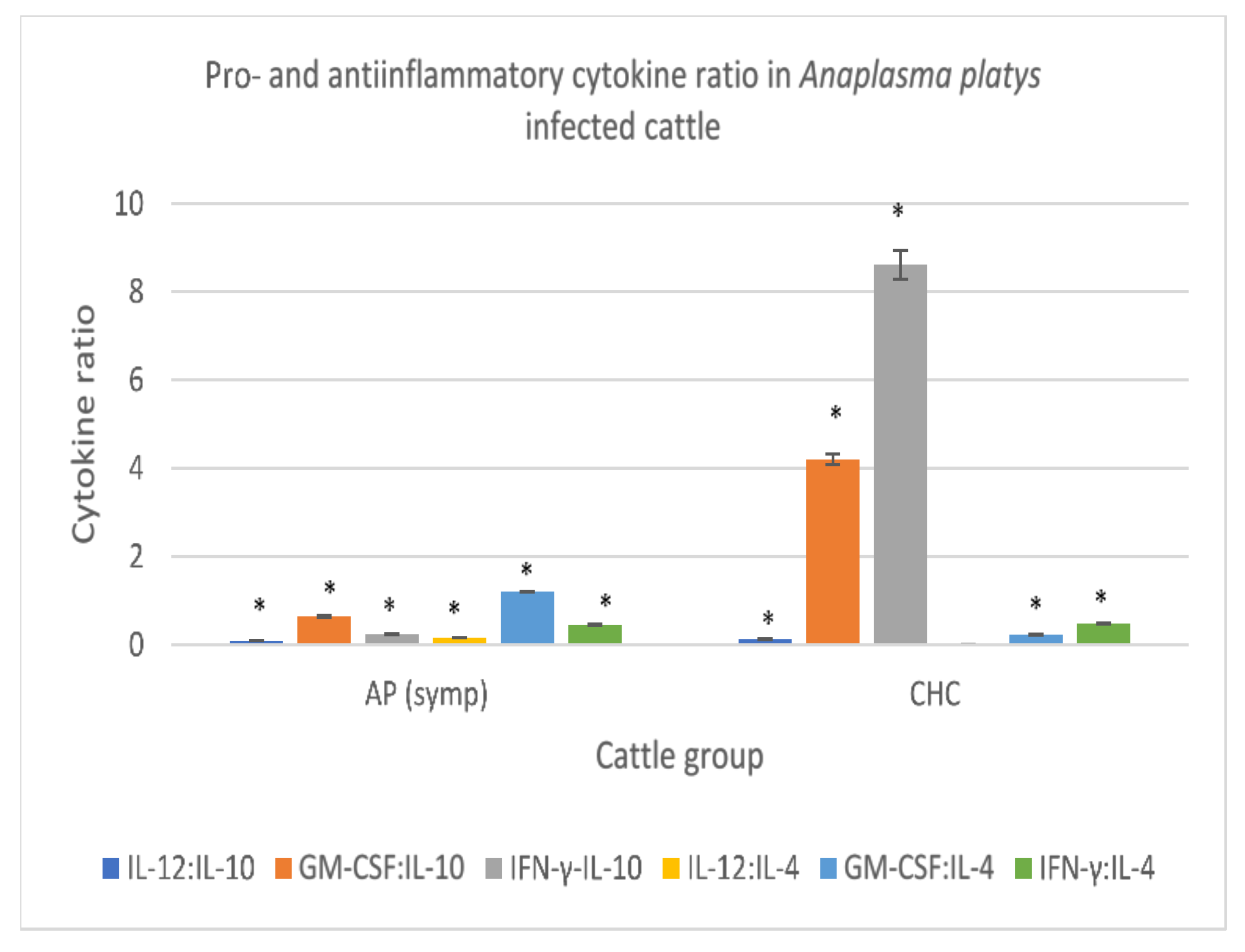
3.3.6. Anaplasma platys-Infected Cattle
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adjou Moumouni, P.F.; Aboge, G.O.; Terkawi, M.A.; Masatani, T.; Cao, S.; Kamyingkird, K.; Jirapattharasate, C.; Zhou, M.; Wang, G.; Liu, M.; et al. Molecular detection and characterization of Babesia bovis, Babesia bigemina, Theileria species and Anaplasma marginale isolated from cattle in Kenya. Parasites Vectors 2015, 8, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilgic, H.B.; Bakırcı, S.; Kose, O.; Unlu, A.H.; Hacılarlıoglu, S.; Eren, H.; Weir, W.; Karagenc, T. Prevalence of tick-borne haemoparasites in small ruminants in Turkey and diagnostic sensitivity of single-PCR and RLB. Parasites Vectors 2017, 10, 1–13. [Google Scholar] [CrossRef]
- Roland, L.; Drillich, M.; Iwersen, M. Hematology as a diagnostic tool in bovine medicine. J. Vet. Diagn. Investig. 2014, 26, 592–598. [Google Scholar] [CrossRef] [Green Version]
- Agina, O.A.; Mohd Rosly, S.; Nur Mahiza, I.; Mokrish, A.; Zamri-Saad, M.; Mazlina, M.; Azim-Salahuddin, M.; Afrah, A.K.; Lee, C.H.; Fairuz, H.R.; et al. First report of bovine anaemia associated Theileria sinensis infection and phylogenetic analyses of partial gene sequences of Theileria and Anaplasma species detected in naturally infected Malaysian cattle. Parasites Vectors 2020. Preprint. [Google Scholar] [CrossRef] [Green Version]
- Shebish, E.; Vemulapalli, R.; Oseto, C. Prevalence and molecular detection of Anaplasma marginale, Babesia bovis and Babesia bigemina in cattle from Puntarenas Province, Costa Rica. Vet. Parasitol. 2012, 188, 164–167. [Google Scholar] [CrossRef]
- Bakari, S.M.; Ofori, J.A.; Kusi, K.A.; Aning, G.K.; Awandare, G.A.; Carrington, M.; Gwira, T.M. Serum biochemical parameters and cytokine profiles associated with natural African trypanosome infections in cattle. Parasites Vectors 2017, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bochniarz, M.; Zdzisińska, B.; Wawron, W.; Szczubiał, M.; Dąbrowski, R. Milk and serum IL-4, IL-6, IL-10, and amyloid A concentrations in cows with subclinical mastitis caused by coagulase-negative staphylococci. J. Dairy Sci. 2017, 100, 9674–9680. [Google Scholar] [CrossRef] [Green Version]
- Hou, R.; Garner, M.; Holmes, C.; Osmond, C.; Teeling, J.; Lau, L.; Baldwin, D.S. Peripheral inflammatory cytokines and immune balance in Generalised Anxiety Disorder: Case-controlled study. Brain Behav. Immun. 2017, 62, 212–218. [Google Scholar] [CrossRef] [Green Version]
- Syed, Z.I.H.; Sharma, N.; Singh, A.K.; Pathak, A.K.; Bacic, I.; Bacic, G.; Macesic, N.; Jeong, D.K. Inflammatory cytokines, helping tool in veterinary diagnostics: From basics to advancement. Vet. Pract. 2019, 20, 159–162. [Google Scholar]
- Jain, S.; Gautam, V.; Naseem, S. Acute-phase proteins: As diagnostic tool. J. Pharm. Bioallied Sci. 2011, 3, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [Green Version]
- Dąbrowski, R.; Pastor, J.; Szczubiał, M.; Piech, T.; Bochniarz, M.; Wawron, W.; Tvarijonaviciute, A. Serum IL-6 and IL-10 concentrations in bitches with pyometra undergoing ovariohysterectomy. Acta Vet. Scand. 2015, 57, 61. [Google Scholar] [CrossRef] [Green Version]
- Baral, T.N. Immunobiology of African Trypanosomes: Need of Alternative Interventions. J. Biomed. Biotechnol. 2010, 2010, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Liu, C.H.; Roberts, A.I.; Das, J.; Xu, G.; Ren, G.; Zhang, Y.; Zhang, L.; Yuan, Z.R.; Tan, H.S.W.; et al. Granulocyte-macrophage colony-stimulating factor (GM-CSF) and T-cell responses: What we do and don’t know. Cell Res. 2006, 16, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Ushach, I.; Zlotnik, A. Biological role of granulocyte macrophage colony-stimulating factor (GM-CSF) and macrophage colony-stimulating factor (M-CSF) on cells of the myeloid lineage. J. Leukoc. Biol. 2016, 100, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.A. Colony-stimulating factors in inflammation and autoimmunity. Nat. Rev. Immunol. 2008, 8, 533–544. [Google Scholar] [CrossRef]
- Francisco-Cruz, A.; Aguilar-Santelises, M.; Ramos-Espinosa, O.; Mata-Espinosa, D.; Marquina-Castillo, B.; Barrios-Payan, J.; Hernandez-Pando, R. Granulocyte-macrophage colony-stimulating factor: Not just another haematopoietic growth factor. Med. Oncol. 2014, 31, 774. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Gadina, M.; Siegel, R.M. Cytokines and Cytokine Receptors, 5th ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2019; Volume 1. [Google Scholar]
- Zhang, M.-Z.; Yao, B.; Yang, S.; Jiang, L.; Wang, S.; Fan, X.; Yin, H.; Wong, K.; Miyazawa, T.; Chen, J.; et al. CSF-1 signaling mediates recovery from acute kidney injury. J. Clin. Investig. 2012, 122, 4519–4532. [Google Scholar] [CrossRef] [Green Version]
- Gasson, J.C. Molecular physiology of granulocyte-macrophage colony-stimulating factor. Blood 1991, 77, 1131–1145. [Google Scholar] [CrossRef]
- Gangadhar, T.C.; Vonderheide, R.H. Mitigating the toxic effects of anticancer immunotherapy. Nat. Rev. Clin. Oncol. 2014, 11, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Loisa, P.; Rinne, T.; Laine, S.; Hurme, M.; Kaukinen, S. Anti-inflammatory cytokine response and the development of multiple organ failure in severe sepsis. Acta Anaesthesiol. Scand. 2003, 47, 319–325. [Google Scholar] [CrossRef]
- El-Sebaei, M.; El-Ashker, M.; El-Boshy, M. The role of acute phase cytokines in the recovery and disease progress of Theileria annulata-infected cattle. Comp. Clin. Path. 2014, 23, 1497–1502. [Google Scholar] [CrossRef]
- El-Ashker, M.; Salama, M.; El-Sebaei, M.; Risha, E.; Abdelhamid, F.; El-Diasty, M.; El-Fadle, E. Significance of clinical variables and selected biochemical markers in predicting the outcome of bovine anaplasmosis. Vet. Med. 2015, 60, 301–308. [Google Scholar] [CrossRef]
- Fundu, T.M. Subcellular Localization of Glutathione Peroxidase, Change in Glutathione System during Ageing and Effects on Cardiometabolic Risks and Associated Diseases. In Glutathione System and Oxidative Stress in Health and Disease; Kapepula, P.M., Ed.; IntechOpen: Rijeka, Croatia, 2020; Chapter 2; ISBN 9781838801267. [Google Scholar]
- Younus, H. Therapeutic potentials of superoxide dismutase. Int. J. Health Sci. 2018, 12, 88–93. [Google Scholar]
- Şimşek, S.; Yüce, A.; Ütük, A.E. Determination of serum malondialdehyde levels in sheep naturally infected with Dicrocoelium dendriticum. Saglık Bilim. Dergisi Fırat Üniversitesi 2006, 20, 217–220. [Google Scholar]
- Bai, Q.; Liu, G.; Yin, H.; Zhao, Q.; Liu, D.; Ren, J.; Li, X. Theileria sinensis sp nov: A new species of bovine Theileria—Molecular taxonomic studies. Acta Vet. Zootech. Sinica 2002, 33, 185–190. [Google Scholar]
- Shkap, V.; Leibovitz, B.; Krigel, Y.; Molad, T.; Fish, L.; Mazuz, M.; Fleiderovitz, L.; Savitsky, I. Concomitant infection of cattle with the vaccine strain Anaplasma marginale ss centrale and field strains of A. marginale. Vet. Microbiol. 2008, 130, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Ota, N.; Mizuno, D.; Kuboki, N.; Igarashi, I.; Nakamura, Y.; Yamashina, H.; Hanzaike, T.; Fujii, K.; Onoe, S.; Hata, H.; et al. Epidemiological survey of Theileria orientalis infection in grazing cattle in the eastern part of Hokkaido, Japan. J. Vet. Med. Sci. 2009, 71, 937–944. [Google Scholar] [CrossRef] [Green Version]
- Urakawa, T.; Claes, F.; Radwanska, M.; Majiwa, P.A.O.; Goddeeris, B.; Büscher, P. Variable surface glycoprotein RoTat 1.2 PCR as a specific diagnostic tool for the detection of Trypanosoma evansi infections. Kinetoplastid Biol. Dis. 2004, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.L.; Song, H.Q.; Lin, R.Q.; Yuan, Z.G.; Yang, J.F.; Zhao, G.H.; Huang, W.Y.; Zhu, X.Q. The detection of “Candidatus Mycoplasma haemobos” in cattle and buffalo in China. Trop. Anim. Health Prod. 2010, 42, 1805–1808. [Google Scholar] [CrossRef] [PubMed]
- Inokuma, H.; Fujii, K.; Matsumoto, K.; Okuda, M.; Nakagome, K.; Kosugi, R.; Hirakawa, M.; Onishi, T. Demonstration of Anaplasma (Ehrlichia) platys inclusions in peripheral blood platelets of a dog in Japan. Vet. Parasitol. 2002, 110, 145–152. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Sousa, G.R.; Gomes, J.A.S.; Fares, R.C.G.; de Souza Damásio, M.P.; Chaves, A.T.; Ferreira, K.S.; Nunes, M.C.P.; Medeiros, N.I.; Valente, V.A.A.; Corrêa-Oliveira, R.; et al. Plasma cytokine expression is associated with cardiac morbidity in chagas disease. PLoS ONE 2014, 9, e87082. [Google Scholar] [CrossRef] [PubMed]
- Uppal, S.S.; Raghupathy, R.; Hayat, S.J.; Longenecker, J.C.; Abraham, M.; Rawoot, P. Disease activity and cytokine production in mitogen-stimulated peripheral blood mononuclear cells from patients with rheumatoid arthritis. Med. Princ. Pract. 2010, 19, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.P.; Brown, D.J.; Vatansever, Z.; Waddington, D.; Taylor, L.H.; Nichani, A.K.; Campbell, J.D.; Adamson, R.E.; Glass, E.J.; Spooner, R.L. Proinflammatory cytokine expression by Theileria annulata infected cell lines correlates with the pathology they cause in vivo. Vaccine 2001, 19, 2932–2944. [Google Scholar] [CrossRef]
- Simmons, E.M.; Himmelfarb, J.; Sezer, M.T.; Chertow, G.M.; Mehta, R.L.; Paganini, E.P.; Soroko, S.; Freedman, S.; Becker, K.; Spratt, D.; et al. Plasma cytokine levels predict mortality in patients with acute renal failure. Kidney Int. 2004, 65, 1357–1365. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Kellum, J.A. Interleukin-6. Crit. Care Med. 2005, 33, S463–S465. [Google Scholar] [CrossRef]
- Petricevich, V.L. Balance Between Pro- and Anti-Inflammatory Cytokines in Mice Treated With Centruroides noxius Scorpion Venom. Mediat. Inflamm. 2006, 2006, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agina, O.A. Haematology and Clinical Biochemistry Findings Associated with Equine Diseases—A Review. Not. Sci. Biol. 2017, 9, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Shiono, H.; Yagi, Y.; Chikayama, Y.; Miyazaki, S.; Nakamura, I. Oxidative damage and phosphatidylserine expression of red blood cells in cattle experimentally infected with Theileria sergenti. Parasitol. Res. 2003, 89, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.R.; Senapati, S.K.; Sahoo, S.C. Trypanosomiasis induced oxidative stress and hemato-biochemical alteration in cattle. J. Entomol. Zool. Stud. 2017, 5, 721–727. [Google Scholar]
- Parashar, R.; Singla, L.D.; Gupta, M.; Sharma, S.K. Evaluation and correlation of oxidative stress and haemato-biochemical observations in horses with natural patent and latent trypanosomosis in Punjab state of India. Acta Parasitol. 2018, 63, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Sulaiman, F.A. Biochemical and morphological changes in Trypanosoma brucei brucei-infected rats treated with homidium chloride and diminazene aceturate. J. Basic Clin. Physiol. Pharmacol. 2012, 23, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Esmaeilnejad, B.; Tavassoli, M.; Samiei, A.; Hajipour, N.; Imani-Baran, A.; Farhang-Pajuh, F. Evaluation of oxidative stress and antioxidant status, serum trace mineral levels and cholinesterases activity in cattle infected with Anaplasma marginale. Microb. Pathog. 2018, 123, 402–409. [Google Scholar] [CrossRef]
- Asri Rezaei, S.; Dalir-Naghadeh, B. Evaluation of antioxidant status and oxidative stress in cattle naturally infected with Theileria annulata. Vet. Parasitol. 2006, 142, 179–186. [Google Scholar] [CrossRef]
- Hassanpour, A.; Sabegh, Y.G.; Sadeghi-nasab, A.; Hassanpour, A. Assessment of serum antioxidant enzymes activity in cattle suffering from Theileriosis. Eur. J. Exp. Biol. 2013, 3, 493–496. [Google Scholar]
- Singh, R.; Bhardwaj, R.K.; Gupta, A.K. Mixed Babesia Canis and Anaplasma Platys/Phagocytophilum Infection in Dog. Dairy Vet. Sci. J. 2018, 7, 555707. [Google Scholar] [CrossRef] [Green Version]
- Abd Ellah, M.R. Involvement of free radicals in parasitic infestations. J. Appl. Anim. Res. 2013, 41, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Grewal, A.; Ahuja, C.S.; Singha, S.P.S.; Chaudhary, K.C. Status of Lipid Peroxidation, Some Antioxidant Enzymes and Erythrocytic Fragility of Crossbred Cattle Naturally Infected with Theileria annulata. Vet. Res. Commun. 2005, 29, 387–394. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD ), catalase (CAT ) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood Parasites | Gene of Interest | Primer Sequences (5′–3′) Forward (F) and Reverse (R) | Thermocycling Conditions | Base Pair (bp) | References |
|---|---|---|---|---|---|
| T. sinensis | MPSP | F-CACTGCTATGTTGTCCAAGAGATATT R- AATGCGCCTAAAGATAGTAGAAAAC | ID: 94 °C/3 min D: 94 °C/1 min A: 56 °C/1 min E: 72 °C/1 min No. of cycles:40 FE: 72 °C/7 min | 887 | [28] |
| A. marginale | MSP4 | F- CATCTCCCATGAGTCACGAAGTGGC R- GCTGAACAGGAATCTTGCTCCAAG | ID: 95 °C/5 min D: 95 °C/1 min A: 65 °C/2 min E: 72 °C/1 min No. of cycles:40 FE: 72 °C/10 min | 761 | [29] |
| T. orientalis | MPSP | F- CTTTGCCTAGGATACTTCCT R -ACGGCAAGTGGTGAGAACT | ID: 94 °C/4 min D: 94 °C/1 min A: 63 °C/1 min E: 72 °C/1 min No. of cycles:40 FE: 72 °C/7 min | 776 | [30] |
| T. evansi | RoTaT 1.2 VSG | F- 5′-GCGGGGTGTTTAAAGCAATA-3′ R- 5′-ATTAGTGCTGCGTGTGTTCG-3′ | ID: 94 °C/4 min D: 94 °C/1 min A: 59 °C/1 min E: 72 °C/1 min No. of cycles:40 FE: 72 °C/5 min | 208 | [31] |
| Candidatus Mycoplasma haemobos | 16 SrRNA | F- GAGTTAGTTATTAAAGCTTTAT R- ATTCATGAGGTACTATCAGTTG | ID: 94 °C/5 min D: 94 °C/30 s A: 55 °C/30 s E: 72 °C/30 s No. of cycles:40 FE: 72 °C/7 min | 279 | [32] |
| A. platys | 16SrRNA | PLATYS F- AAGTCGAACGGATTTTTGTC PLATYS R -CTTTAACTTACCGAACC | ID: 95 °C/5 min D: 95 °C/30 s A: 55 °C/30 s E: 72 °C/90 s No. of cycles:40 FE: 72 °C/5 min | 504 | [33] |
| Blood Parasites | No. of Cattle | Occurrence (%) (95% Confidence Interval) |
|---|---|---|
| Candidatus Mycoplasma haemobos | 70 | 53.9 (45.3–62.2) |
| Theileria orientalis | 65 | 50 (41.5–58.5) |
| Anaplasma marginale | 49 | 37.8 (29.8–46.3) |
| Theileria sinensis | 32 | 24.6 (18.0–32.3) |
| Trypanosoma evansi | 4 | 3.08 (1.20–7.64) |
| Anaplasma platys | 7 | 5.38 (2.63–10.7) |
| Proinflammatory/ Anti-Inflammatory Cytokines | C. Mycoplasma haemobos Infected Cattle (Symptomatic) (n = 3) | C. Mycoplasma haemobos Infected Cattle (Asymptomatic) (n = 12) | Clinically Healthy Cattle (n = 8) |
|---|---|---|---|
| Interleukin 12 (ng/L) | 4.11 ± 0.05 b | 5.71 ± 1.19 c | 2.36 ± 0.08 a |
| Interleukin 10 (ng/L) | 40.78 ± 3.87 b | 50.45 ± 3.27 b | 19.53 ± 0.43 a |
| GM-CSF (pg/mL) | 745.73 ± 3.02 b | 717.75 ± 2.81 b | 81.68 ± 1.26 a |
| Interleukin 4 (ng/L) | 864.70 ± 13.36 c | 795.64 ± 12.74 b | 349.85 ± 3.05 a |
| Interferon-γ (pg/mL) | 163.18 ± 1.05 a | 184.50 ± 5.26 b | 167.42 ± 4.38 a |
| Oxidant/antioxidant Markers | |||
| Malondialdehyde (nM/mL) | 3.70 ± 1.60 c | 2.51 ± 0.16 b | 1.37 ± 0.02 a |
| Superoxide dismutase (U/mL) | 945.36 ± 4.87 b | 977.16 ± 9.72 b | 546.6 ± 4.64 a |
| Glutathione peroxidase (U/mL) | 138.24 ± 2.10 b | 138.63 ± 1.59 b | 54.40 ± 0.45 a |
| Proinflammatory/ Anti-Inflammatory Cytokines | T. orientalis Infected Cattle (Symptomatic) (n = 4) | T. orientalis Infected Cattle (Asymptomatic) (n = 9) | Clinically Healthy Cattle (n = 8) |
|---|---|---|---|
| Interleukin 12 (ng/L) | 5.83 ± 0.53 b | 5.20 ± 0.24 b | 2.36 ± 0.08 a |
| Interleukin 10 (ng/L) | 45.87 ± 5.16 b | 38.60 ± 2.41 b | 19.53 ± 0.43 a |
| GM-CSF (pg/mL) | 454.55 ± 8.74 b | 473.88 ± 8.97 b | 81.68 ± 1.26 a |
| Interleukin 4 (ng/L) | 689.50 ± 10.34 b | 705.15 ± 11.64 b | 349.85 ± 3.05 a |
| Interferon-γ (pg/mL) | 276.97 ± 16.35 b | 261.15 ± 10.87 b | 167.42 ± 4.38 a |
| Oxidant/antioxidant markers | |||
| Malondialdehyde (nM/mL) | 3.97 ± 0.93 b | 3.01 ± 0.45 b | 1.37 ± 0.02 a |
| Superoxide dismutase (U/mL) | 854.31 ± 20.83 b | 921.80 ± 21.77 b | 546.6 ± 4.64 a |
| Glutathione peroxidase (U/mL) | 188.83 ± 6.64 b | 186.00 ± 3.97 b | 54.40 ± 0.45 a |
| Proinflammatory/ Anti-Inflammatory Cytokines | Anaplasma marginale Infected Cattle (Symptomatic) (n = 3) | Anaplasma marginale Infected Cattle (Asymptomatic) (n = 3) | Clinically Healthy Cattle (n = 8) |
|---|---|---|---|
| Interleukin 12 (ng/L) | 20.03 ± 1.80 b | 37.35 ± 1.12 c | 2.36 ± 0.08 a |
| Interleukin 10 (ng/L) | 39.50 ± 3.18 a | 342.53 ± 12.41 b | 19.53 ± 0.43 a |
| GM-CSF (pg/mL) | 515.62 ± 12.41 b | 517.80 ± 19.76 b | 81.68 ± 1.26 a |
| Interleukin 4 (ng/L) | 749.66 ± 9.66 b | 964.86 ± 4.35 b | 349.85 ± 3.05 a |
| Interferon-γ (pg/mL) | 291.96 ± 29.48 b | 373.91 ± 28.045 c | 167.42 ± 4.38 a |
| Oxidant/Antioxidant Markers | |||
| Malondialdehyde (nM/mL) | 3.88 ± 1.81 b | 2.18 ± 0.05 a | 1.37 ± 0.02 a |
| Superoxide dismutase (U/mL) | 1236.33 ± 10.88 b | 932.99 ± 17.24 b | 546.6 ± 4.64 a |
| Glutathione peroxidase (U/mL) | 145.20 ± 1.12 b | 277.74 ± 9.49 c | 54.40 ± 0.45 a |
| Proinflammatory/ Anti-Inflammatory Cytokines | Theileria sinensis Infected Cattle (Symptomatic) (n = 3) | Theileria sinensis Infected Cattle (Asymptomatic) (n = 3) | Clinically Healthy Cattle (n = 8) |
|---|---|---|---|
| Interleukin 12 (ng/L) | 17.88 ± 3.35 b | 3.50 ± 0.66 a | 2.36 ± 0.08 a |
| Interleukin 10 (ng/L) | 52.36 ± 1.60 c | 40.92 ± 1.71 b | 19.53 ± 0.43 a |
| GM-CSF (pg/mL) | 340.63 ± 15.24 b | 318.09 ± 12.77 b | 81.68 ± 1.26 a |
| Interleukin 4 (ng/L) | 965.24 ± 17.29 c | 686.64 ± 6.41 b | 349.85 ± 3.05 a |
| Interferon-γ (pg/mL) | 352.90 ± 14.50 b | 256.05 ± 12.94 ab | 167.42 ± 4.38 a |
| Oxidant/antioxidant markers | |||
| Malondialdehyde (nM/mL) | 5.10 ± 0.05 b | 4.00 ± 1.51 b | 1.37 ± 0.02 a |
| Superoxide dismutase (U/mL) | 969.90 ± 5.69 c | 937.66 ± 13.03 b | 546.6 ± 4.64 a |
| Glutathione peroxidase (U/mL) | 242.53 ± 12.82 b | 235.91 ± 6.34 b | 54.40 ± 0.45 a |
| Proinflammatory/ Anti-Inflammatory Cytokines | T. evansi Infected Cattle (Symptomatic) (n = 4) | Clinically Healthy Cattle (n = 8) |
|---|---|---|
| Interleukin 12 (ng/L) * | 8.96 ± 0.53 | 2.36 ± 0.08 |
| Interleukin 10 (ng/L) * | 71.00 ± 1.83 | 19.53 ± 0.43 |
| GM-CSF (pg/mL) * | 555.42 ± 20.7 | 81.68 ± 1.26 |
| Interleukin 4 (ng/L) * | 770.41 ± 14.82 | 349.85 ± 3.05 |
| Interferon-γ (pg/mL) * | 277.02 ± 5.41 | 167.42 ± 4.38 |
| Oxidant/Antioxidant Markers | ||
| Malondialdehyde (nM/mL) * | 13.73 ± 0.15 | 1.37 ± 0.02 |
| Superoxide dismutase (U/mL) * | 1162.76 ± 19.01 | 546.6 ± 4.64 |
| Glutathione peroxidase (U/mL) * | 128.51 ± 2.10 | 54.40 ± 0.45 |
| Proinflammatory/ Anti-Inflammatory Cytokines | Anaplasma platys Infected Cattle (Symptomatic) (n = 3) | Clinically Healthy Cattle (n = 8) |
|---|---|---|
| Interleukin 12 (ng/L) * | 58.60 ± 0.07 | 2.36 ± 0.08 |
| Interleukin 10 (ng/L) * | 665.04 ± 5.05 | 19.53 ± 0.43 |
| GM-CSF (pg/mL) * | 422.28 ± 10.93 | 81.68 ± 1.26 |
| Interleukin 4 (ng/L) * | 1279.33 ± 20.12 | 349.85 ± 3.05 |
| Interferon-γ (pg/mL) * | 350.12 ± 4.26 | 167.42 ± 4.38 |
| Oxidant/Antioxidant Markers | ||
| Malondialdehyde (nM/mL) * | 5.84 ± 0.15 | 1.37 ± 0.02 |
| Superoxide dismutase (U/mL) * | 1561.46 ± 15.74 | 546.6 ± 4.64 |
| Glutathione peroxidase (U/mL) * | 343.14 ± 14.09 | 54.40 ± 0.45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agina, O.A.; Cheah, K.T.; Sayuti, N.S.A.; Shaari, M.R.; Isa, N.M.M.; Ajat, M.; Zamri-Saad, M.; Mazlan, M.; Hamzah, H. High Granulocyte-Macrophage Colony Stimulating Factor to Interleukin 10 Ratio and Marked Antioxidant Enzyme Activities Predominate in Symptomatic Cattle Naturally Infected with Candidatus Mycoplasma haemobos, Theileria orientalis, Theileria sinensis and Trypanosoma evansi. Animals 2021, 11, 2235. https://doi.org/10.3390/ani11082235
Agina OA, Cheah KT, Sayuti NSA, Shaari MR, Isa NMM, Ajat M, Zamri-Saad M, Mazlan M, Hamzah H. High Granulocyte-Macrophage Colony Stimulating Factor to Interleukin 10 Ratio and Marked Antioxidant Enzyme Activities Predominate in Symptomatic Cattle Naturally Infected with Candidatus Mycoplasma haemobos, Theileria orientalis, Theileria sinensis and Trypanosoma evansi. Animals. 2021; 11(8):2235. https://doi.org/10.3390/ani11082235
Chicago/Turabian StyleAgina, Onyinyechukwu Ada, Kim Tho Cheah, Nurul Syahirah Ahmad Sayuti, Mohd Rosly Shaari, Nur Mahiza Md Isa, Mokrish Ajat, Mohd Zamri-Saad, Mazlina Mazlan, and Hazilawati Hamzah. 2021. "High Granulocyte-Macrophage Colony Stimulating Factor to Interleukin 10 Ratio and Marked Antioxidant Enzyme Activities Predominate in Symptomatic Cattle Naturally Infected with Candidatus Mycoplasma haemobos, Theileria orientalis, Theileria sinensis and Trypanosoma evansi" Animals 11, no. 8: 2235. https://doi.org/10.3390/ani11082235
APA StyleAgina, O. A., Cheah, K. T., Sayuti, N. S. A., Shaari, M. R., Isa, N. M. M., Ajat, M., Zamri-Saad, M., Mazlan, M., & Hamzah, H. (2021). High Granulocyte-Macrophage Colony Stimulating Factor to Interleukin 10 Ratio and Marked Antioxidant Enzyme Activities Predominate in Symptomatic Cattle Naturally Infected with Candidatus Mycoplasma haemobos, Theileria orientalis, Theileria sinensis and Trypanosoma evansi. Animals, 11(8), 2235. https://doi.org/10.3390/ani11082235