Bias of Calf Sex on Milk Yield and Fat Yield in Holstein Crossbreed Cows

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Statistical Analyses and Models
- Yijklmsn—phenotypic expression of the investigated trait;
- µ—general average of the population;
- Fi—fixed effect of i-th farm (i = 1–7);
- Gj—fixed effect of j-th year of calving (j = 1985–2017);
- Sk—fixed effect of k-th season of calving (k = 1–4; seasons: winter (December, January and February), spring (December, January and February), summer (June, July and August), and autumn (September, October and November));
- Ll—fixed effect of l-th lactation (l = 1–2);
- Hm—fixed effect of the group according to the share of genes of the Holstein Friesian breed, 1st group—animals with 0 to 50% share of genes of the Holstein Friesian breed; 2nd group—animals with 51 to 75% share of genes of the Holstein Friesian breed; 3rd group—animals with 76 to 87.5% share of genes of the Holstein Friesian breed; 4th group—animals with 88 to 93.75 share of genes of the Holstein Friesian breed; 5th group—animals with more than 93.5% share of genes of Holstein Friesian breed);
- Ks—fixed effect of the level of milk yield on farms (above and below average farms; (s = 1–2);
- Rn—fixed effect of sex of the calf (n = H,B; heifer-H; bull-B);
- (RxL)nl—interaction between sex and lactation order; (nl = 1–4);
- eijklmsn—random error.
- Rn—effect of calf sex class (HH, HB, BH, BB);
- (R × K)ns—interaction between calf sex class and level of milk yield on farms (above and below average farms);
- eijklmsn—random error of observation.
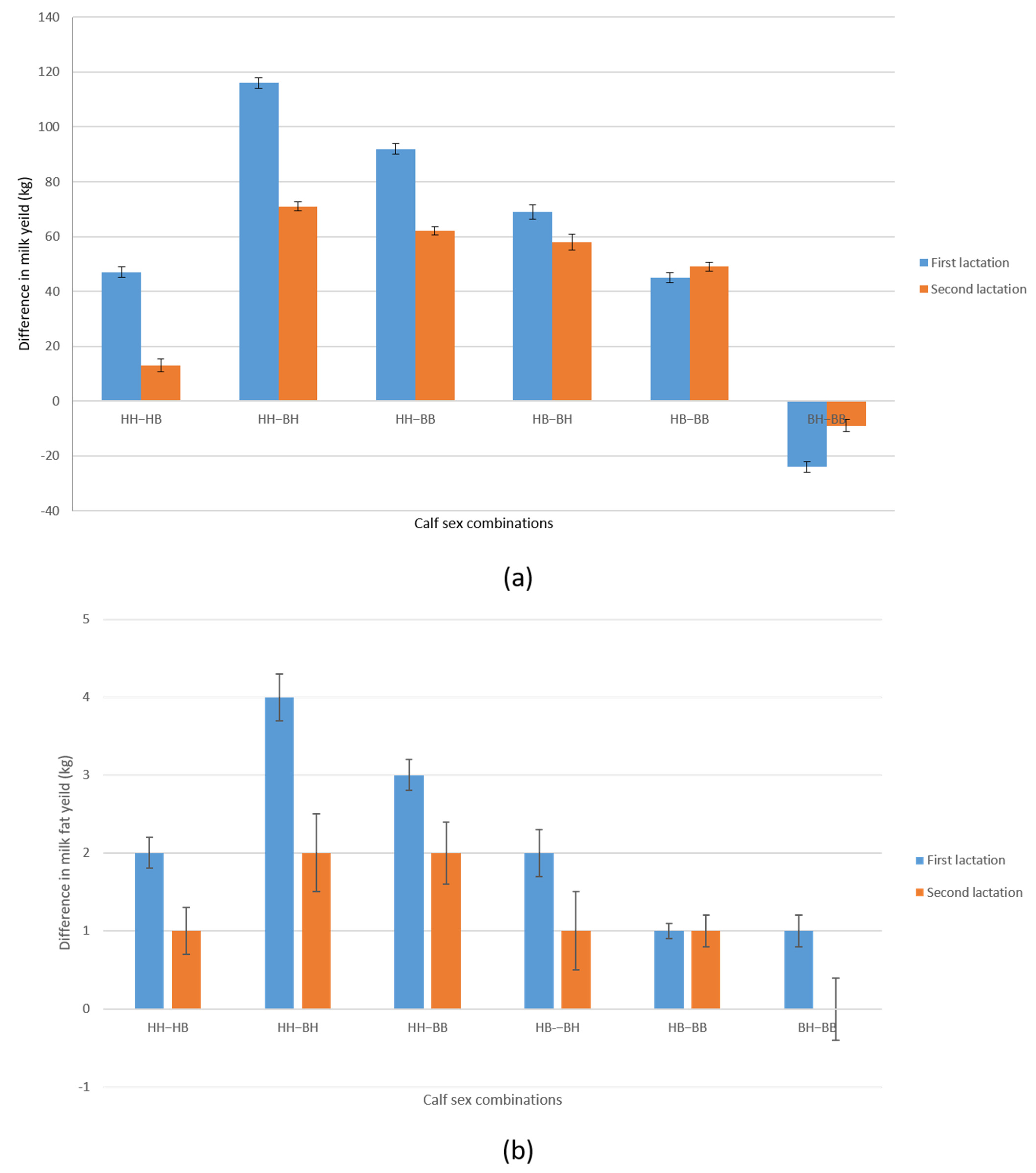
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hinde, K.; Carpenter, A.J.; Clay, J.S.; Bradford, B. Holsteins Favor Heifers, Not Bulls: Biased Milk Production Programmed during Pregnancy as a Function of Fetal Sex. PLoS ONE 2014, 9, e86169. [Google Scholar] [CrossRef] [Green Version]
- Quaresma, M.; Payan-Carreira, R. Calf-Sex Influence in Bovine Milk Production. In Animal Reproduction in Veterinary Medicine; Aral, F., Payan-Carreira, R., Quaresma, M., Eds.; IntechOpen: London, UK, 2021. [Google Scholar]
- Trivers, R.L.; Willard, D.E. Natural Selection of Parental Ability to Vary the Sex Ratio of Offspring. Science 1973, 179, 90–92. [Google Scholar] [CrossRef] [Green Version]
- Hewison, A.J.M.; Gaillard, J.M. Successful sons or advantaged daughters? The Trivers-Willard model and sex-biased maternal investment in ungulates. Trends Ecol. Evol. 1999, 14, 229–234. [Google Scholar] [CrossRef]
- Cameron, E.Z. Facultative adjustment of mammalian sex ratios in support of the Trivers-Willard hypothesis: Evidence for a mechanism. Proc. R. Soc. B Biol. Sci. 2004, 271, 1723–1728. [Google Scholar] [CrossRef] [Green Version]
- Fujita, M.; Roth, E.; Lo, Y.-J.; Hurst, C.; Vollner, J.; Kendell, A. In poor families, mothers’ milk is richer for daughters than sons: A test of Trivers-Willard hypothesis in agropastoral settlements in Northern Kenya. Am. J. Phys. Anthr. 2012, 149, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Douhard, M. Offspring sex ratio in mammals and the Trivers-Willard hypothesis: In pursuit of unambiguous evidence. BioEssays 2017, 39, 1700043. [Google Scholar] [CrossRef] [PubMed]
- Powe, C.E.; Knott, C.D.; Conklin-Brittain, N. Infant sex predicts breast milk energy content. Am. J. Hum. Biol. 2010, 22, 50–54. [Google Scholar] [CrossRef]
- Beavers, L.; Van Doormaal, B. Is Sex-Biased Milk Production a Real Thing? Canadian Dairy Network. 2014. Available online: https://www.google.at/url?sa=t&rct=j&q=&esrc=s&source=web&cd=1&cad=rja&uact=8&ved=0CCAQFjAA&url=https://www.cdn.ca/document.php?id=348&ei=L6lZVcu9HMHoUMuZgYgI&usg=AFQjCNFksVPmiJ3bmEew-wqkC2n3E_Ilvg&sig2=qQVunmVgV5ROSBwxYYU57A&bvm=bv.93564037,d.d24 (accessed on 26 April 2021).
- Chegini, A.; Hossein-Zadeh, N.G.; Hosseini-Moghadam, H. Effect of calf sex on some productive, reproductive and health traits in Holstein cows. Span. J. Agric. Res. 2015, 13, e0605. [Google Scholar] [CrossRef] [Green Version]
- Hess, M.; Hess, A.; Garrick, D. The Effect of Calf Gender on Milk Production in Seasonal Calving Cows and Its Impact on Genetic Evaluations. PLoS ONE 2016, 11, e0151236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yudin, N.S.; Aitnazarov, R.B.; Voevoda, M.I.; Gerlinskaya, L.A.; Moshkin, M.P. Association of Polymorphism Harbored by Tumor Necrosis Factor Alpha Gene and Sex of Calf with Lactation Performance in Cattle. Asian-Australas. J. Anim. Sci. 2013, 26, 1379–1387. [Google Scholar] [CrossRef] [PubMed]
- Barbat, A.; Lefebvre, R.; Boichard, D. Replication study in French Holstein and Montbeliarde Cattle Data. 2014. Available online: http://www.plosone.org/annotation/listThread.action?root=78955 (accessed on 4 May 2021).
- Græsbøll, K.; Kirkeby, C.; Nielsen, S.S.; Christiansen, L.E. Danish Holsteins Favor Bull Offspring: Biased Milk Production as a Function of Fetal Sex, and Calving Difficulty. PLoS ONE 2015, 10, e0124051. [Google Scholar] [CrossRef] [Green Version]
- Djedović, R.; Bogdanović, V.; Stanojević, D.; Nemes, Z.; Gáspárdy, A.; Cseh, S. Involuntary reduction in vigour of calves born from sexed semen. Acta Vet. Hung. 2016, 64, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Djedović, R.; Bogdanović, V.; Stanojević, D.; Samolovac, L.; Brka, M. Phenotypic variability of fertility and milk traits in offspring obtained by insemination by sexed and conventional semen of Holstein breed bulls. J. Agric. Food Environ. Sci. 2017, 71, 1–8. [Google Scholar]
- Clark, R.D.; Touchberry, R. Effect of Body Weight and Age at Calving on Milk Production in Holstein Cattle. J. Dairy Sci. 1962, 45, 1500–1510. [Google Scholar] [CrossRef]
- Řehák, D.; Volek, J.; Bartoň, L.; Vodková, Z.; Kubešová, M.; Rajmon, R. Relationships among milk yield, body weight, and reproduction in Holstein and Czech Fleckvieh cows. Czech J. Anim. Sci. 2012, 57, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Sellen, D.W. Evolution of Infant and Young Child Feeding: Implications for Contemporary Public Health. Annu. Rev. Nutr. 2007, 27, 123–148. [Google Scholar] [CrossRef] [Green Version]
- Hinde, K. Richer milk for sons but more milk for daughters: Sex-biased investment during lactation varies with maternal life history in rhesus macaques. Am. J. Hum. Biol. 2009, 21, 512–519. [Google Scholar] [CrossRef]
- Hinde, K.; Milligan, L.A. Primate milk: Proximate mechanisms and ultimate perspectives. Evol. Anthropol. Issues News Rev. 2011, 20, 9–23. [Google Scholar] [CrossRef]
- Forsyth, A. Book Reviews and Notes: A Natural History of Sex. Bull. Sci. Technol. Soc. 1988, 8, 93. [Google Scholar]
- Gorewit, R.C.; Svennersten, K.; Butler, W.R.; Uvnäs-Moberg, K. Endocrine Responses in Cows Milked by Hand and Machine. J. Dairy Sci. 1992, 75, 443–448. [Google Scholar] [CrossRef]
- Knight, C.H. Lactation and gestation in dairy cows: Flexibility avoids nutritional extremes. Proc. Nutr. Soc. 2001, 60, 527–537. [Google Scholar] [CrossRef] [Green Version]
- Hennighausen, L.; Robinson, G.W. Information networks in the mammary gland. Nat. Rev. Mol. Cell Biol. 2005, 6, 715–725. [Google Scholar] [CrossRef]
- Gillespie, A.V.; Ehrlich, J.L.; Grove-White, D.H. Effect of Calf Gender on Milk Yield and Fatty Acid Content in Holstein Dairy Cows. PLoS ONE 2017, 12, e0169503. [Google Scholar] [CrossRef] [PubMed]
- Soyeurt, H.; Dehareng, F.; Gengler, N.; McParland, S.; Wall, E.; Berry, D.P.; Coffey, M.; Dardenne, P. Mid-infrared prediction of bovine milk fatty acids across multiple breeds, production systems, and countries. J. Dairy Sci. 2011, 94, 1657–1667. [Google Scholar] [CrossRef] [Green Version]
- Woolpert, M.; Dann, H.; Cotanch, K.; Melilli, C.; Chase, L.; Grant, R.; Barbano, D. Management, nutrition, and lactation performance are related to bulk tank milk de novo fatty acid concentration on northeastern US dairy farms. J. Dairy Sci. 2016, 99, 8486–8497. [Google Scholar] [CrossRef] [PubMed]
- Stoop, W.; Bovenhuis, H.; Heck, J.; van Arendonk, J. Effect of lactation stage and energy status on milk fat composition of Holstein-Friesian cows. J. Dairy Sci. 2009, 92, 1469–1478. [Google Scholar] [CrossRef]
- Sawa, A.; Jankowska, M.; Glowska, M. Effect of some factors on sex of the calf born, and of sex of the calf on performance of dairy cows. Acta Sci. Pol. Zootech. 2014, 13, 75–83. [Google Scholar]
- Hayr, M.K.; Hess, A.S.; Garrick, D.J. Impact of Including Calf Gender in Models to Predict Breeding Values for Lactation Yields in Dairy Cattle. Iowa State Univ. Anim. Ind. Rep. 2015, 12. [Google Scholar] [CrossRef]
- Swali, A.; Wathes, D. Influence of the dam and sire on size at birth and subsequent growth, milk production and fertility in dairy heifers. Theriogenology 2006, 66, 1173–1184. [Google Scholar] [CrossRef] [PubMed]
- Mee, J. Prevalence and risk factors for dystocia in dairy cattle: A review. Vet. J. 2008, 176, 93–101. [Google Scholar] [CrossRef]
- Zaborski, D.; Grzesiak, W.; Szatkowska, I.; Dybus, A.; Muszynska, M.; Jedrzejczak, M. Factors Affecting Dystocia in Cattle. Reprod. Domest. Anim. 2009, 44, 540–551. [Google Scholar] [CrossRef]
- Atashi, H.; Abdolmohammadi, A.; Dadpasand, M.; Asaadi, A. Prevalence, Risk Factors and Consequent Effect of Dystocia in Holstein Dairy Cows in Iran. Asian-Australas. J. Anim. Sci. 2012, 25, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Anwar, M.; Mirza, M.A. Some Factors Affecting Milk Yield and Lactation. Pak. Vet. J. 2007, 27, 113–117. [Google Scholar]
- Booksmythe, I.; Mautz, B.; Davis, J.; Nakagawa, S.; Jennions, M.D. Facultative adjustment of the offspring sex ratio and male attractiveness: A systematic review and meta-analysis. Biol. Rev. 2015, 92, 108–134. [Google Scholar] [CrossRef] [PubMed]
- Hossein-Zadeh, N.G.; Nejati-Javaremi, A.; Miraei-Ashtiani, S.R.; Kohram, H. Bio-economic evaluation of the use of sexed semen at different conception rates and herd sizes in Holstein populations. Anim. Reprod. Sci. 2010, 121, 17–23. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Categories | Criterion |
|---|---|
| Bulls for insemination | Only Holstein bulls Semen from the same bulls was used on all analyzed farms Data (dates) of all inseminations during the period from 1982 to 2017 are known |
| Cows | Holstein type cows, and the average share of Holstein Friesian genes for animals covered by the analysis was 84%. Both parents, i.e., the origins are known No sexed semen or recombinant bovine somatotropin were used for cows covered by the analysis All cows remain in the same herd-farm from the first to the last calving Data about the sex of calves are known for every calving |
| Farms | Farms were divided into two groups (above average and below average) based on two criteria Average milk yield in the first two standard lactations was 14,337, i.e., 6738 kg for the first and 7599 kg for the second lactation. Body weight for two categories (first calvers and cows) First calvers—below average farms < 520 kg First calvers—above average farms > 520 kg Cows—below average farms < 670 kg Cows—above average farms > 670 kg |
| Combinations of Calf Sexes at First and Second Calving | Farms with Below Average Milk Production in the First Two Lactations | Farms with Above Average Milk Production in the First Two Lactations |
|---|---|---|
| MY, kg ± SD | MY, kg ± SD | |
| HH | 13,027 ± 2794 (n = 1083) | 14,750 ± 3608 (n = 2735) |
| HB | 12,914 ± 2692 (n = 1169) | 14,691 ± 2993 (n = 2817) |
| BH | 13,130 ± 2805 (n = 1117) | 14,525 ± 3001 (n = 2480) |
| BB | 12,844 ± 2662 (n = 1158) | 14,587 ± 3080 (n = 2622) |
| Item | 1st Lactation | 2nd Lactation | ||
|---|---|---|---|---|
| Sex | H | B | H | B |
| (n) | (n = 7830) | (n = 7351) | (n = 7362) | (n = 7819) |
| MY, kg ( ± SD) | 6777 A ± 1564 | 6697 B ± 1563 | 7601 C ± 1902 | 7597 D ± 1888 |
| FY, kg ( ± SD) | 242 a ± 53.74 | 240 b ± 54.68 | 270 ± 65.45 | 269 ± 65.32 |
| Trait/Source | DF | MY, kg | FY, kg | ||
|---|---|---|---|---|---|
| F-Value Estimate ±SE | p-Value | F-Value Estimate ±SE | p-Value | ||
| Farm | 6 | 368 *** −69.2 ± 3.6 | <0.001 | 516 *** −4 ± 0.2 | <0.001 |
| Year of first calving | 31 | 382 *** 117.8 ± 1.9 | <0.001 | 203 *** 6 ± 0.4 | <0.001 |
| Season of calving | 3 | 0.12 ns −1.9 ± 6.6 | =0.8612 | 0.10 ns −0.09 ± 0.1 | =0.912 |
| Lactation order | 1 | 412 *** 869.4 ± 14.6 | <0.001 | 270 *** 7 ± 0.3 | <0.001 |
| Share of genes of the HF breed | 4 | 205 *** 148.2 ± 9.9 | <0.001 | 235 *** 5 ± 0.1 | <0.001 |
| Sex (H,B) | 1 | 51 ** 15.6 ± 14.5 | <0.01 | 29 ** 2 ± 0.6 | <0.01 |
| Level of milk yield on farms (above and below-average milk production) | 1 | 285 *** 248 ± 15 | <0.001 | 8.4 * 1 ± 1.2 | <0.05 |
| Combination of calf sexes at first and second calving | 3 | 33 ** 15.6 ± 14.5 | <0.01 | 0.85 ns −0.9 ± 5 | =0.771 |
| Interaction between sex and lactation order | 3 | 47 ** 16.3 ± 15.1 | <0.01 | 0.91 ns 1.1 ± 7 | =0.861 |
| Interaction between the combination of calf sexes (sex class) and level of milk yield on farms | 7 | 67 ** 13.6 ± 12.6 | <0.01 | 9.6 * 1.2 ± 1.5 | <0.05 |
| Combinations of Calf Sexes at First and Second Calving | MY, kg | FY, kg | ||
|---|---|---|---|---|
| 1st Lactation ± SD | 2nd Lactation ± SD | 1st Lactation ± SD | 2nd Lactation ± SD | |
| HH | 6801 A ± 1562 (n = 3788) | 7634 E ± 1909 (n = 3788) | 243 A,a ± 53.74 (n = 3788) | 270 B ± 65.45 (n = 3788) |
| HB | 6754 B ± 1566 (n = 3563) | 7621 F ± 1892 (n = 3563) | 241 A ± 54.20 (n = 3563) | 269 B ± 65.32 (n = 3563) |
| BH | 6685 C ± 1558 (n = 4031) | 7563 G ± 1894 (n = 4031) | 239 A,b ± 54.50 (n = 4031) | 268 B ± 65.07 (n = 4031) |
| BB | 6709 D ± 1569 (n = 3799) | 7572 H ± 1884 (n = 3799) | 240 A ± 54.68 (n = 3799) | 268 B ± 65.53 (n = 3799) |
| Combinations of Calf Sexes at First and Second Calving | Farms with Below Average Milk Production | Farms with Above Average Milk Production | Farms with Below Average Milk Production | Farms with Above Average Milk Production | ||||
|---|---|---|---|---|---|---|---|---|
| MY, kg ± SD | MY, kg ± SD | FY, kg ± SD | FY, kg ± SD | |||||
| 1st Lactation | 2nd Lactation | 1st Lactation | 2nd Lactation | 1st Lactation | 2nd Lactation | 1st Lactation | 2nd Lactation | |
| HH | 6150 A,B,¶ ± 1346 | 6877 C,§ ± 1781 | 6962 A,£ ± 1529 | 7788 C,¥ ± 1879 | 224 £ ± 48.41 | 248 A,B,C,¶ ± 64.06 | 246 ¶ ± 50.76 | 271 § ± 61.44 |
| HB | 6093 A,¶ ± 1366 | 6821 D,§ ± 1671 | 6928 B,£ ± 1511 | 7762 C,D,¥ ± 1830 | 221 £ ± 48.87 | 245 A ± 50.28 | 245 ¶ ± 54.77 | 271 § ± 61.73 |
| BH | 6161 A,B,¶ ± 1391 | 6868 E,§ ± 1692 | 6820 A,B,£ ± 1501 | 7705 D,¥ ± 1846 | 223 £ ± 49.32 | 251 B,¶ ± 62.04 | 242 D,¶ ± 50.45 | 269 § ± 60.96 |
| BB | 6061 B,¶ ± 1327 | 6783 F,§ ± 1670 | 6821 A,B,£ ± 1488 | 7774 C,D,¥ ± 1886 | 220 £ ± 46.76 | 243 C,¶ ± 58.61 | 243 D,¶ ± 53.32 | 270 § ± 62.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djedović, R.; Stanojević, D.; Bogdanović, V.; Andrić, D.O.; Samolovac, L.; Stamenić, T. Bias of Calf Sex on Milk Yield and Fat Yield in Holstein Crossbreed Cows. Animals 2021, 11, 2536. https://doi.org/10.3390/ani11092536
Djedović R, Stanojević D, Bogdanović V, Andrić DO, Samolovac L, Stamenić T. Bias of Calf Sex on Milk Yield and Fat Yield in Holstein Crossbreed Cows. Animals. 2021; 11(9):2536. https://doi.org/10.3390/ani11092536
Chicago/Turabian StyleDjedović, Radica, Dragan Stanojević, Vladan Bogdanović, Dušica Ostojić Andrić, Ljiljana Samolovac, and Tamara Stamenić. 2021. "Bias of Calf Sex on Milk Yield and Fat Yield in Holstein Crossbreed Cows" Animals 11, no. 9: 2536. https://doi.org/10.3390/ani11092536
APA StyleDjedović, R., Stanojević, D., Bogdanović, V., Andrić, D. O., Samolovac, L., & Stamenić, T. (2021). Bias of Calf Sex on Milk Yield and Fat Yield in Holstein Crossbreed Cows. Animals, 11(9), 2536. https://doi.org/10.3390/ani11092536