
Prey-Base Does Not Influence Breeding Success in Eagle Owls (Bubo bubo) in Judea, Israel





Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Study Area
2.3. Data Collection
2.4. Statistical Analysis
3. Results
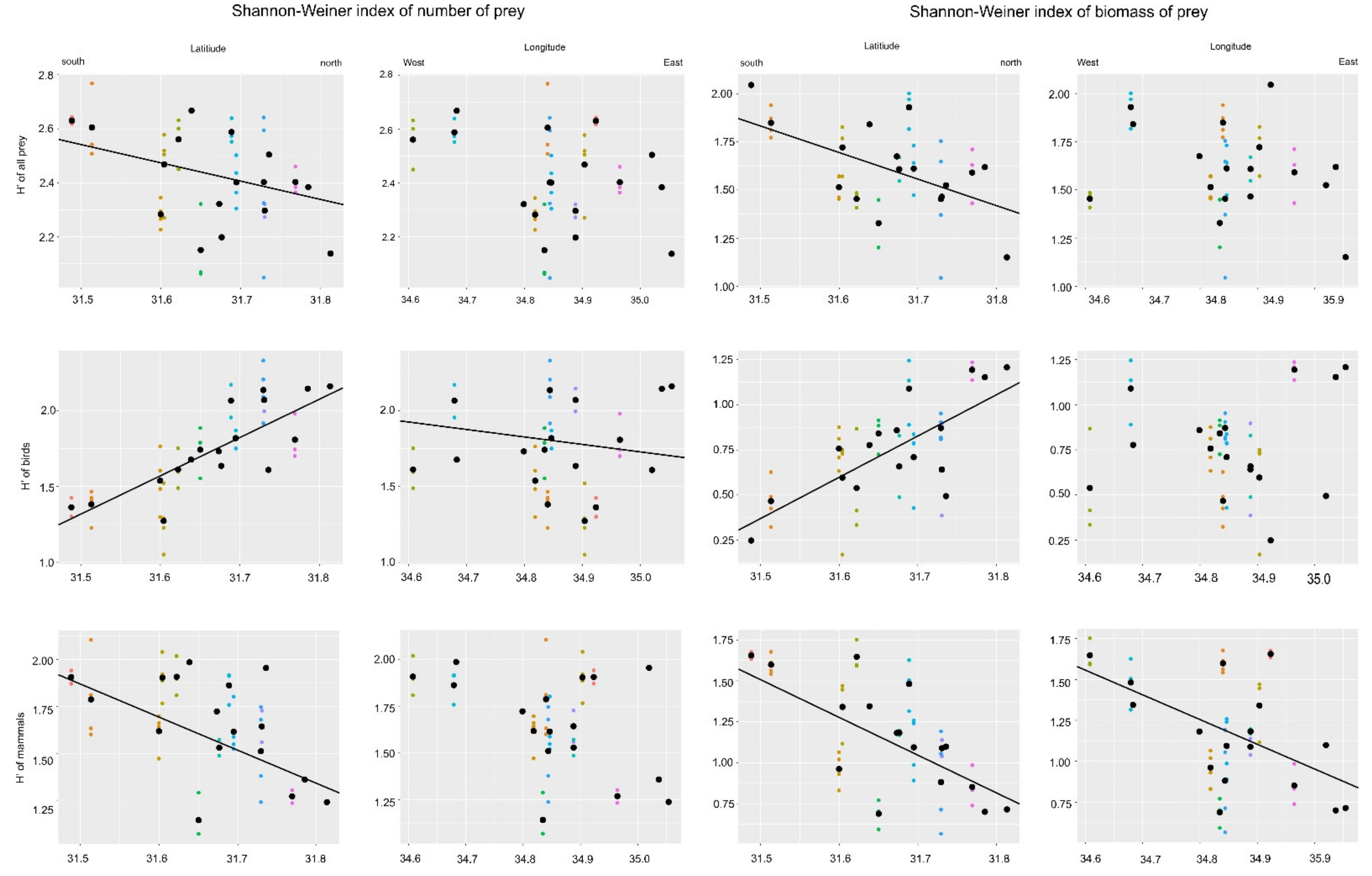
3.1. Diversity of Prey Numbers
3.2. Diversity of Prey Biomass
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newton, I. Population Ecology of Raptors; Poyser: Berkhamstead, UK, 1979. [Google Scholar]
- Newton, I. Population limitation in birds: The last 100 years. Br. Birds 2007, 100, 518–539. [Google Scholar]
- Lehikoinen, A.; Ranta, E.; Pietiäinen, H.; Byholm, P.; Saurola, P.; Valkama, J. The impact of climate and cyclic food abundance on the timing of breeding and brood size in four boreal owl species. Oecologia 2011, 165, 349–355. [Google Scholar] [CrossRef]
- Charter, M.; Izhaki, I.; Meyrom, K.; Aviel, S.; Leshem, Y.; Roulin, A. The relationship between weather and reproduction of the barn owl Tyto alba in a semi-arid agricultural landscape in Israel. Avian Biol. Res. 2017, 10, 253–258. [Google Scholar] [CrossRef]
- Klok, C.; De Roos, A.M. Effects of vole fluctuations on the population dynamics of the barn owl Tyto alba. Acta Biotheor. 2007, 55, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Amr, Z.S.; Handal, E.N.; Bibi, F.; Najajrah, M.H.; Qumsiyeh, M.B. Change in diet of the Eurasian eagle owl (Bubo bubo) suggests decline in biodiversity in Wadi Al Makhrour, Bethlehem Governorate, Palestinian Territories. Slovak Raptor J. 2016, 10, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Sergio, F.; Hiraldo, F. Intraguild predation in raptor assemblages: A review. Ibis 2008, 150, 132–145. [Google Scholar] [CrossRef] [Green Version]
- Pande, S.; Pawashe, A.; Mahajan, M.; Mahabal, A.; Joglekar, C.; Yosef, R. Breeding biology, nesting habitat, and diet of the Rock Eagle-Owl (Bubo bengalensis). J. Raptor Res. 2011, 45, 211–219. [Google Scholar] [CrossRef]
- Charter, M.; Izhaki, I.; Leshem, Y.; Roulin, A. Diet and breeding success of long-eared owls in a semi-arid environment. J. Arid Environ. 2012, 85, 142–144. [Google Scholar] [CrossRef] [Green Version]
- Charter, M.; Izhaki, I.; Leshem, Y.; Meyrom, K.; Roulin, A. Relationship between diet and reproductive success in the Israeli barn owl. J. Arid Environ. 2015, 122, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, D.P.; Reid, R.; Haworth, P.F.; Madders, M.; Marquiss, M.; Tingay, R.; Fielding, A.H. Diet specificity is not associated with increased reproductive performance of Golden Eagles Aquila chrysaetos in Western Scotland. Ibis 2009, 151, 255–264. [Google Scholar] [CrossRef]
- Charter, M.; Izhaki, I.; Meyrom, K.; Roulin, A. The relationship between intra–guild diet overlap and breeding in owls in Israel. Popul. Ecol. 2018, 60, 397–403. [Google Scholar] [CrossRef]
- Korpimäki, E.; Norrdahl, K. Numerical and functional responses of kestrels, short-eared owls, and long-eared owls to vole densities. Ecology 1991, 72, 814–826. [Google Scholar] [CrossRef]
- Salamolard, M.; Butet, A.; Leroux, A.; Bretagnolle, V. Responses of an avian predator to variations in prey density at a temperate latitude. Ecology 2000, 81, 2428–2441. [Google Scholar] [CrossRef]
- Reif, V.; Jungell, S.; Korpimäki, E.; Tornberg, R.; Mykrä, S. Numerical response of common buzzards and predation rate of main and alternative prey under fluctuating food conditions. Ann. Zool. Fenn. 2004, 41, 599–607. [Google Scholar]
- Newton, I. Population Limitation in Birds; Academic Press: London, UK, 1998. [Google Scholar]
- Shirihai, H. The Birds of Israel; Oxford University Press: Oxford, UK, 1996. [Google Scholar]
- Serrano, D. Relationship between raptors and rabbits in the diet of Eagle Owls in southwestern Europe: Competition removal or food stress? J. Raptor Res. 2000, 34, 305–310. [Google Scholar]
- Sergio, F.; Marchesi, L.; Pedrini, P.; Penteriani, V. Coexistence of a generalist owl with its intraguild predator: Distance-sensitive or habitat-mediated avoidance? Anim. Behav. 2007, 74, 1607–1616. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.E.; Martínez, J.A.; Zuberogoitia, I.; Zabala, J.; Redpath, S.M.; Calvo, J.F. The effect of intra- and interspecific interactions on the large-scale distribution of cliff-nesting raptors. Ornis Fenn. 2008, 85, 13–21. [Google Scholar]
- Lourenço, R.; Santos, S.M.; Rabaça, J.E.; Penteriani, V. Superpredation patterns in four large European raptors. Popul. Ecol. 2011, 53, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Penteriani, V.; del Mar Delgado, M. The Eagle Owl; T & AD Poyser Ltd.: London, UK, 2019. [Google Scholar]
- Brambilla, M.; Rubolini, D.; Guidali, F. Eagle Owl Bubo bubo proximity can lower productivity of cliff-nesting Peregrines Falco peregrinus. Ornis Fenn. 2006, 83, 20–26. [Google Scholar]
- Tumurbat, J.; Gombobaatar, S.; Yosef, R. Nest site and food composition of the Eagle Owl bubo bubo in Mongolia. Ardea 2009, 97, 519–523. [Google Scholar] [CrossRef]
- Korpimäki, E.; Huhtala, K.; Sulkava, S.; Korpimaki, E. Does the year-to-year variation in the diet of Eagle and Ural Owls support the alternative prey hypothesis? Oikos 1990, 58, 47–54. [Google Scholar] [CrossRef]
- Martinez, E.; Sanchez, A.; Carmona, D.; Sanchez, A.; Ortuño, A.; Martinez, R. The ecology and conservation of the Eagle Owl Bubo bubo in Murcia, south-east Spain. In The Ecology and Conservation of European Owls; Joint Nature Conservation Committee: Peterborough, UK, 1992; pp. 84–88. [Google Scholar]
- Simeonov, S.; Milchev, B.; Boev, Z. Study of the Eagle Owl (Bubo bubo (L.)) (Aves: Strigiformes) in the Strandzha Mountain (Southeast Bulgaria). II. Food spectrum and trophic specialization. Acta Zool. 1998, 50, 87–100. [Google Scholar]
- Penteriani, V.; Sergio, F.; del Mar Delgado, M.; Gallardo, M.; Ferrer, M. Biases in population diet studies due to sampling in heterogeneous environments: A case study with the Eagle Owl. J. Field Ornithol. 2005, 76, 237–244. [Google Scholar] [CrossRef]
- Lourenco, R. The food habits of Eurasian eagle-owls in southern Portugal. J. Raptor Res. 2006, 40, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Sándor, A.D.; Ionescu, D.T. Diet of the eagle owl (Bubo bubo) in Braşov, Romania. North West. J. Zool. 2009, 5, 170–178. [Google Scholar]
- Obuch, J.; Karaska, D. The Eurasian eagle-owl (Bubo bubo) diet in the Orava Region (N Slovakia). Slovak Raptor J. 2010, 4, 83–98. [Google Scholar] [CrossRef]
- Hadad, E.; Shochat, E. First evidence of hunting of a Crested Indian Porcupine by an avian predator: The Eurasian Eagle Owl. Isr. J. Ecol. Evol. 2021, 1, 1–3. [Google Scholar] [CrossRef]
- Redpath, S.M.; Clarke, R.; Madders, M.; Thirgood, S.J. Assessing raptor diet: Comparing pellets, prey remains, and observational data at hen harrier nests. Condor 2001, 103, 184–188. [Google Scholar] [CrossRef]
- Yom-Tov, Y.; Wool, D. Do the contents of Barn Owl pellets accurately represent the proportion of prey species in the field. Condor 1997, 99, 972–976. [Google Scholar] [CrossRef] [Green Version]
- Penteriani, V.; del Mar Delgado, M.; Gallardo, M.; Ferrer, M. Spatial heterogeneity and structure of bird populations: A case example with the eagle owl. Popul. Ecol. 2004, 46, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Dalbeck, L.; Heg, D. Reproductive success of a reintroduced population of Eagle Owls Bubo bubo in relation to habitat characteristics in the Eifel, Germany. Ardea 2006, 94, 3–21. [Google Scholar]
- León-Ortega, M.; Martínez, J.E.; Pérez, E.; Lacalle, J.A.; Calvo, J.F. The contribution of non-protected areas to the conservation of Eurasian Eagle-owls in Mediterranean ecosystems. Ecosphere 2017, 8, e01952. [Google Scholar] [CrossRef]
- Andreychev, A.V.; Lapshin, A.S.; Kuznetsov, V.A. Breeding success of the Eurasian eagle owl (Bubo bubo) and rodent population dynamics. Biol. Bull. 2016, 43, 851–861. [Google Scholar] [CrossRef]
- Bionda, R.; Brambilla, M. Rainfall and landscape features affect productivity in an alpine population of Eagle Owl Bubo bubo. J. Ornithol. 2012, 153, 167–171. [Google Scholar] [CrossRef]
- Shehab, A.H. Diet of the Eagle Owl, Bubo bubo, in Syria. Zool. Middle East 2004, 33, 21–26. [Google Scholar] [CrossRef]
- Obuch, J. On the diet of owls (Strigiformes) in Jordan. Slovak Raptor J. 2018, 12, 9–40. [Google Scholar] [CrossRef]
- Donázar, J.A. Geographic variation in clutch and brood size of the Eagle Owl Bubo bubo in the Western Palearctic. J. Ornithol. 1990, 131, 439–443. [Google Scholar] [CrossRef]
- Korpimaki, E.; Marti, C.D. Geographical trends in trophic characteristics of mammal-eating and bird-eating raptors in Europe and North America. Auk 1995, 112, 1004–1023. [Google Scholar] [CrossRef]
- Olsson, V. Breeding success, dispersal, and long-term changes in a population of Eagle Owls Bubo bubo in southeastern Sweden 1952–1996. Ornis Svec. 1997, 7, 49–60. [Google Scholar] [CrossRef]
- Martínez, J.A.; López, G. Breeding ecology of the Barn Owl (Tyto alba) in Valencia (SE Spain). J. Ornithol. 1999, 99, 93–99. [Google Scholar] [CrossRef]
- Penteriani, V.; Gallardo, M.; Roche, P. Landscape structure and food supply affect eagle owl (Bubo bubo) density and breeding performance: A case of intra-population heterogeneity. J. Zool. 2002, 257, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Marchesi, L.; Pedrini, P.; Sergio, F. Biases associated with diet study methods in the Eurasian Eagle-Owl. J. Raptor Res. 2002, 36, 11–16. [Google Scholar]
- Mendelssohn, H.; Yom-Tov, Y. Appendix to Volume 7: Mammals. Skull and body measurements. In Plants and Animals of the Land of Israel; Alon, A., Ed.; Ministry of Defence Publishing House: Tel Aviv, Israel, 1987. [Google Scholar]
- Mendelssohn, H.; Yom-Tov, Y. Fauna Palaestina. In Mammalia of Israel; The Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1999. [Google Scholar]
- Snow, D.W.; Perrins, C.M. The Birds of the Western Palearctic; Concise Edition; Vol. 1 Non-Passerines, Vol. 2. Passerines; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Charter, M.; Izhaki, I.; Meyrom, K.; Motro, Y.; Leshem, Y. Diets of barn owls differ in the same agricultural region. Wilson J. Ornithol. 2009, 121, 378–383. [Google Scholar] [CrossRef]
- Wilson, D.E.; Mittermeier, R.A.; Lacher, T.E. Handbook of the Mammals of the World; Vol. 7. Rodents II; Lynx Ediciones: Barcelona, Spain, 2017. [Google Scholar]
- Krystufek, B.; Ivanitskaya, E.; Arslan, A.; Arslan, E.; Buzan, E. Evolutionary history of mole rats (genus Nannospalax) inferred from mitochondrial cytochrome b sequence. Biol. J. Linn. Soc. 2011, 105, 446–455. [Google Scholar] [CrossRef] [Green Version]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Delgado, M.M.; Caferri, E.; Mendez, M.; Godoy, J.A.; Campioni, L.; Penteriani, V. Population characteristics may reduce the levels of individual call identity. PLoS ONE 2013, 8, e77557. [Google Scholar] [CrossRef] [Green Version]
- Cochet, G. Quelques aspects de la biologie du Hibou grand-duc dans le Massif Central, les Causses et les Cevennes. In Actes du Colloque Interregional de St. Etienne, France; Delachaux et Niestlé: Paris, France, 1989. [Google Scholar]
- Cochet, G. Les sites a Hibou grand-duc, Bubo bubo, et la geomorphologie. In Rapaces Nocturnes: Actes du 30 Colloque Interregional s’ornithologie, Porrentruy (Suise); Julian, D., Bassin, P., Baudvin, H., Genot, J.-C., Ravussin, P.-A., Reletez, C., Eds.; Nos Oiseaux: La Chaux-de-Fonds, Switzerland, 1999; pp. 87–91. [Google Scholar]
- Wilamowski, A.; Moran, S.; Greenburg, Z. Commensal rodents and their parasites in Israel. In Proceedings of the 4th International Conference on Urban Pests, Charleston, SC, USA, 7–10 July 2002; pp. 103–113. [Google Scholar]
- Hadad, E. The bioinvasion of the Brown Rat in Israel. Teva HaDvarim 2021, 306, 32–43. (In Hebrew) [Google Scholar]
- Marchesi, L.; Sergio, F.; Pedrini, P. Costs and benefits of breeding in human-altered landscapes for the Eagle Owl Bubo bubo. Ibis 2002, 144, E164–E177. [Google Scholar] [CrossRef]
- Nevo, A. Mole rat Spalax ehrenbergi: Mating behavior and its evolutionary significance. Science 1969, 163, 3866. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species; Version 2021-3; IUCN: Gland, Switzerland, 2022. [Google Scholar]
- Hadad, E. The Beech Marten in Israel. Teva HaDvarim 2015, 240, 22–34. (In Hebrew) [Google Scholar]
- Hadad, E. The Marbled Polecat in Israel. Teva HaDvarim 2015, 242, 27–36. (In Hebrew) [Google Scholar]
- Sieradzki, A.; Mikkola, H. A review of European owls as predators of bats. In Owls; Mikkola, H., Ed.; IntechOpen: London, UK, 2020; 20p. [Google Scholar] [CrossRef] [Green Version]
- Rossina, V.V.; Shokhrin, V.P. Bats in the diet of owls from Russian Far East, southern Sikhote Alin. Hystrix 2011, 22, 205–213. [Google Scholar] [CrossRef]
- Nemtzov, S.C. Management of wildlife-human conflicts in Israel: A wide variety of vertebrate pest problems in a difficult and compact environment. In Proceedings of the 20th Vertebrate Pest Conference; Timm, R.M., Schmidt, R.H., Eds.; University of California: Oakland, CA, USA, 2002; pp. 348–353. [Google Scholar]
- Hadad, E. The terrible injustice caused to bats by humans. Teva HaDvarim 2018, 271, 16–28. (In Hebrew) [Google Scholar]
- Dolev, A.; Pervolutzky, A. (Eds.) Endangered Species in ISRAEL—Red List of Threatened Species; Nature and Parks Authority and Society for Protection of Nature: Jerusalem, Israel, 2002. [Google Scholar]
- Papageorgiou, N.K.; Vlachos, C.G.; Bakaloudis, D.E. Diet and nest site characteristics of Eagle Owl (Bubo bubo) breeding in two different habitats in north-eastern Greece. Avocetta 1993, 17, 49–54. [Google Scholar]
- Dalbeck, L. Der Uhu Bubo bubo (L.) in Deutschland—Autokologische Analysen an Einer Wieder Angesiedelten Population—Resumee Eines Artenschutzprojekts; D98 (PhD Diss. Univ. Bonn); Shaker Verlag: Herzogenrath, Germany, 2003; p. 156. [Google Scholar]
- Milchev, B. Dietary comparison of coexisting barn owl (Tyto alba) and eagle owl (Bubo bubo) during consecutive breeding seasons. Anim. Biol. 2016, 66, 219–228. [Google Scholar] [CrossRef]
- Donázar, J.A.; Hiraldo, F.; Delibes, M.; Estrella, R.R. Comparative food habits of the Eagle Owl Bubo bubo and the Great Horned Owl Bubo virginianus in six Palearctic and Nearctic biomes. Ornis Scand. 1989, 20, 298–306. [Google Scholar] [CrossRef]
- Martinez, J.E.; Calvo, J.E. Diet and breeding success of Eagle Owl in Southeastern Spain: Effect of rabbit haemorrhagic disease. J. Raptor Res. 2001, 35, 259–262. [Google Scholar]
- Fernandez-de-Simon, J.; Diaz-Ruiz, F.; Cirilli, F.; Tortosa, F.S.; Villafuerte, R.; Ferreras, P. Role of prey and intraspecific density dependence on the population growth of an avian top predator. Acta Oecol. 2014, 60, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Tobajas, J.; Rouco, C.; Fernandez de-Simon, F.; Diaz-Ruiz, F.; Castro, F.; Villafuerte, R.; Ferreras, P. Does prey abundance affect prey size selection by the Eagle Owl (Bubo bubo). J. Ornithol. 2021, 162, 699–708. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Number of Nestlings (±SE) | H’all (±SE) | H’mammals (±SE) | H’birds (±SE) | Biomass H’all (±SE) | Biomass H’mammals (±SE) | Biomass H’birds (±SE) |
|---|---|---|---|---|---|---|---|
| 2006 | 2.68 ± 0.28 | 2.39 ± 0.05 | 1.63 ± 0.08 | 1.76 ± 0.09 | 1.59 ± 0.06 | 1.14 ± 0.10 | 0.73 ± 0.08 |
| 2007 | 2.71 ± 0.26 | 2.40 ± 0.08 | 1.63 ± 0.08 | 1.73 ± 0.09 | 1.59 ± 0.06 | 1.15 ± 0.09 | 0.74 ± 0.08 |
| 2008 | 2.72 ± 0.27 | 2.42 ± 0.05 | 1.66 ± 0.08 | 1.69 ± 0.09 | 1.63 ± 0.06 | 1.17 ± 0.10 | 0.75 ± 0.08 |
| 2009 | 2.71 ± 0.26 | 2.41 ± 0.05 | 1.67 ± 0.08 | 1.68 ± 0.09 | 1.62 ± 0.06 | 1.16 ± 0.10 | 0.75 ± 0.08 |
| all | 2.67 ± 0.13 | 2.41 ± 0.02 | 1.64 ± 0.04 | 1.71 ± 0.04 | 1.61 ± 0.03 | 1.17 ± 0.05 | 0.78 ± 0.04 |
| Prey Specimens | Number | % of Total Prey | Body Mass (g, Range) | Total Biomass (g) | % Biomass | |||
|---|---|---|---|---|---|---|---|---|
| Vertebrata | (Average) | |||||||
| Mammals | Insectivora | |||||||
| Erinaceidae | Erinaceus concolor | 720 | 7.61% | 350–880 (492.0) | 354,240.0 | 4.2 | ||
| Hemiechinus auritus | 14 | 0.15% | 131–358 (244.5) | 3423.0 | 0.04 | |||
| Paraechinus aethiopicus | 1 | 0.01% | 227–605 (416.0) | 416.0 | 0.004 | |||
| Soricidae | Crocidura spp. | 1 | 0.01% | 1.8–3.6 (2.7) | 2.7 | 0.001 | ||
| Crocidura suaveolens | 60 | 0.63% | 2.4–8.5 (5.5) | 330.0 | 0.01 | |||
| Crocidura leucodon | 14 | 0.15% | 7–15 (10.5) | 147.0 | 0.008 | |||
| Chiroptera | ||||||||
| Pteropodidae | Rousettus aegyptiacus | 1103 | 11.66% | 90–210 (150.0) | 165,450.0 | 2 | ||
| Rhinopomatidae | Rhinopoma microphyllum | 2 | 0.02% | 16–24 (20.0) | 40.0 | 0.002 | ||
| Emballonuridae | Taphozous nudiventris | 3 | 0.03% | 30–60 (45.0) | 135.0 | 0.007 | ||
| Taphozous perforatus | 2 | 0.02% | 22–33 (27.5) | 55.0 | 0.002 | |||
| Nycteridae | Nycteris thebaica | 1 | 0.01% | 8–14 (11.0) | 11.0 | 0.0005 | ||
| Rhinolophidae | Rhinolophus ferrumequinum | 35 | 0.37% | 10–20 (15.0) | 525.0 | 0.03 | ||
| Vespertilionidae | Eptesicus serotinus | 1 | 0.01% | 20–33 (26.5) | 26.5 | 0.01 | ||
| Hipposideridae | Otonycteris hemprichii | 5 | 0.05% | 16.5–23 (20.0) | 100.0 | 0.3 | ||
| Molossidae | Tadarida teniotis | 9 | 0.10% | 14–38 (26.0) | 234.0 | 0.01 | ||
| Lagomorpha | ||||||||
| Leporidae | Lepus capensis | 87 | 0.92% | 1250–2300 (1775.0) | 154,425.0 | 1.8 | ||
| Rodentia | ||||||||
| Cricetidae | Cricetulus migratorius | 1 | 0.01% | 18–35 (26.5) | 26.5 | 0.001 | ||
| Microtus guentheri | 2248 | 23.76% | 29–69 (49.0) | 110,152.0 | 5.6 | |||
| Muridae | Meriones tristrami | 998 | 10.55% | 48–104 (76.0) | 75,848.0 | 3.9 | ||
| Apodemus mystacinus | 26 | 0.27% | 20–57 (38.5) | 1001.0 | 0.05 | |||
| Rattus rattus | 562 | 5.94% | 100–200 (150.0) | 84,300.0 | 1.2 | |||
| Rattus norvegicus | 295 | 3.12% | 200–600 (400.0) | 118,000.0 | 1.4 | |||
| Mus musculus | 58 | 0.61% | 8–15 (11.5) | 667.0 | 0.03 | |||
| Gerbillus dasyurus | 18 | 0.19% | 15–34 (24.5) | 441.0 | 0.02 | |||
| Acomys dimidiatus | 30 | 0.32% | 26–57 (41.5) | 1245.0 | 0.06 | |||
| Spalacidae | Nannospalax ehrenbergi | 572 | 6.04% | 118–240 (179.0) | 102,388.0 | 5.2 | ||
| Dipodidae | Jaculus jaculus | 1 | 0.01% | 33–91 (63.5) | 63.5 | 0.003 | ||
| Hystricidae | Hystrix indica | 2 | 0.02% | 2000 | 4000.0 | 0.05 | ||
| Echimyidae | Myocastor coypus | 3 | 0.03% | 1000–2000 (1500.0) | 4500.0 | 0.05 | ||
| Carnivora | ||||||||
| Mustelidae | Meles meles | 1 | 0.01% | 2000–3000 (2500.0) | 2500.0 | 0.03 | ||
| Vormela peregusna | 6 | 0.06% | 200–500 (350.0) | 2100.0 | 0.03 | |||
| Martes foina | 2 | 0.02% | 700–1300 (1000.0) | 2000.0 | 0.03 | |||
| Canidae | Vulpes vulpes | 7 | 0.07% | 2200 | 15,400.0 | 0.2 | ||
| Herpestidae | Herpestes ichneumon | 1 | 0.01% | 2000 | 2000.0 | 0.03 | ||
| Felidae | Felis catus | 7 | 0.07% | 1500 | 10,500.0 | 0.1 | ||
| Birds | ||||||||
| Pelecaniformes | ||||||||
| Ardeidae | Nycticorax nycticorax | 1 | 0.01% | 500–800 (650.0) | 650.0 | 0.03 | ||
| Bubulcus ibis | 3 | 0.03% | 300–400 (350.0) | 1050.0 | 0.05 | |||
| Threskiornithidae | Plegadis falcinellus | 1 | 0.01% | 530–760 (645.0) | 645.0 | 0.03 | ||
| Anaseriformes | ||||||||
| Anatidae | Anas platyrhynchos | 6 | 0.06% | 750–1450 (1100.0) | 6600.0 | 0.3 | ||
| Spatula clypeata | 1 | 0.01% | 470–750 (310.0) | 610.0 | 0.03 | |||
| Anas crecca\Spatula querquedula | 7 | 0.07% | 200–450 (325.0) | 2275.0 | 0.1 | |||
| Anatidae sp. | 2 | 0.02% | 200–450 (325.0) | 650.0 | 0.03 | |||
| Accipitriformes | ||||||||
| Accipitridae | Circaetus gallicus | 6 | 0.06% | 1200–2200 (1700.0) | 10,200.0 | 0.5 | ||
| Circus pygargus | 1 | 0.01% | 227–445 (336.0) | 336.0 | 0.02 | |||
| Buteo rufinus | 9 | 0.10% | 590–1760 (1175.0) | 10,575.0 | 0.5 | |||
| Buteo (buteo) vulpinus | 10 | 0.11% | 550–1300 (925.0) | 9250.0 | 0.5 | |||
| Accipiter nisus | 5 | 0.05% | 110–342 (226.0) | 1130.0 | 0.1 | |||
| Falconiformes | ||||||||
| Falconidae | Falco tinnunculus | 55 | 0.58% | 156–252 (204.0) | 11,220.0 | 0.6 | ||
| Falco naumanni | 1 | 0.01% | 90–208 (149.0) | 149.0 | 0.002 | |||
| Falco subbuteo | 2 | 0.02% | 131–340 (235.5) | 471.0 | 0.03 | |||
| Galliformes | ||||||||
| Phasianidae | Alectoris chukar | 418 | 4.42% | 360–560 (460.0) | 192,280.0 | 9.8 | ||
| Coturnix coturnix | 9 | 0.10% | 75–135 (105.0) | 945.0 | 0.05 | |||
| Gallus gallus domesticus | 3 | 0.03% | 1000 | 3000.0 | 0.2 | |||
| Gruiiformes | ||||||||
| Rallidae | Rallus aquaticus | 1 | 0.01% | 80–180 (130.0) | 130.0 | 0.002 | ||
| Porzana porzana | 1 | 0.01% | 70–110 (90.0) | 90.0 | 0.004 | |||
| Gallinula chloropus | 6 | 0.06% | 240–420 (330.0) | 1980.0 | 0.1 | |||
| Fulica atra | 7 | 0.07% | 700–1000 (850.0) | 5950.0 | 0.3 | |||
| Crex crex | 6 | 0.06% | 120–200 (160.0) | 960.0 | 0.05 | |||
| Charadriiformes | ||||||||
| Recurvirostridae | Himantopus himantopus | 2 | 0.02% | 150–210 (180.0) | 360.0 | 0.02 | ||
| Burhinidae | Burhinus oedicnemus | 48 | 0.51% | 430–500 (465.0) | 22,320.0 | 1.1 | ||
| Charadriidae | Vanellus spinosus | 37 | 0.39% | 127–159 (145.0) | 5365.0 | 0.3 | ||
| Scolopacidae | Tringa spp. | 4 | 0.04% | 40–70 (55.0) | 220.0 | 0.01 | ||
| Numenius arquata | 1 | 0.01% | 540–1300 (920.0) | 920.0 | 0.05 | |||
| Glareolidae | Cursorius cursor | 3 | 0.03% | 102–119 (110.5) | 331.5 | 0.02 | ||
| Laridae | Larus spp. | 1 | 0.01% | 600–940 (770.0) | 770.0 | 0.04 | ||
| Pterocliformes | ||||||||
| Pteroclidae | Pterocles orientalis | 1 | 0.01% | 300–550 (425.0) | 425.0 | 0.03 | ||
| Columbiformes | ||||||||
| Columbidae | Columba livia | 534 | 5.64% | 230–370 (280.0) | 149,520.0 | 7.7 | ||
| Streptopelia spp. | 54 | 0.57% | 80–240 (160.0) | 8640.0 | 0.4 | |||
| Cuculiformes | ||||||||
| Cuculidae | Clamator glandarius | 3 | 0.03% | 138–192 (165.0) | 495.0 | 0.03 | ||
| Strigiformes | ||||||||
| Tytonidae | Tyto alba | 71 | 0.75% | 240–350 (295.0) | 20,945.0 | 1.1 | ||
| Strigidae | Asio otus | 66 | 0.70% | 220–370 (295.0) | 19,470.0.0 | 1 | ||
| Athene noctua | 16 | 0.17% | 140–220 (180.0) | 2880.0 | 0.1 | |||
| Otus scops | 19 | 0.20% | 60–120 (90.0) | 1710.0 | 0.1 | |||
| Caprimulgiformes | ||||||||
| Caprimulgidae | Caprimulgus europaeus | 2 | 0.02% | 65–100 (82.5) | 165.0 | 0.002 | ||
| Bucerotiformes | ||||||||
| Upupidae | Upupa epops | 2 | 0.02% | 47–87 (67.0) | 134.0 | 0.002 | ||
| Coraciiformes | ||||||||
| Alcedinidae | Halcyon smyrnensis | 3 | 0.03% | 85–110 (97.5) | 292.5 | 0.003 | ||
| Meropidae | Merops apiaster | 11 | 0.12% | 44–78 (61.0) | 671.0 | 0.03 | ||
| Coraciidae | Coracias garrulus | 24 | 0.25% | 120–160 (140.0) | 3360.0 | 0.2 | ||
| Psittaciformes | ||||||||
| Psittaculidae | Psittacula krameri | 2 | 0.02% | 96–139 (120.0) | 240.0 | 0.003 | ||
| Passeriformes | Passeriformes spp. | 62 | 0.66% | 10–25 (17.5) | 1085.0 | 0.01 | ||
| Alaudidae | Galerida cristata | 1 | 0.01% | 37–55 (46.0) | 46.0 | 0.005 | ||
| Motacillidae | Motacilla alba | 1 | 0.01% | 17–25 (21.0) | 21.0 | 0.003 | ||
| Turdidae | Turdus merula | 1 | 0.01% | 80–125 (102.5) | 102.5 | 0.001 | ||
| Laniidae | Lanius senator | 2 | 0.02% | 30–40 (35.0) | 70.0 | 0.008 | ||
| Lanius nubicus | 1 | 0.01% | 22–30 (26.0) | 26.0 | 0.0003 | |||
| Lanius excubitor | 2 | 0.02% | 48–81 (64.5) | 129.0 | 0.002 | |||
| Corvidae | Garrulus glandarius | 41 | 0.43% | 140–190 (165.0) | 6765.0 | 0.08 | ||
| Coloeus monedula | 538 | 5.69% | 180–260 (220.0) | 118,360.0 | 6.1 | |||
| Corvus cornix | 140 | 1.48% | 430–650 (540.0) | 75,600.0 | 3.9 | |||
| Oriolidae | Oriolus oriolus | 1 | 0.01% | 56–79 (67.5) | 67.5 | 0.008 | ||
| Reptiles | Squamata | |||||||
| Boidae | Eryx jaculus | 77 | 0.81% | 350 | 26,950.0 | 1.4 | ||
| Viperidae | Daboia palaestinae | 4 | 0.04% | 1500 | 6000.0 | 0.3 | ||
| Colubridae | Rhynchocalamus melanocephalus | 1 | 0.01% | 20 | 20.0 | 0.002 | ||
| Hemorrhois nummifer | 1 | 0.01% | 600 | 600.0 | 0.003 | |||
| Lamprophiidae | Micrelaps muelleri | 2 | 0.04% | 25 | 50.0 | 0.003 | ||
| Testudines | ||||||||
| Testudinidae | Testudo graeca | 2 | 0.02% | 60 | 120.0 | 0.002 | ||
| Amphibian | Anura | |||||||
| Bufonidae | Bufotes variabilis | 1 | 0.01% | 25 | 25.0 | 0.002 | ||
| Invertebrata | ||||||||
| Insecta | Insecta spp. | 2 | 0.02% | |||||
| Gryllotalpidae | Gryllotalpa gryllotalpa | 21 | 0.22% | |||||
| Cerambycidae | Cerambyx dux | 6 | 0.06% | |||||
| Orthoptera spp. | 6 | 0.06% | ||||||
| Coleoptera spp. | 5 | 0.05% | ||||||
| Arachnida | Scorpiones | Scorpiones sp. | 1 | 0.01% | ||||
| Buthidae | Leiurus hebraeus | 2 | 0.02% | |||||
| Buthotus judaicus | 1 | 0.01% | ||||||
| Androctonus bicolor | 2 | 0.02% | ||||||
| Scorpionidae | Scorpio palmatus | 3 | 0.03% | |||||
| Scorpio fuscus | 144 | 1.52% | ||||||
| Diplocentridae | Nebo hierichonticus | 10 | 0.11% | |||||
| Solifugae | Solifugae spp. | 17 | 0.18% | |||||
| Crustacea | Potamidae | Potamon potamios | 2 | 0.02% | ||||
| Total Prey | 9461 | |||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hadad, E.; Charter, M.; Kosicki, J.Z.; Yosef, R. Prey-Base Does Not Influence Breeding Success in Eagle Owls (Bubo bubo) in Judea, Israel. Animals 2022, 12, 1280. https://doi.org/10.3390/ani12101280
Hadad E, Charter M, Kosicki JZ, Yosef R. Prey-Base Does Not Influence Breeding Success in Eagle Owls (Bubo bubo) in Judea, Israel. Animals. 2022; 12(10):1280. https://doi.org/10.3390/ani12101280
Chicago/Turabian StyleHadad, Ezra, Motti Charter, Jakub Z. Kosicki, and Reuven Yosef. 2022. "Prey-Base Does Not Influence Breeding Success in Eagle Owls (Bubo bubo) in Judea, Israel" Animals 12, no. 10: 1280. https://doi.org/10.3390/ani12101280
APA StyleHadad, E., Charter, M., Kosicki, J. Z., & Yosef, R. (2022). Prey-Base Does Not Influence Breeding Success in Eagle Owls (Bubo bubo) in Judea, Israel. Animals, 12(10), 1280. https://doi.org/10.3390/ani12101280