
First Insights on the Karyotype Diversification of the Endemic Malagasy Leaf-Toed Geckos (Squamata: Gekkonidae: Uroplatus)





Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Sampling
2.2. Molecular Analysis
2.3. Cytogenetic Analysis
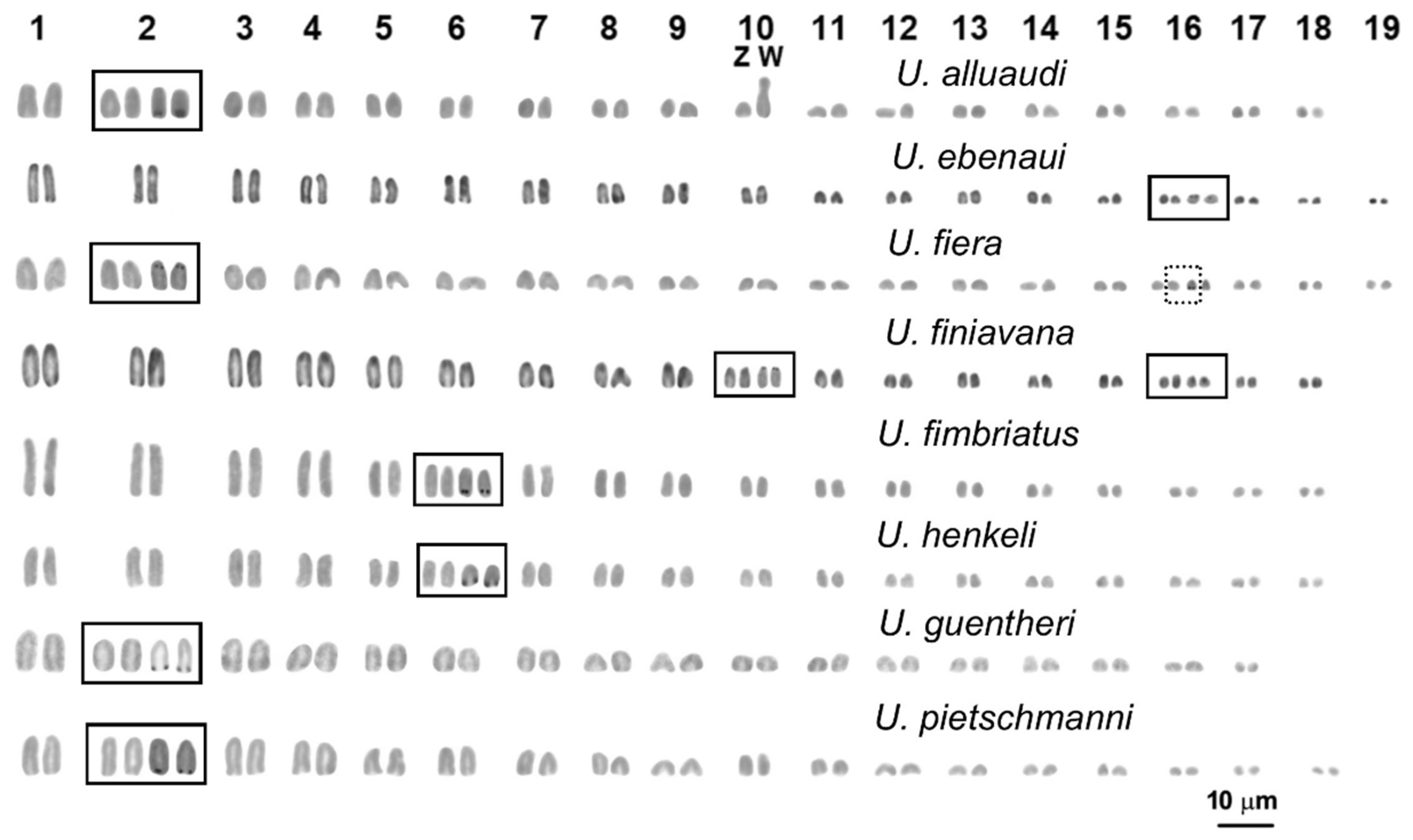
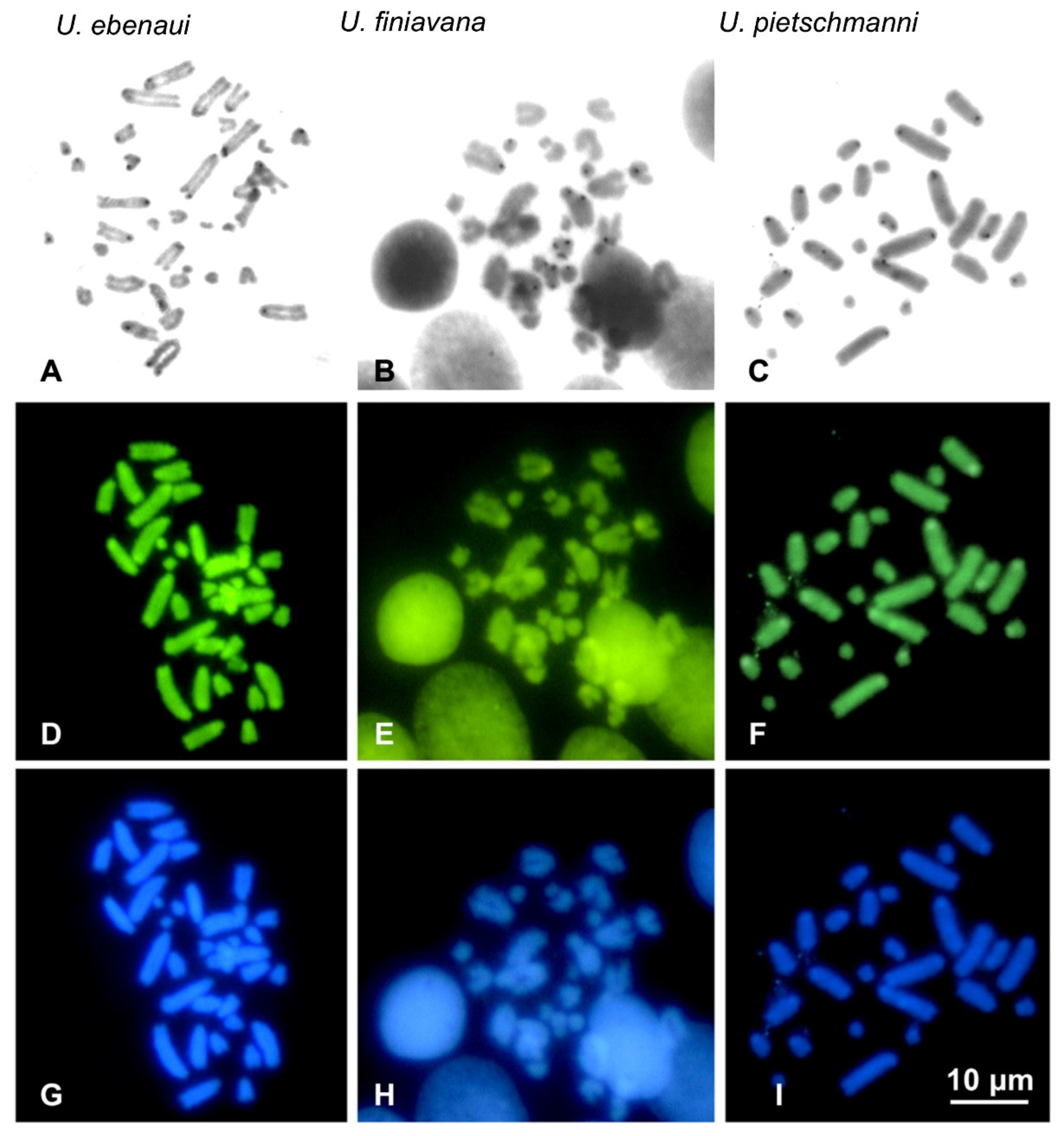
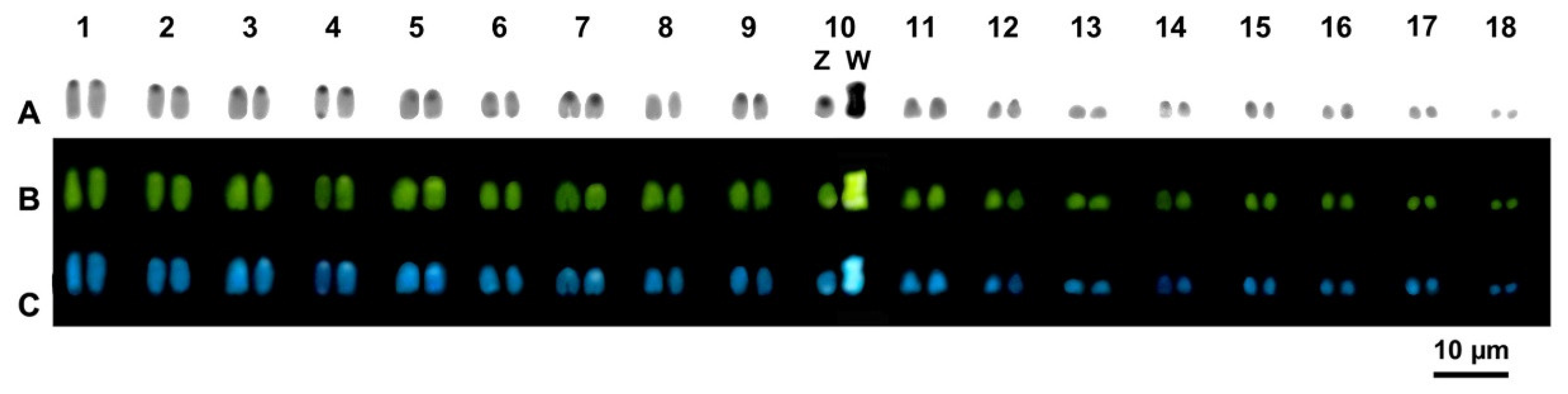
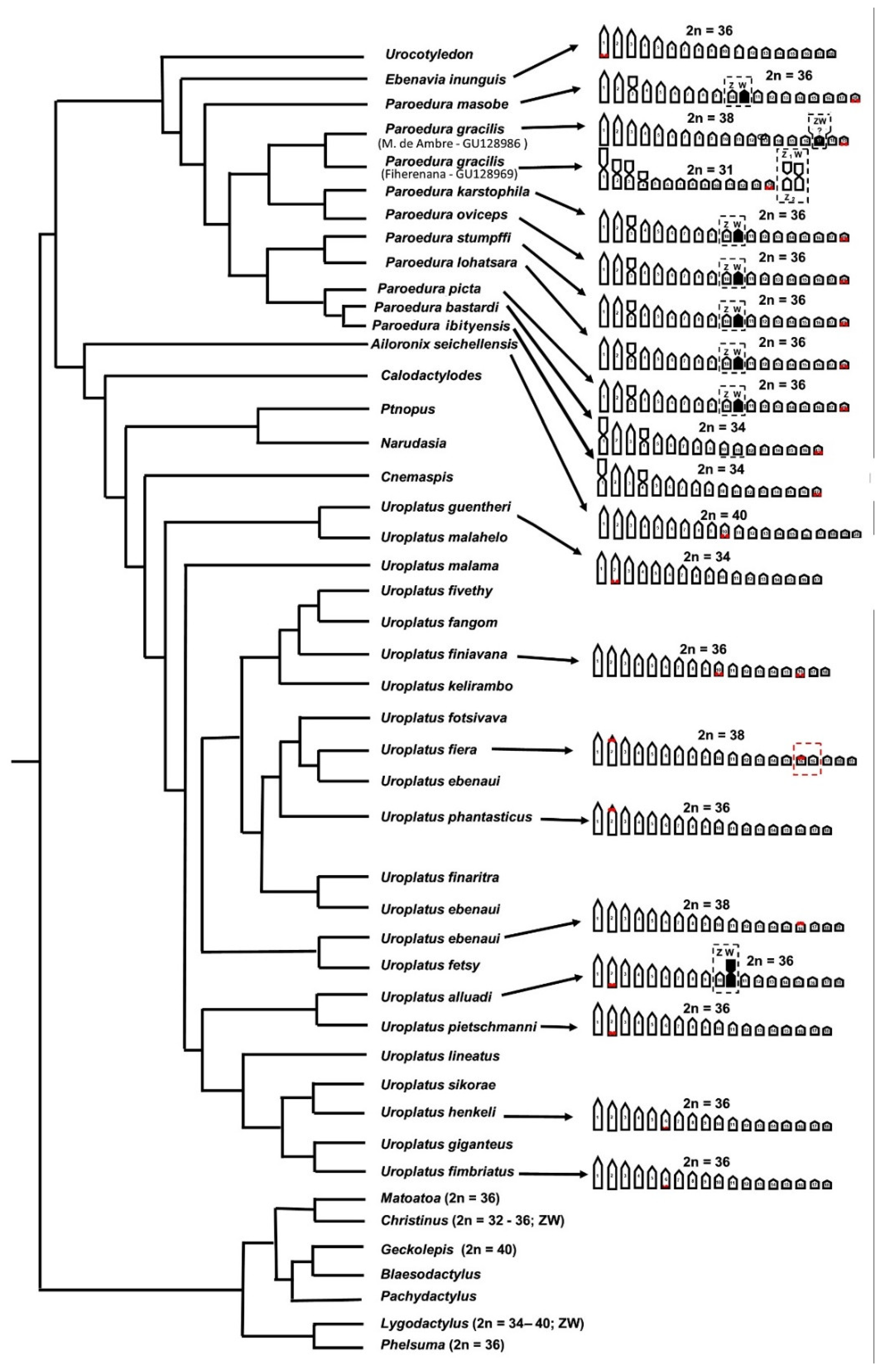
3. Results
Cytogenetic Analysis
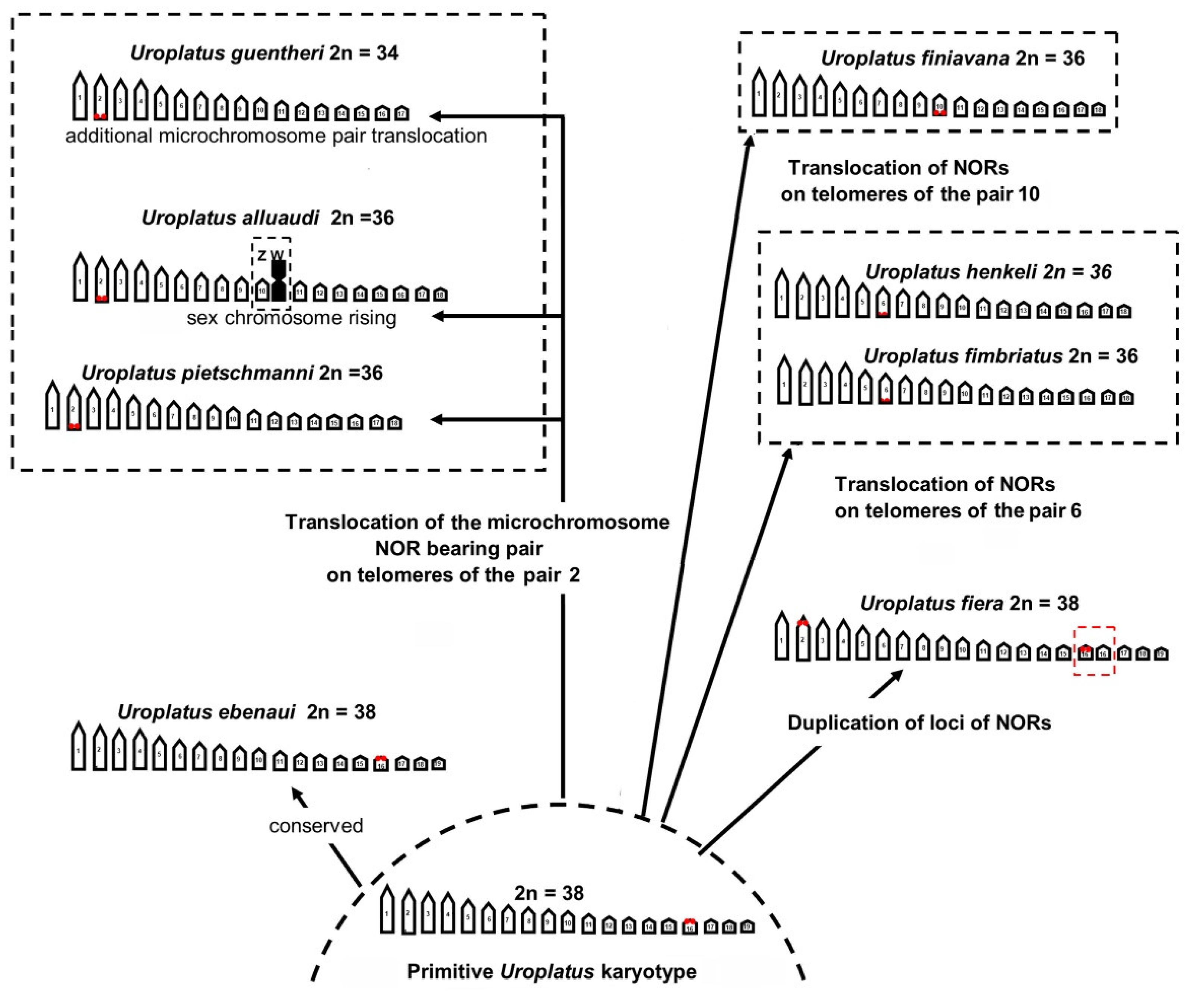
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hot-spots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Ganzhorn, J.U.; Lowry, P.P.; Schatz, G.E.; Sommer, S. The biodiversity of Madagascar: One of the world’s hottest hotspots on its way out. Oryx 2001, 35, 346–348. [Google Scholar] [CrossRef]
- Vences, M.; Wollenberg, K.C.; Vieites, D.R.; Lees, D.C. Madagascar as a model region of species diversification. Trends Ecol. Evol. 2009, 24, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Glaw, F.; Vences, M. A Field Guide to the Amphibians and Reptiles of Madagascar, 3rd ed.; Vences, M., Glaw, F., Eds.; Verlags GbR: Köln, Germany, 2007; p. 496. [Google Scholar]
- Uetz, P.; Freed, P.; Hošek, J. (Eds.) The Reptile Database. Available online: http://www.reptile-database.org (accessed on 9 June 2022).
- Koubová, M.; Pokorná, M.J.; Rovatsos, M.; Farkačová, K.; Altmanová, M.; Kratochvíl, L. Sex determination in Madagascar geckos of the genus Paroedura (Squamata: Gekkonidae): Are differentiated sex chromosomes indeed so evolutionary stable? Chromosom. Res. 2014, 22, 441–452. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Andreone, F.; Branch, W.R.; Glaw, F.; Guarino, F.M.; Nagy, Z.T.; Odierna, G.; Aprea, G. Chromosome evolution in pseudoxyrhophiine snakes from Madagascar: A wide range of karyotypic variability. Biol. J. Linn. Soc. 2014, 112, 450–460. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Andreone, F.; Aprea, G.; Glaw, F.; Odierna, G.; Guarino, F.M. Molecular phylogeny, biogeography and chromosome evolution of Malagasy dwarf geckos of the genus Lygodactylus (Squamata, Gekkonidae). Zool. Scr. 2017, 46, 42–54. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Andreone, F.; Glaw, F.; Odierna, G.; Petraccioli, A.; Guarino, F.M. Chromosome aneupolyploidy in an endemic Malagasy gecko (Gekkonidae: Geckolepis). Salamandra 2018, 54, 56–62. [Google Scholar]
- Mezzasalma, M.; Andreone, F.; Glaw, F.; Guarino, F.M.; Odierna, G.; Petraccioli, A.; Picariello, O. Changes in heterochromatin content and ancient chromosome fusion in the endemic Malagasy boid snakes Sanzinia and Acrantophis (Squamata: Serpentes). Salamandra 2019, 55, 140–144. [Google Scholar]
- Rovatsos, M.; Pokorná, M.J.; Altmanová, M.; Kratochvíl, L. Female heterogamety in Madagascar chameleons (Squamata: Chamaeleonidae: Furcifer): Differentiation of sex and neo-sex chromosomes. Sci. Rep. 2014, 5, 13196. [Google Scholar] [CrossRef]
- Rovatsos, M.; Altmanová, M.; Johnson Pokorná, M.; Velenský, P.; Sánchez Baca, A.; Kratochvíl, L. Evolution of karyotypes in chameleons. Genes 2017, 8, 382. [Google Scholar] [CrossRef]
- Glaw, F.; Kosuch, J.; Henkel, F.W.; Sound, P.; Böhme, W. Genetic and morphological variation of the leaf-tailed gecko Uroplatus fimbriatus from Madagascar, with description of a new giant species. Salamandra 2006, 42, 129144. [Google Scholar]
- Greenbaum, E.; Bauer, A.M.; Jackman, T.R.; Vences, M.; Glaw, F. A phylogeny of the enigmatic Madagascan geckos of the genus Uroplatus (Squamata: Gekkonidae). Zootaxa 2007, 1493, 41–51. [Google Scholar] [CrossRef]
- Raxworthy, C.J.; Pearson, R.G.; Zimkus, B.M.; Reddy, S.; Deo, A.J.; Nussbaum, R.A.; Ingram, C.M. Continental speciation in the tropics: Contrasting biogeographic patterns of divergence in the Uroplatus leaf-tailed gecko radiation of Madagascar. J. Zool. 2008, 275, 423–440. [Google Scholar] [CrossRef]
- Ratsoavina, F.M.; Louis, E.E., Jr.; Crottini, A.; Randrianiaina, R.-D.; Glaw, F.; Vences, M. A new leaf tailed gecko species from northern Madagascar with a preliminary assessment of molecular and morphological variability in the Uroplatus ebenaui group. Zootaxa 2011, 3022, 39–57. [Google Scholar] [CrossRef]
- Ratsoavina, F.M.; Vences, M.; Louis, E.E., Jr. Phylogeny and phylogeography of the Malagasy leaf-tailed geckos in the Uroplatus ebenaui group. Afr. J. Herpetol. 2012, 61, 143–158. [Google Scholar] [CrossRef]
- Ratsoavina, F.M.; Raminosoa, N.R.; Louis, E.E., Jr.; Raselimanana, A.P.; Glaw, F.; Vences, M. An overview of Madagascar’s leaf tailed geckos (genus Uroplatus): Species boundaries, candidate species and review of geographical distribution based on molecular data. Salamandra 2013, 49, 115–148. [Google Scholar]
- Ratsoavina, F.M.; Ranjanaharisoa, F.A.; Glaw, F.; Raselimanana, A.P.; Miralles, A.; Vences, M. A new leaf-tailed gecko of the Uroplatus ebenaui group (Squamata: Gekkonidae) from Madagascar’s central eastern rainforests. Zootaxa 2015, 4006, 143–160. [Google Scholar] [CrossRef]
- Ratsoavina, F.M.; Gehring, P.-S.; Scherz, M.D.; Vieites, D.R.; Glaw, F.; Vences, M. Two new species of leaf-tailed geckos (Uroplatus) from the Tsaratanana mountain massif in northern Madagascar. Zootaxa 2017, 4347, 446–464. [Google Scholar] [CrossRef]
- Ratsoavina, F.M.; Raselimanana, A.P.; Scherz, M.D.; Rakotoarison, A.; Razafindraibe, J.H.; Glaw, F.; Vences, M. Finaritra! A new leaf-tailed gecko (Uroplatus) species from Marojejy National Park in north-eastern Madagascar. Zootaxa 2019, 4545, 563–577. [Google Scholar] [CrossRef]
- Ratsoavina, F.M.; Scherz, M.D.; Tolley, K.A.; Raselimanana, A.P.; Glaw, F.; Vences, M. A new species of Uroplatus (Gekkonidae) from Ankarana National Park, Madagascar, of remarkably high genetic divergence. Zootaxa 2019, 4683, 84–96. [Google Scholar] [CrossRef]
- Ratsoavina, F.M.; Glaw, F.; Raselimanana, A.P.; Rakotoarison, A.; Vieites, D.R.; Hawlitschek, O.; Vences, M.; Scherz, M.D. Towards completion of the species inventory of small-sized leaf-tailed geckos: Two new species of Uroplatus from northern Madagascar. Zootaxa 2020, 4895, 251–271. [Google Scholar] [CrossRef] [PubMed]
- Aprea, G.; Andreone, F.; Fulgione, D.; Petraccioli, A.; Odierna, G. Chromosomal Rearrangements Occurred Repeatedly and Independently during Species Diversification in Malagasy Geckos, genus Paroedura. Afr. Zool. 2013, 48, 96–108. [Google Scholar] [CrossRef]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- King, M.; Rofe, R. Karyotypic variation in the Australian Gekko Phyllodactylus marmoratus (Gray) (Gekkonidae: Reptilia). Chromosoma 1976, 54, 75–87. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Lab Press: New York, NY, USA, 1989. [Google Scholar]
- Kocher, T.D.; Thomas, W.K.; Meyer, A.; Edwards, S.V.; Paabo, S.; Villablanca, F.X.; Wilson, A.C. Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA 1989, 86, 6196–6200. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Jeong, T.J.; Jun, J.; Han, S.; Kim, H.T.; Oh, K.; Kwak, M. DNA barcode reference data for the Korean herpetofauna and their applications. Mol. Ecol. Resour. 2013, 13, 1019–1032. [Google Scholar] [CrossRef]
- Nugraha, F.A.D.; Fatchiyah, F.; Smith, E.N.; Kurniawan, N. Phylogenetic analysis of colubrid snakes based on 12S rDNA reveals distinct lineages of Dendrelaphis pictus (Gmelin, 1789) populations in Sumatra and Java. Biodiversitas 2018, 19, 303–310. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Visone, V.; Petraccioli, A.; Odierna, G.; Capriglione, T.; Guarino, F.M. Non-random accumulation of LINE1-like sequences on differentiated snake W chromosomes. J. Zool. 2016, 300, 67–75. [Google Scholar] [CrossRef]
- Howell, W.M.; Black, D.A. Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: A 1-step method. Experientia 1980, 36, 1014–1015. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Andreone, F.; Odierna, G.; Guarino, F.M.; Crottini, A. Comparative cytogenetics on eight Malagasy Mantellinae (Anura, Mantellidae) and a synthesis of the karyological data on the subfamily. Comp. Cytogenet. 2022, 16, 1–17. [Google Scholar] [CrossRef]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for Centromeric Position on Chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Porter, C.A.; Hamilton, M.J.; Sites, J.W., Jr.; Baker, R.J. Location of ribosomal DNA in chromosomes of squamate reptiles: Systematic and evolutionary implications. Herpetologica 1991, 47, 271–280. [Google Scholar]
- Altmanová, M.; Rovatsos, M.; Kratochvíl, L.; Johnson Pokorná, M. Minute Y chromosomes and karyotype evolution in Madagascan iguanas (Squamata: Iguania: Opluridae). Biol. J. Linn. Soc. 2016, 118, 618–633. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Guarino, F.M.; Odierna, G. Lizards as Model Organisms of Sex Chromosome Evolution: What We Really Know from a Systematic Distribution of Available Data? Genes 2021, 12, 1341. [Google Scholar] [CrossRef]
- Stults, D.M.; Killen, M.W.; Williamson, E.P.; Hourigan, J.S.; Vargas, H.D.; Arnold, S.M.; Moscow, J.A.; Pierce, A.J. Human rRNA Gene Clusters Are Recombinational Hotspots in Cancer. Cancer Res. 2009, 69, 9096–9104. [Google Scholar] [CrossRef]
- McStay, B. Nucleolar organizer regions: Genomic ‘dark matter’ requiring illumination. Genes Dev. 2016, 30, 1598–1610. [Google Scholar] [CrossRef]
- Odierna, G.; Capriglione, T.; Olmo, E.; Cardone, A.; Rosati, C. The karyology of some South African lacertids belonging to three genera Heliobolus, Meroles, and Pedioplanis. J. Afr. Zool. 1990, 104, 541–547. [Google Scholar]
- Odierna, G.; Olmo, E.; Capriglione, T.; Caputo, V. Karyological Differences between Lacerta lepida and Lacerta pater. S. Am. J. Herpetol. 1990, 24, 97–99. [Google Scholar] [CrossRef]
- Mezzasalma, M.; Guarino, F.; Loader, S.; Odierna, G.; Streicher, J.; Cooper, N. First karyological analysis of the endemic Malagasy phantom gecko Matoatoa brevipes (Squamata: Gekkonidae). Acta Herpetol. 2020, 15, 137–141. [Google Scholar] [CrossRef]
- White, M.J.D. Animal Cytology and Evolution, 3rd ed.; Cambridge University Press: Cambridge, UK, 1973. [Google Scholar]
- Aprea, G.; Odierna, G.; Capriglione, T.; Caputo, V.; Morescalchi, A.; Olmo, E. Heterochromatin and NOR distribution in the chromosomes of six gekkonid species of the genus Phelsuma (Squamata: Gekkonidae). J. Afr. Zool. 1996, 119, 341–349. [Google Scholar]
- Volobouev, V.; Ineich, I. A chromosome banding study of Ailuronyx seychellensis (Reptilia, Gekkonidae). J. Herpetol. 1994, 28, 267–270. [Google Scholar] [CrossRef]
- King, M.; King, D. An additional chromosome race of Phyllodactyllus marmoratus (Gray) (Reptilia: Gekkonidae) and its phylogenetic implications. Aust. J. Zool. 1977, 25, 667–672. [Google Scholar] [CrossRef]
- Olmo, E. Trends in the evolution of reptilian chromosomes. Integr. Comp. Biol. 2008, 48, 486–493. [Google Scholar] [CrossRef]
- Beçak, W. Constititução cromossomica e mecanismo de determinação do sexo em ofidios sudamericanos. I. Aspectos cariotipicos. Mem. Do Inst. Butantan 1965, 32, 37–78. [Google Scholar]
- Wright, A.E.; Dean, R.; Zimmer, F.; Mank, J.E. How to make a sex chromosome. Nat. Commun. 2016, 7, 12087. [Google Scholar] [CrossRef]
- Ohno, S. Sex Chromosomes and Sex-Linked Genes; Springer: Berlin, Germany, 1967. [Google Scholar]
- Charlesworth, B. The evolution of sex chromosomes. Science 1991, 251, 1030–1033. [Google Scholar] [CrossRef]
- Pallotta, M.M.; Turano, M.; Ronca, R.; Mezzasalma, M.; Petraccioli, A.; Odierna, G.; Capriglione, T. Brain Gene Expression is Influenced by Incubation Temperature During Leopard Gecko (Eublepharis macularius) Development. J. Exp. Zool. Mol. Dev. Evol. 2017, 328, 360–370. [Google Scholar] [CrossRef]
- Sidhom, M.; Said, K.; Chatti, N.; Guarino, F.M.; Odierna, G.; Petraccioli, A.; Picariello, O.; Mezzasalma, M. Karyological characterization of the common chameleon (Chamaeleo chamaeleon) provides insights on the evolution and diversification of sex chromosomes in Chamaeleonidae. Zoology 2020, 141, 125738. [Google Scholar] [CrossRef]
- Charlesworth, D. Sex differences in fitness and selection for centric fusions between sex-chromosomes and autosomes. Genet. Res. 1980, 35, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Farkačová, K.; Altmanová, M.; Johnson Pokorná, M.J.; Kratochvíl, L. The rise and fall of differentiated sex chromosomes in geckos. Mol. Ecol. 2019, 28, 3042–3052. [Google Scholar] [CrossRef] [PubMed]
- Thépot, D. Sex Chromosomes and Master Sex-Determining Genes in Turtles and Other Reptiles. Genes 2021, 12, 1822. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | FN | Sex | Locality | Max Identity |
|---|---|---|---|---|
| U. alluaudi Mocquard, 1894 | GA 476 | female | Montagne d’Ambre | 100% vs. KF160464 |
| U. henkeli Böhme & Ibisch, 1990 | GA 477 | male | Montagne d’Ambre | 99.3% vs. JX205281 |
| U. henkeli | GA 1099 | male | Montagne d’Ambre | 99.3% vs. JX205281 |
| U. ebenaui (Boettger, 1879) | FGMV 2205 | female | Manongarivo | 99.4% vs. JX205278 |
| U. ebenaui | GA 1100 | female | NA | 99.4% vs. JX205278 |
| U. fiera Ratsoavina, Ranjanaharisoa, Glaw, Raselimanana, Miralles & Vences, 2015 | FGMV 3097 | male | Fiherenana region | 100% vs. JX205263 |
| U. fiera | GA 140 | juvenile | Fiherenana region | 100% vs. JX205263 |
| U. finiavana Ratsoavina, Louis Jr., Crottini, Randrianiaina, Glaw & Vences, 2011 | FGMV 3084 | male | Montagne d’Ambre | 100% vs. MW035835 |
| U. finiavana | GA 1100 | juvenile | Montagne d’Ambre | 100% vs. MW035835 |
| U. fimbriatus (Schneider, 1797) | FGMV 2234 | male | NA | 99.5% vs. AB612276 |
| U. prope guentheri Mocquard, 1908 | GA 328 | male | Marofandilia | 96.8% vs. EU596688 |
| U. prope guentheri | GA 329 | male | Marofandilia | 96.8% vs. EU596688 |
| U. pietschmanni Böhle & Schönecker, 2004 | FAZC 11627 | male | NosyBe | 99.7% vs. EU596687 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mezzasalma, M.; Brunelli, E.; Odierna, G.; Guarino, F.M. First Insights on the Karyotype Diversification of the Endemic Malagasy Leaf-Toed Geckos (Squamata: Gekkonidae: Uroplatus). Animals 2022, 12, 2054. https://doi.org/10.3390/ani12162054
Mezzasalma M, Brunelli E, Odierna G, Guarino FM. First Insights on the Karyotype Diversification of the Endemic Malagasy Leaf-Toed Geckos (Squamata: Gekkonidae: Uroplatus). Animals. 2022; 12(16):2054. https://doi.org/10.3390/ani12162054
Chicago/Turabian StyleMezzasalma, Marcello, Elvira Brunelli, Gaetano Odierna, and Fabio Maria Guarino. 2022. "First Insights on the Karyotype Diversification of the Endemic Malagasy Leaf-Toed Geckos (Squamata: Gekkonidae: Uroplatus)" Animals 12, no. 16: 2054. https://doi.org/10.3390/ani12162054
APA StyleMezzasalma, M., Brunelli, E., Odierna, G., & Guarino, F. M. (2022). First Insights on the Karyotype Diversification of the Endemic Malagasy Leaf-Toed Geckos (Squamata: Gekkonidae: Uroplatus). Animals, 12(16), 2054. https://doi.org/10.3390/ani12162054