
Addition of Organic Acids and Lactobacillus acidophilus to the Leguminous Forage Chamaecrista rotundifolia Improved the Quality and Decreased Harmful Bacteria of the Silage

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silage Preparation
2.2. Sampling and Chemical Analysis
2.3. DNA Extraction, Amplicon Library Preparation, and Sequencing
2.4. Sequence Processing and Analysis
2.5. Sampling and Chemical Analysis
3. Results and Discussion
3.1. Effect of Additives on the Nutrient Composition of Chamaecrista Rotundifolia Silage
3.2. Effect of Additives on the Fermentation Parameters of Chamaecrista rotundifolia
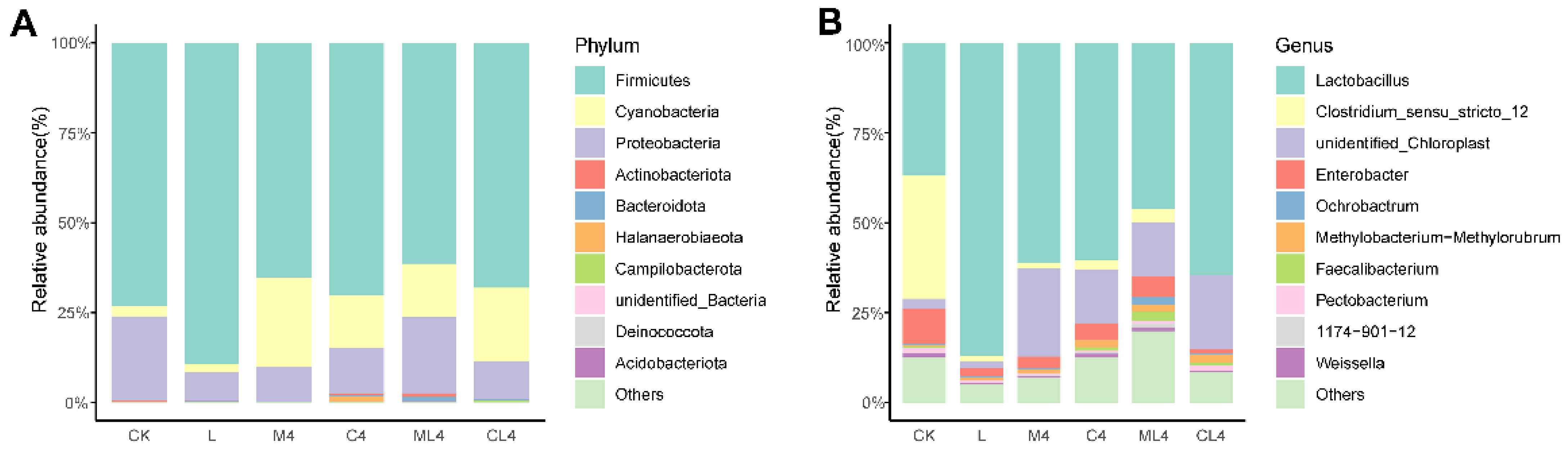
3.3. Microbial Diversity in Chamaecrista rotundifolia Silage
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Minggang, X.; McDonald, C.; Liu, C.; Hacker, J. Variation in temperature requirements for germination and early seedling root development in Chamaecrista rotundifolia and three allied species. Genet. Resour. Crop Evol. 2000, 47, 25–34. [Google Scholar] [CrossRef]
- Cruz, E.D.; Camarão, A.P.; Simão Neto, M. Forage production and nutritive value of Chamaecrista rotundifolia Persoon Greene in the eastern Amazon, Brazil. Pasturas Tropicales 1999, 21, 46–48. [Google Scholar]
- Xu, D.; Wang, N.; Rinne, M.; Ke, W.; Weinberg, Z.G.; Da, M.; Bai, J.; Zhang, Y.; Li, F.; Guo, X. The bacterial community and metabolome dynamics and their interactions modulate fermentation process of whole crop corn silage prepared with or without inoculants. Microb. Biotechnol. 2021, 14, 561–576. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, S.; Dong, Z.; Li, J.; Jia, Y.; Shao, T. Effect of storage time and the level of formic acid on fermentation characteristics, epiphytic microflora, carbohydrate components and in vitro digestibility of rice straw silage. Anim. Biosci. 2021, 34, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Li, J.; Wang, S.; Zhao, J.; Dong, D.; Shao, T. Gamma-ray irradiation and microbiota transplantation to separate the effects of chemical and microbial diurnal variations on the fermentation characteristics and bacterial community of napier grass silage. J. Sci. Food Agric. 2022, 102, 4322–4332. [Google Scholar] [CrossRef]
- Zhu, Y.; Xiong, H.; Wen, Z.; Tian, H.; Chen, Y.; Wu, L.; Guo, Y.; Sun, B. Effects of Different concentrations of Lactobacillus plantarum and Bacillus licheniformis on silage quality, in vitro fermentation and microbial community of hybrid Pennisetum. Animals 2022, 12, 1752. [Google Scholar] [CrossRef] [PubMed]
- Mahmood Fashandi, H.; Abbasi, R.; Mousavi Khaneghah, A. The detoxification of aflatoxin M1 by Lactobacillus acidophilus and Bifidobacterium spp.: A review. J. Food Process. Preserv. 2018, 42, e13704. [Google Scholar] [CrossRef]
- Anjum, N.; Maqsood, S.; Masud, T.; Ahmad, A.; Sohail, A.; Momin, A. Lactobacillus acidophilus: Characterization of the species and application in food production. Crit. Rev. Food Sci. Nutr. 2014, 54, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Pian, R.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Effects of citric acid on fermentation characteristics and bacterial diversity of Amomum villosum silage. Bioresour. Technol. 2020, 307, 123290. [Google Scholar] [CrossRef]
- Martin, S.; Sullivan, H.; Evans, J. Effect of sugars and malate on ruminal microorganisms. J. Dairy Sci. 2000, 83, 2574–2579. [Google Scholar] [CrossRef]
- El-Zaiat, H.M.; Kholif, A.E.; Mohamed, D.A.; Matloup, O.H.; Anele, U.Y.; Sallam, S.M. Enhancing lactational performance of Holstein dairy cows under commercial production: Malic acid as an option. J. Sci. Food Agric. 2019, 99, 885–892. [Google Scholar] [CrossRef]
- Tian, H.; Wang, Y.; Liu, Z.; Hu, Z.; Guo, Y.; Deng, M.; Liu, G.; Sun, B. Effects of malic acid and sucrose on the fermentation parameters, CNCPS nitrogen fractions, and bacterial community of Moringa oleifera leaves silage. Microorganisms 2021, 9, 2102. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jiang, D.; Zheng, M.; Tian, P.; Zheng, M.; Xu, C. Microbial community dynamics during alfalfa silage with or without clostridial fermentation. Sci. Rep. 2020, 10, 17782. [Google Scholar] [CrossRef]
- Guo, X.; Ke, W.; Ding, W.; Ding, L.; Xu, D.; Wang, W.; Zhang, P.; Yang, F.Y. Profiling of metabolome and bacterial community dynamics in ensiled Medicago sativa inoculated without or with Lactobacillus plantarum or Lactobacillus buchneri. Sci. Rep. 2018, 8, 357. [Google Scholar] [CrossRef] [PubMed]
- Thomas, T.A. An automated procedure for the determination of soluble carbohydrates in herbage. J. Sci. Food Agric. 1977, 28, 639–642. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Monti, A.; Di Virgilio, N.; Venturi, G. Mineral composition and ash content of six major energy crops. Biomass Bioenergy 2008, 32, 216–223. [Google Scholar] [CrossRef]
- Broderick, G.A.; Kang, J.H. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 1980, 63, 64–75. [Google Scholar] [CrossRef]
- Wang, S.; Sun, Y.; Zhao, J.; Dong, Z.; Li, J.; Nazar, M.; Shao, T. Assessment of inoculating various epiphytic microbiota on fermentative profile and microbial community dynamics in sterile Italian ryegrass. J. Appl. Microbiol. 2020, 129, 509–520. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ. 2016, 4, e2584. [Google Scholar] [CrossRef]
- Chen, T.; Liu, Y.X.; Huang, L. ImageGP: An easy-to-use data visualization web server for scientific researchers. iMeta 2022, 1, e5. [Google Scholar] [CrossRef]
- Tao, X.; Chen, S.; Zhao, J.; Wang, S.; Dong, Z.; Li, J.; Sun, F.; Shao, T. Effects of citric acid residue and lactic acid bacteria on fermentation quality and aerobic stability of alfalfa silage. Ital. J. Anim. Sci. 2020, 19, 744–752. [Google Scholar] [CrossRef]
- Ke, W.C.; Ding, W.R.; Xu, D.M.; Ding, L.M.; Zhang, P.; Li, F.D.; Guo, X.S. Effects of addition of malic or citric acids on fermentation quality and chemical characteristics of alfalfa silage. J. Dairy Sci. 2017, 100, 8958–8966. [Google Scholar] [CrossRef]
- Liu, Q.; Shao, T.; Bai, Y. The effect of fibrolytic enzyme, Lactobacillus plantarum and two food antioxidants on the fermentation quality, alpha-tocopherol and beta-carotene of high moisture napier grass silage ensiled at different temperatures. Anim. Feed Sci. Technol. 2016, 221, 1–11. [Google Scholar] [CrossRef]
- Zhang, Y.C.; Li, D.X.; Wang, X.K.; Lin, Y.L.; Zhang, Q.; Chen, X.Y.; Yang, F.Y. Fermentation dynamics and diversity of bacterial community in four typical woody forages. Ann. Microbiol. 2019, 69, 233–240. [Google Scholar] [CrossRef]
- Zhao, J.; Dong, Z.; Li, J.; Chen, L.; Bai, Y.; Jia, Y.; Shao, T. Ensiling as pretreatment of rice straw: The effect of hemicellulase and Lactobacillus plantarum on hemicellulose degradation and cellulose conversion. Bioresour. Technol. 2018, 266, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Desta, S.T.; Yuan, X.; Li, J.; Shao, T. Ensiling characteristics, structural and nonstructural carbohydrate composition and enzymatic digestibility of Napier grass ensiled with additives. Bioresour. Technol. 2016, 221, 447–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hristov, A.; Harper, M.; Roth, G.; Canale, C.; Huhtanen, P.; Richard, T.; DiMarco, K. Effects of ensiling time on corn silage neutral detergent fiber degradability and relationship between laboratory fiber analyses and in vivo digestibility. J. Dairy Sci. 2020, 103, 2333–2346. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, J.; Cui, X.; Stinner, W.; Guo, J.; Dong, R. Enhancement mechanism of biogas potential from lignocellulosic substrates in the ensiling process via acid-based hydrolysis and biological degradation. J. Clean. Prod. 2021, 319, 128826. [Google Scholar] [CrossRef]
- Muck, R.E. Silage microbiology and its control through additives. Rev. Bras. Zootec. 2010, 39, 183–191. [Google Scholar] [CrossRef]
- Li, P.; Ji, S.; Hou, C.; Tang, H.; Wang, Q.; Shen, Y. Effects of chemical additives on the fermentation quality and N distribution of alfalfa silage in south of China. Anim. Sci. J. 2016, 87, 1472–1479. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.; Ding, W.; Xu, D.; Shah, M.N.; Zhang, P.; Guo, X. Influences of addition of malic acid or citric acid, Lactobacillus plantarum and their mixtures on fermentation quality, proteolysis and fatty acid composition of ensiled alfalfa. Arch. Anim. Nutr. 2018, 72, 492–502. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Wang, X.; Lu, Y.; Guo, L.; Li, X.; Yang, F. Exploring the silage quality of alfalfa ensiled with the residues of astragalus and hawthorn. Bioresour. Technol. 2020, 297, 122249. [Google Scholar] [CrossRef]
- Dong, Z.; Shao, T.; Li, J.; Yang, L.; Yuan, X. Effect of alfalfa microbiota on fermentation quality and bacterial community succession in fresh or sterile Napier grass silages. J. Dairy Sci. 2020, 103, 4288–4301. [Google Scholar] [CrossRef]
- Yang, L.; Yuan, X.; Li, J.; Dong, Z.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef]
- Wang, C.; He, L.; Xing, Y.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Effects of mixing Neolamarckia cadamba leaves on fermentation quality, microbial community of high moisture alfalfa and stylo silage. Microb. Biotechnol. 2019, 12, 869–878. [Google Scholar] [CrossRef]
- Du, Z.; Lin, Y.; Sun, L.; Yang, F.; Cai, Y. Microbial community structure, co-occurrence network and fermentation characteristics of woody plant silage. J. Sci. Food Agric. 2022, 102, 1193–1204. [Google Scholar] [CrossRef]
- da Silva, E.B.; Smith, M.L.; Savage, R.M.; Polukis, S.A.; Drouin, P.; Kung, L., Jr. Effects of Lactobacillus hilgardii 4785 and Lactobacillus buchneri 40788 on the bacterial community, fermentation and aerobic stability of high-moisture corn silage. J. Appl. Microbiol. 2021, 130, 1481–1493. [Google Scholar] [CrossRef]
- Li, M.; Zhang, L.; Zhang, Q.; Zi, X.; Lv, R.; Tang, J.; Zhou, H. Impacts of citric acid and malic acid on fermentation quality and bacterial community of cassava foliage silage. Front. Microbiol. 2020, 11, 595622. [Google Scholar] [CrossRef]
- Tian, H.; Zhu, Y.; Dai, M.; Li, T.; Guo, Y.; Deng, M.; Sun, B. Additives altered bacterial communities and metabolic profiles in silage hybrid pennisetum. Front. Microbiol. 2022, 12, 770728. [Google Scholar] [CrossRef]
- Ding, Z.; Bai, J.; Xu, D.; Li, F.; Zhang, Y.; Guo, X. Microbial community dynamics and natural fermentation profiles of ensiled alpine grass Elymus nutans prepared from different regions of the Qinghai-Tibetan Plateau. Front. Microbiol. 2020, 11, 855. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z. Bacterial Dynamics of Wheat Silage. Front. Microbiol. 2019, 10, 1532. [Google Scholar] [CrossRef]
- Liu, B.; Huan, H.; Gu, H.; Xu, N.; Shen, Q.; Ding, C. Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 2019, 273, 212–219. [Google Scholar] [CrossRef]
- Ren, H.; Feng, Y.; Pei, J.; Li, J.; Wang, Z.; Fu, S.; Zheng, Y.; Li, Z.; Peng, Z. Effects of Lactobacillus plantarum additive and temperature on the ensiling quality and microbial community dynamics of cauliflower leaf silages. Bioresour. Technol. 2020, 307, 123238. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, H.; Yu, Z. Effects of sucrose, formic acid and lactic acid bacteria inoculant on quality, in vitro rumen digestibility and fermentability of drooping wild ryegrass (Elymus nutans Griseb.) silage. J. Anim. Feed Sci. 2017, 26, 26–32. [Google Scholar] [CrossRef]
- Ávila, C.; Carvalho, B. Silage fermentation—updates focusing on the performance of micro-organisms. J. Appl. Microbiol. 2020, 128, 966–984. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Magaji, U.; Hussin, G.; Ramli, A.; Miah, G. Fermentation quality and additives: A case of rice straw silage. Biomed Res. Int. 2016, 2016, 7985167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Nishino, N. Effects of inoculation of Lactobacillus rhamnosus and Lactobacillus buchneri on fermentation, aerobic stability and microbial communities in whole crop corn silage. Grassl. Sci. 2011, 57, 184–191. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item and Ensiling Days | Treatments | SEM | P | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | L | M1 | M2 | M3 | M4 | C1 | C2 | C3 | C4 | D | T | D × T | ||
| DM (g/kg FW) | ||||||||||||||
| 30 | 245.1 c | 267.3 a | 256.5 b | 262.5 ab | 266.4 ab | 271.8 a | 258.6 b | 263.1 ab | 268.5 ab | 272.1 a | 0.972 | 0.003 | <0.001 | 1.000 |
| 45 | 243.6 c | 262.8 ab | 252.6 bc | 255.0 b | 260.4 ab | 267.6 a | 253.8 bc | 255.6 b | 264.0 ab | 267.6 a | ||||
| 60 | 246.1 c | 262.6 a | 249.9 bc | 257.0 abc | 261.3 ab | 266.5 a | 250.8 bc | 257.9 ab | 263.2 a | 267.4 a | ||||
| CP (g/kg DM) | ||||||||||||||
| 30 | 139.0 Ac | 163.6 ab | 141.2 Ac | 156.7 Aab | 161.5 ab | 168.6 a | 152.4 Abc | 156.6 ab | 163.3 ab | 169.4 a | 1.615 | <0.001 | <0.001 | <0.001 |
| 45 | 124.8 ABd | 159.5 a | 136.4 ABcd | 145.9 Bbc | 156.5 ab | 163.2 a | 138.8 Bc | 143.7 c | 157.1 ab | 165.1 a | ||||
| 60 | 115.1 Bb | 153.2 a | 123.2 Bb | 143.0 Ba | 152.3 a | 155.0 a | 125.2 Cb | 143.9 a | 153.4 a | 156.4 a | ||||
| WSC (g/kg DM) | ||||||||||||||
| 30 | 11.1 bc | 8.2 c | 10.4 Abc | 11.3 bc | 11.4 bc | 16.9 Aa | 10.6 bc | 12.1 Abc | 13.0 ab | 13.3 ab | 0.339 | <0.001 | <0.001 | 0.775 |
| 45 | 10.3 bcd | 7.9 d | 9.4 ABcd | 10.4 bcd | 15.4 a | 15.1 ABa | 10.3 bcd | 12.5 Aabc | 13.9 ab | 13.0 abc | ||||
| 60 | 9.7 abcd | 5.0 f | 6.1 Bef | 8.0 cde | 9.3 bcd | 11.8 Bab | 7.2 def | 8.1 Bcde | 10.6 abc | 12.1 a | ||||
| NDF (g/kg DM) | ||||||||||||||
| 30 | 638.1 Ba | 613.5 Abc | 646.5 Aa | 640.8 Aa | 624.3 Ab | 551.0 Bf | 620.3 Bb | 604.0 Bcd | 598.8 Bde | 591.7 Ade | 4.035 | <0.001 | <0.001 | <0.001 |
| 45 | 652.5 Ab | 619.7 Ade | 671.0 Aa | 652.2 Ab | 630.5 Acd | 610.9 Aef | 639.3 Abc | 628.0 Acd | 625.5 Acd | 599.6 Ae | ||||
| 60 | 563.8 Cc | 522.6 Be | 604.9 Ba | 597.6 Bab | 594.9 Bab | 553.1 Bcd | 582.4 Cb | 549.9 Ccd | 546.5 Ccd | 542.5 Bd | ||||
| ADF (g/kg DM) | ||||||||||||||
| 30 | 439.1 Bbc | 405.8 Bf | 469.4 Aa | 436.3 Bbc | 430.0 Bcd | 405.4 Bf | 448.9 Ab | 406.2 Bf | 419.7 Bde | 415.2 ABef | 2.300 | <0.001 | <0.001 | <0.001 |
| 45 | 450.6 Ab | 435.3 Ac | 464.6 Aa | 453.8 Ab | 447.6 Ab | 414.1 Ad | 446.5 Ab | 434.7 Ac | 432.2 Ac | 425.7 Ac | ||||
| 60 | 426.6 Ca | 391.0 Bd | 435.3 Ba | 407.2 Cbc | 404.6 Cc | 376.0 Ce | 426.4 Ba | 413.0 Bbc | 414.3 Bb | 409.4 Bbc | ||||
| Ash (g/kg DM) | ||||||||||||||
| 30 | 57.7 | 56.6 A | 52.2 B | 52.9 B | 52.9 | 53.7 | 53.7 B | 53.7 A | 55.1 A | 56.5 A | 0.444 | <0.001 | 0.030 | <0.001 |
| 45 | 61.2 a | 57.7 Ab | 57.8 Ab | 57.2 Ab | 53.7 c | 52.7 c | 62.7 Aa | 57.5 Ab | 57.1 Ab | 53.4 Bc | ||||
| 60 | 54.4 a | 48.4 Bbcd | 50.7 Bb | 52.5 Babc | 52.1 bcd | 52.9 abc | 44.8 Ce | 50.3 Bbcd | 50.9 Bab | 53.5 Bab | ||||
| Item and Ensiling Days | Treatments | SEM | p | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | L | ML1 | ML2 | ML3 | ML4 | CL1 | CL2 | CL3 | CL4 | D | T | D × T | ||
| DM (g/kg FW) | ||||||||||||||
| 30 | 245.1 c | 267.3 a | 261.6 b | 263.7 b | 269.7 ab | 277.8 a | 262.2 b | 265.2 b | 271.2 ab | 278.4 a | 1.072 | 0.001 | <0.001 | 1.000 |
| 45 | 243.6 c | 262.8 ab | 255.0 bc | 257.4 b | 264.6 ab | 268.2 ab | 255.0 bc | 259.2 ab | 266.4 ab | 271.1 a | ||||
| 60 | 246.1 c | 262.6 a | 252.2 bc | 258.4 abc | 263.6 ab | 268.4 a | 252.7 bc | 259.4 ab | 264.1 ab | 271.2 a | ||||
| CP (g/kg DM) | ||||||||||||||
| 30 | 139.0 Ac | 163.6 ab | 153.8 bc | 161.0 ab | 165.0 Aab | 172.8 Aa | 154.0 Abc | 161.3 Aab | 165.7 ab | 174.0 a | 1.659 | <0.001 | <0.001 | 0.964 |
| 45 | 124.8 ABd | 159.5 a | 142.4 d | 148.1 cd | 160.2 ABbc | 167.2 ABab | 142.4 ABd | 154.6 ABbcd | 165.7 ab | 173.7 a | ||||
| 60 | 115.1 Bb | 153.2 a | 132.5 c | 147.7 bc | 153.9 Bab | 158.2 Bab | 136.5 Bc | 147.7 Bbc | 154.1 ab | 168.2 a | ||||
| WSC (g/kg DM) | ||||||||||||||
| 30 | 11.1 bc | 8.2 c | 9.7 bc | 10.9 abc | 11.2 abc | 12.7 a | 09.1 bc | 9.3 bc | 11.4 ab | 11.5 ab | 0.263 | 0.258 | <0.001 | 0.850 |
| 45 | 10.3 bcd | 7.9 d | 11.2 ab | 11.9 ab | 13.0 a | 13.6 a | 10.4 ab | 10.9 ab | 11.0 ab | 13.4 a | ||||
| 60 | 9.7 abcd | 5.0 f | 10.0 a | 10.1 a | 12.2 a | 13.8 a | 10.3 a | 10.9 a | 12.8 a | 13.4 a | ||||
| NDF (g/kg DM) | ||||||||||||||
| 30 | 638.1 Ba | 613.5 Abc | 608.6 Bcde | 602.9 Bcde | 597.4 Be | 579.8 Af | 629.5 Bab | 616.6 Bbc | 606.7 Bde | 601.1 Bde | 0.442 | <0.001 | <0.001 | <0.001 |
| 45 | 652.5 Ab | 619.7 Ade | 657.6 Aa | 642.2 Abc | 629.5 Acd | 593.7 Ae | 641.7 Abc | 633.2 Ac | 634.9 Ac | 571.6 Ae | ||||
| 60 | 563.8 Cc | 522.6 Be | 577.5 Cc | 572.3 Cb | 540.5 Cd | 538.0 Bd | 595.0 Ca | 583.5 Cb | 525.8 Ce | 525.7 Ce | ||||
| ADF (g/kg DM) | ||||||||||||||
| 30 | 439.1 Bbc | 405.8 Bf | 451.8 Ab | 433.1 Bc | 420.0 Bd | 415.6 Ade | 464.5 Aa | 469.0 Aa | 397.2 Cf | 404.0 Bef | 2.880 | <0.001 | <0.001 | <0.001 |
| 45 | 450.6 Ab | 435.3 Ac | 465.5 Aa | 453.9 Aab | 449.2 Ab | 416.8 Ad | 463.6 Aa | 443.8 Bbc | 444.2 Abc | 416.3 Ad | ||||
| 60 | 426.6 Ca | 391.0 Bd | 394.4 Bc | 394.0 Bc | 387.3 Ccd | 376.5 Be | 416.1 Bb | 414.1 Cb | 411.1 Bb | 383.2 Cde | ||||
| Ash (g/kg DM) | ||||||||||||||
| 30 | 57.7 | 56.6 A | 54.4 | 58.8 A | 58.1 | 59.7 A | 57.8 AB | 58.3 A | 60.0 A | 61.6 A | 0.476 | <0.001 | 0.104 | <0.001 |
| 45 | 61.2 a | 57.7 Ab | 54.0 bc | 54.1 ABbc | 54.1 bc | 49.7 Bc | 60.1 Aa | 53.7 Bbc | 51.8 Cc | 49.8 Cc | ||||
| 60 | 54.4 a | 48.4 Bbcd | 52.6 d | 52.6 Bd | 54.6 bcd | 55.6 ABbc | 53.7 Bbcd | 53.1 Bcd | 56.2 Bab | 58.4 Ba | ||||
| Item and Ensiling Days | Treatments | SEM | p | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | L | M1 | M2 | M3 | M4 | C1 | C2 | C3 | C4 | D | T | D × T | ||
| Lactic acid (% DM) | ||||||||||||||
| 30 | 0.52 f | 1.44 bc | 1.53 Aabc | 0.99 de | 1.70 Aab | 1.26 Acd | 0.77 ef | 1.72 ab | 1.50 abc | 1.85 Aa | 0.048 | 0.006 | <0.001 | 0.017 |
| 45 | 0.45 c | 1.30 ab | 0.89 Cbc | 1.37 ab | 1.29 Bab | 1.73 ABa | 0.95 bc | 1.18 ab | 1.21 ab | 1.21 Bab | ||||
| 60 | 0.44 c | 1.50 ab | 1.28 Bb | 1.63 ab | 1.56 ABab | 2.01 Ba | 1.26 bb | 1.51 ab | 1.41 b | 1.37 ABb | ||||
| Acetic acid (% DM) | ||||||||||||||
| 30 | 1.83 a | 1.37 ab | 1.69 ab | 1.55 ab | 1.30 Aabc | 1.24 Abc | 1.78 ab | 1.54 ab | 0.82 Ac | 1.65 Aab | 0.057 | <0.001 | <0.001 | 0.334 |
| 45 | 1.63 a | 0.92 bc | 1.58 a | 1.47 a | 1.38 Aab | 1.50 Aa | 1.47 a | 1.41 ab | 0.54 Ac | 1.34 Aab | ||||
| 60 | 1.91 a | 1.09 ab | 1.44 ab | 0.92 ab | 0.41 Bb | 0.49 Bb | 1.52 ab | 1.02 ab | 0.59 ABb | 0.61 Bb | ||||
| Propionic acid (% DM) | ||||||||||||||
| 30 | 0.24 Ba | 0.11 ABc | 0.18 b | 0.07 Acde | 0.06 Bde | 0.09 cd | 0.09 ABcd | 0.10 Bcd | 0.04 e | 0.04 Be | 0.001 | <0.001 | <0.001 | ,0.001 |
| 45 | 0.16 Ca | 0.05 Bb | 0.13 a | 0.04 ABb | 0.04 ABb | 0.02 b | 0.03 Bb | 0.15 Aa | 0.03 b | 0.04 Bb | ||||
| 60 | 0.38 Aa | 0.17 Ab | 0.13 bcd | 0.02 Bd | 0.02 Bd | 0.04 cd | 0.15 Abc | 0.19 Ab | 0.03 cd | 0.22 Ab | ||||
| pH | ||||||||||||||
| 30 | 5.16 a | 4.50 c | 4.84 b | 4.63 c | 4.53 c | 4.36 ABd | 4.89 b | 4.61 c | 4.51 c | 4.35 ABd | 0.028 | <0.001 | <0.001 | 0.988 |
| 45 | 5.25 a | 4.54 cde | 4.87 b | 4.66 cd | 4.50 def | 4.38 Afg | 4.84 b | 4.68 c | 4.51 def | 4.37 Ag | ||||
| 60 | 5.15 a | 4.45 c | 4.70 b | 4.57 c | 4.44 c | 4.24 Bd | 4.75 b | 4.56 c | 4.47 c | 4.29 Bd | ||||
| NH3-N:TN | ||||||||||||||
| 30 | 4.96 Aab | 4.86 Ab | 5.07 Aa | 2.82 Ae | 3.00 Ad | 1.80 Ah | 3.57 Ac | 3.05 Ad | 2.42 Af | 2.07 Ag | 0.135 | <0.001 | <0.001 | <0.001 |
| 45 | 6.16 Bb | 5.32 Bc | 6.63 Ba | 4.22 Bd | 4.00 Be | 3.31 Bg | 3.58 Af | 4.07 Bde | 3.13 Bh | 2.38 ABi | ||||
| 60 | 6.20 Ba | 4.58 Abc | 5.83 Ca | 3.96 Bcd | 4.47 Bbc | 2.97 Cef | 5.01 Bb | 4.17 Bc | 3.38 Bde | 2.60 Bf | ||||
| Item and Ensiling Days | Treatments | SEM | p | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CK | L | ML1 | ML2 | ML3 | ML4 | CL1 | CL2 | CL3 | CL4 | D | T | D × T | ||
| Lactic acid (% DM) | ||||||||||||||
| 30 | 0.52 d | 1.44 c | 1.57 Abc | 1.62 bc | 1.61 bc | 2.66 a | 2.08 Aabc | 2.19 Aab | 1.99 bc | 1.97 Abc | 0.060 | 0.003 | <0.001 | 0.199 |
| 45 | 0.45 d | 1.30 bc | 0.93 Bcd | 1.56 ab | 1.65 ab | 1.91 a | 1.25 Bbc | 1.38 Babc | 1.66 ab | 1.45 Babc | ||||
| 60 | 0.44 b | 1.50 a | 1.69 Aa | 1.99 a | 1.80 a | 1.97 a | 1.34 ABa | 1.65 ABa | 1.69 a | 1.76 ABa | ||||
| Acetic acid (% DM) | ||||||||||||||
| 30 | 1.83 a | 1.37 b | 1.46 ab | 1.35 b | 0.65 Bd | 0.84 Bd | 1.28 bc | 1.16 Abc | 0.54 d | 0.82 Bcd | 0.051 | <0.001 | <0.001 | <0.001 |
| 45 | 1.63 a | 0.92 c | 1.48 ab | 1.26 abc | 1.16 Abc | 1.21 Aabc | 1.32 abc | 1.19 Abc | 0.49 d | 1.17 Abc | ||||
| 60 | 1.91 a | 1.09 bc | 0.92 bcd | 0.83 bcd | 0.35 Bcd | 0.31 Cd | 1.33 ab | 0.83 Bbcd | 0.45 cd | 0.48 Bcd | ||||
| Propionic acid (% DM) | ||||||||||||||
| 30 | 0.24 Ba | 0.11 ABb | 0.05 Bc | 0.05 ABc | 0.04 c | 0.05 Bc | 0.04 c | 0.04 c | 0.03 Bc | 0.09 ABb | 0.009 | <0.001 | <0.001 | <0.001 |
| 45 | 0.16 Ca | 0.05 Bbc | 0.04 Bbc | 0.03 Bbc | 0.02 c | 0.02 Cc | 0.03 bc | 0.04 bc | 0.06 Ab | 0.06 Bbc | ||||
| 60 | 0.38 Aa | 0.17 Ab | 0.12 Abcd | 0.05 Ade | 0.03 e | 0.18 Ab | 0.08 cde | 0.02 e | 0.04 ABe | 0.14 Abc | ||||
| pH | ||||||||||||||
| 30 | 5.16 a | 4.50 c | 4.73 ABb | 4.52 c | 4.42 ABc | 4.26 ABd | 4.66 b | 4.48 c | 4.45 c | 4.28 ABd | 0.028 | <0.001 | <0.001 | 0.999 |
| 45 | 5.25 a | 4.54 c | 4.76 Ab | 4.55 c | 4.49 Ac | 4.34 Ad | 4.70 b | 4.54 c | 4.50 c | 4.35 Ad | ||||
| 60 | 5.15 a | 4.45 bc | 4.58 Bb | 4.47 bc | 4.33 Bcd | 4.19 Bd | 4.56 b | 4.40 c | 4.39 c | 4.22 Bd | ||||
| NH3-N:TN | ||||||||||||||
| 30 | 4.96 Aa | 4.86 Aa | 3.92 Ab | 3.31 d | 2.24 Ae | 2.13 e | 3.64 Ac | 3.38 cd | 3.19 d | 1.97 Ae | 0.131 | <0.001 | <0.001 | <0.001 |
| 45 | 6.16 Ba | 5.32 Bc | 5.84 Bb | 3.34 e | 2.97 Bf | 2.41 g | 4.57 Bd | 3.37 e | 3.31 e | 2.06 ABh | ||||
| 60 | 6.20 Ba | 4.58 Ab | 4.66 Cb | 3.57 c | 2.76 Cde | 2.16 e | 4.44 Bb | 3.64 c | 3.04 cd | 2.26 Be | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Q.; Shi, W.; Chen, S.; Degen, A.A.; Qi, Y.; Yang, F.; Zhou, J. Addition of Organic Acids and Lactobacillus acidophilus to the Leguminous Forage Chamaecrista rotundifolia Improved the Quality and Decreased Harmful Bacteria of the Silage. Animals 2022, 12, 2260. https://doi.org/10.3390/ani12172260
Feng Q, Shi W, Chen S, Degen AA, Qi Y, Yang F, Zhou J. Addition of Organic Acids and Lactobacillus acidophilus to the Leguminous Forage Chamaecrista rotundifolia Improved the Quality and Decreased Harmful Bacteria of the Silage. Animals. 2022; 12(17):2260. https://doi.org/10.3390/ani12172260
Chicago/Turabian StyleFeng, Qixian, Wenjiao Shi, Siqi Chen, Abraham Allan Degen, Yue Qi, Fulin Yang, and Jing Zhou. 2022. "Addition of Organic Acids and Lactobacillus acidophilus to the Leguminous Forage Chamaecrista rotundifolia Improved the Quality and Decreased Harmful Bacteria of the Silage" Animals 12, no. 17: 2260. https://doi.org/10.3390/ani12172260
APA StyleFeng, Q., Shi, W., Chen, S., Degen, A. A., Qi, Y., Yang, F., & Zhou, J. (2022). Addition of Organic Acids and Lactobacillus acidophilus to the Leguminous Forage Chamaecrista rotundifolia Improved the Quality and Decreased Harmful Bacteria of the Silage. Animals, 12(17), 2260. https://doi.org/10.3390/ani12172260