
Inflammatory Changes and Composition of Collagen during Cervical Ripening in Cows

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
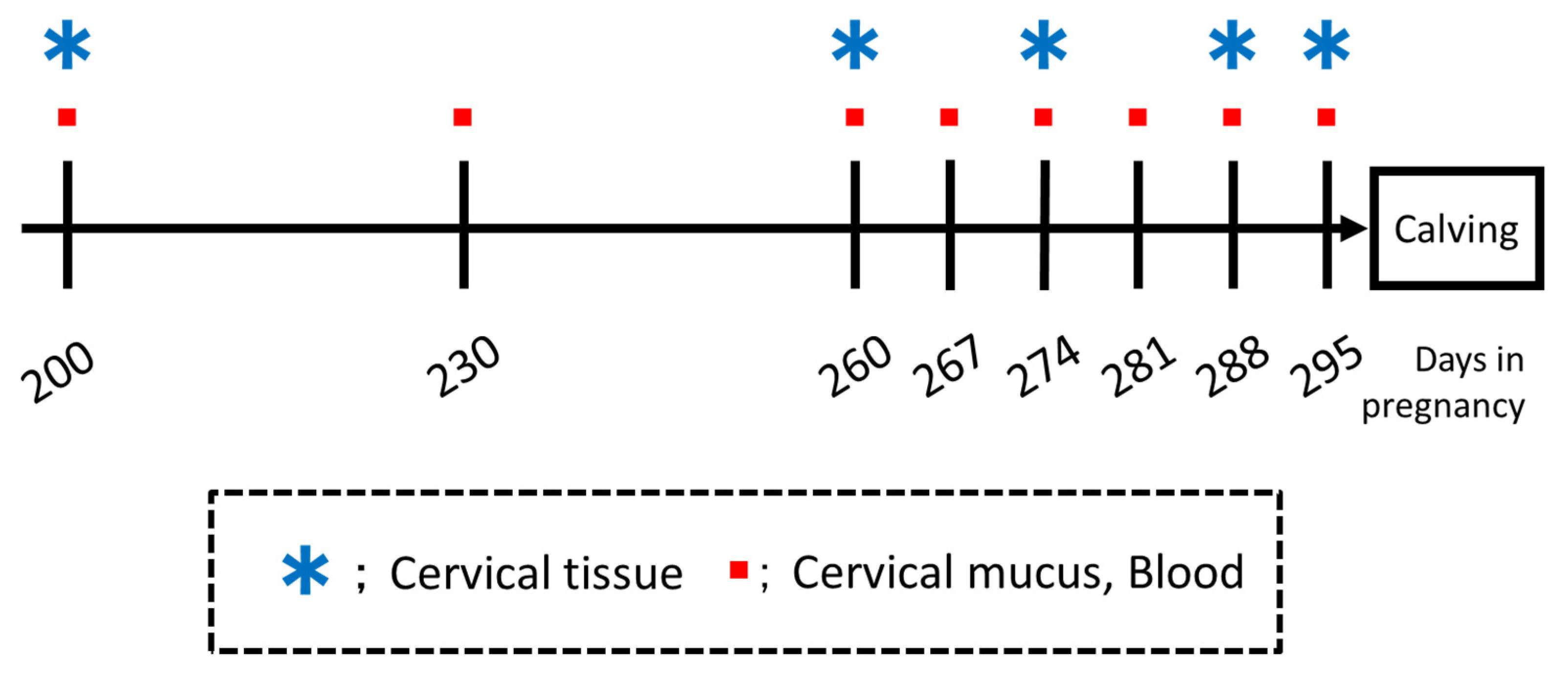
2.2. Study Design
2.3. Cervical Mucus Collection
2.3.1. Percentage of Polymorphonuclear Neutrophils (PMN%) of Cervical Mucus
2.3.2. Protein Analysis of Cervical Mucus
Cervical Mucus Processing and Protein Quantification
Determination of IL-8 Concentrations in Cervical Mucus
2.3.3. Collagen Analysis of Cervical Tissue
Sample Collections and Processing
Image Analyses
2.4. Leukocyte Counts in Peripheral Blood
2.5. Statistical Analysis
3. Results
3.1. Cervical Mucus
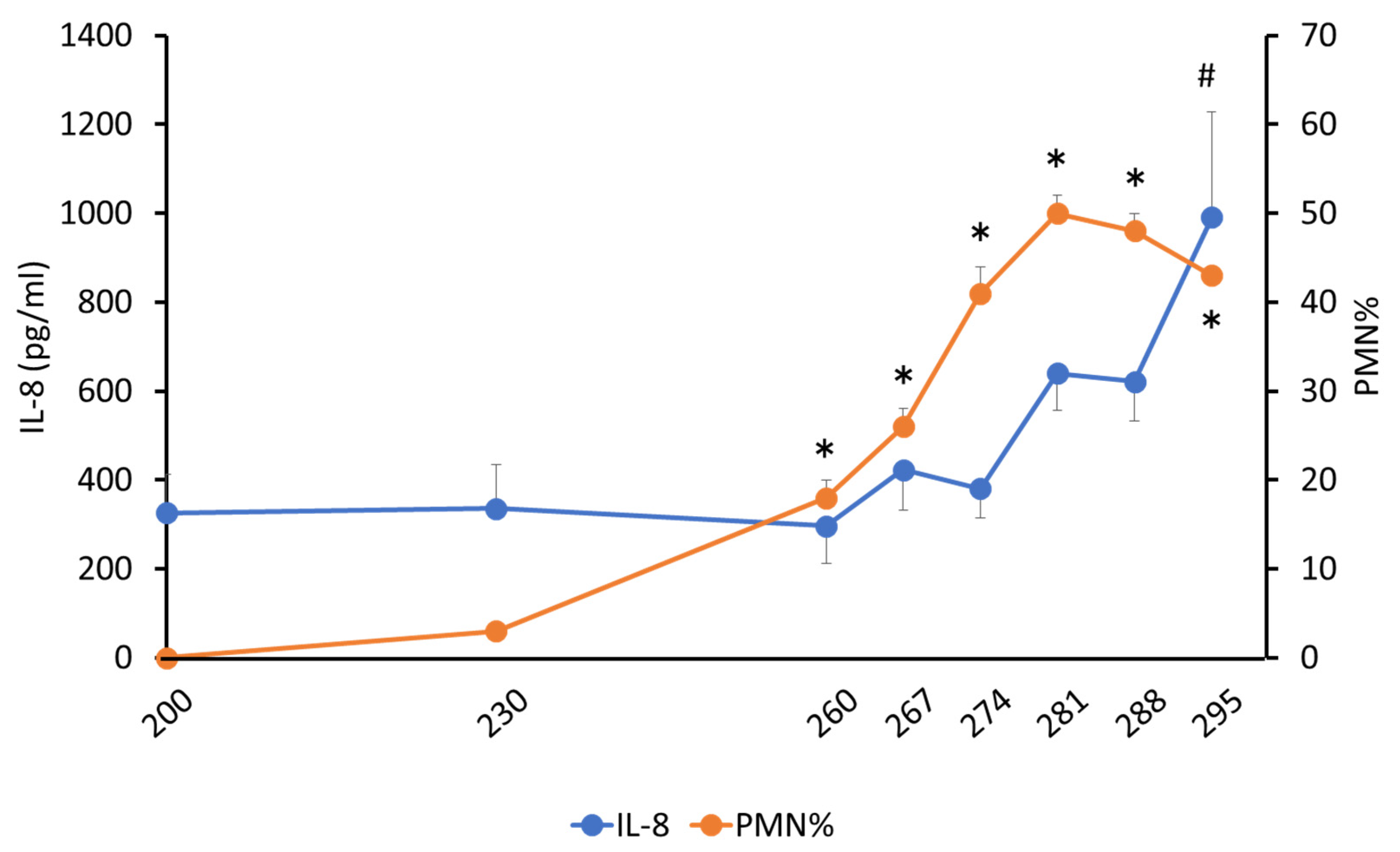
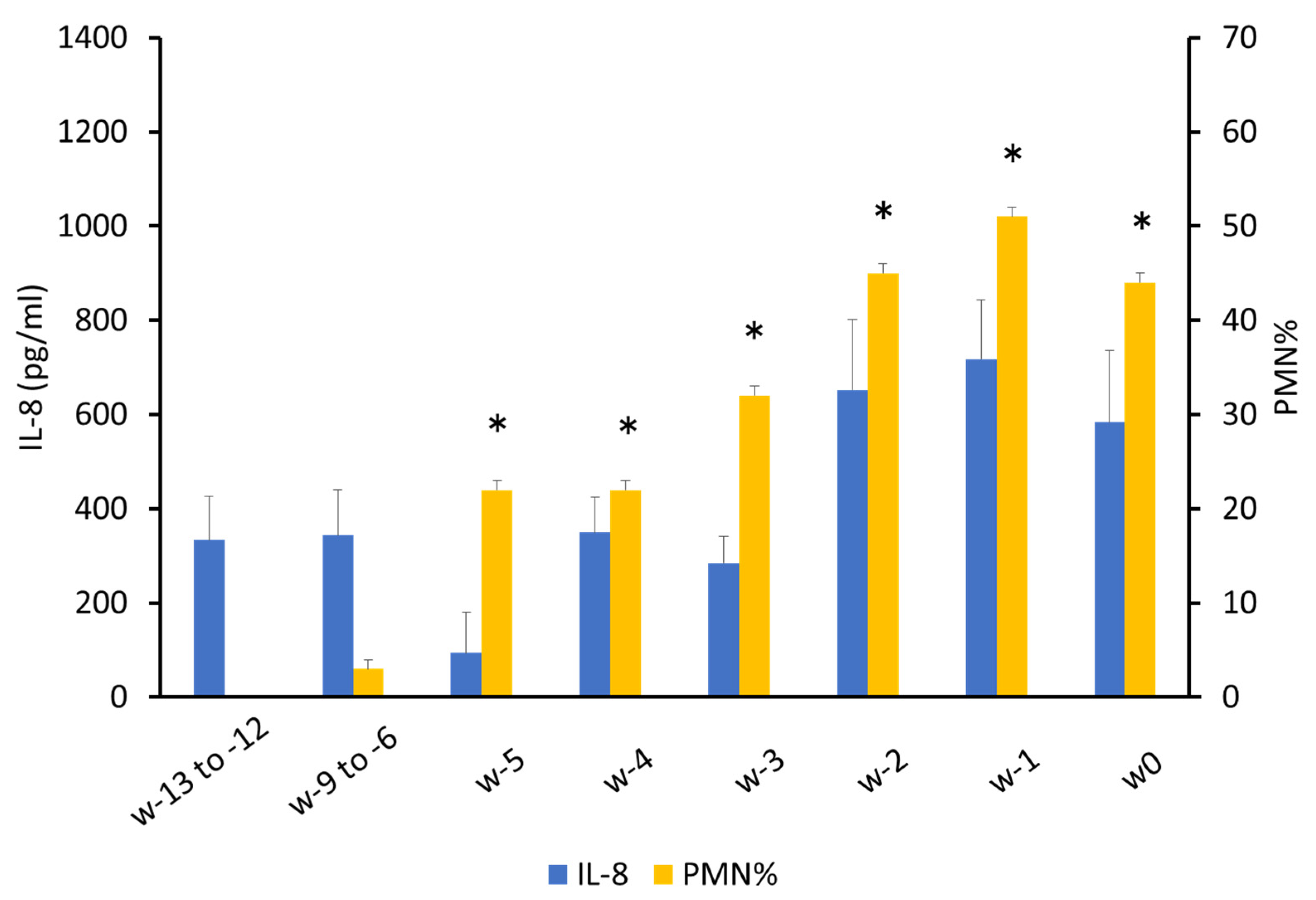
3.1.1. PMN%
3.1.2. Determination of Total Protein and Cytokine Concentrations
3.2. Changes in Collagen Composition in Cervical Canal Tissue
3.3. Leukocyte Count in Peripheral Blood
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uematsu, M.; Sasaki, Y.; Kitahara, G.; Sameshima, H.; Osawa, T. Risk factors for stillbirth and dystocia in Japanese Black cattle. Vet. J. 2013, 198, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Nix, J.M.; Spitzer, J.C.; Grimes, L.W.; Burns, G.L.; Plyler, B.B. A retrospective analysis of factors contributing to calf mortality and dystocia in beef cattle. Theriogenology 1998, 49, 1515–1523. [Google Scholar] [CrossRef]
- Danforth, D.N. The fibrous nature of the human cervix, and its relation to the isthmic segment in gravid and nongravid uteri. Am. J. Obstet. Gynecol. 1947, 53, 541–560. [Google Scholar] [CrossRef]
- Yellon, S.M. Immunobiology of cervix ripening. Front. Immunol. 2020, 10, 3156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mee, J.F. Prevalence and risk factors for dystocia in dairy cattle: A review. Vet. J. 2008, 176, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Ito, A.; Mori, Y. Dynamics of collagen and proteoglycans in uterine cervix: With special reference to cervical ripening and dilation. Connect. Tissue 1987, 19, 1–11. [Google Scholar]
- Sheetal, S.K.; Choudhary, S.K.; Sengupta, D. Mineral deficiency predisposes occurrence of retention of placenta in crossbred. Vet. World 2014, 7, 1140–1143. [Google Scholar] [CrossRef] [Green Version]
- Myers, K.; Socrate, S.; Tzeranis, D.; House, M. Changes in the biochemical constituents and morphologic appearance of the human cervical stroma during pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2009, 144, S82–S89. [Google Scholar] [CrossRef] [Green Version]
- Granstrom, L.; Ekman, G.; Ulmsten, U.; Malmstrom, A. Changes in the connective tissue of corpus and cervix uteri during ripening and labour in term pregnancy. BJOG 1989, 96, 1198–1202. [Google Scholar] [CrossRef]
- Kelly, R.W. Inflammatory mediators and cervical ripening. J. Reprod. Immunol. 2002, 57, 217–224. [Google Scholar] [CrossRef]
- Kirby, M.A.; Heuerman, A.C.; Custer, M.; Dobyns, A.E.; Strilaeff, R.; Stutz, K.N.; Cooperrider, J.; Elsissy, J.G.; Yellon, S.M. Progesterone receptor–mediated actions regulate remodeling of the cervix in preparation for preterm parturition. Reprod. Sci. 2016, 23, 1473–1483. [Google Scholar] [CrossRef] [PubMed]
- Mackler, A.M.; Iezza, G.; Akin, M.R.; McMillan, P.; Yellon, S.M. Macrophage trafficking in the uterus and cervix precedes parturition in the mouse. Biol. Reprod. 1999, 61, 879–883. [Google Scholar] [CrossRef] [Green Version]
- Winkler, M. Role of cytokines and other inflammatory mediators. BJOG 2003, 110, 118–123. [Google Scholar] [CrossRef]
- Chwalisz, K.; Benson, M.; Scholz, P.; Daum, J.; Beier, H.M.; Hegele-Hartung, C. Pregnancy: Cervical ripening with the cytokines interleukin 8, interleukin 1β and tumour necrosis factor α in guinea-pigs. Hum. Reprod. 1994, 9, 2173–2181. [Google Scholar] [CrossRef] [PubMed]
- Sennström, M.K.; Brauner, A.; Lu, Y.; Granström, L.M.; Malmström, A.L.; Ekman, G.E. Interleukin-8 is a mediator of the final cervical ripening in humans. Eur. J. Obstet. Gynecol. Reprod. Biol. 1997, 74, 89–92. [Google Scholar] [CrossRef]
- Kanayama, N.; Terao, T. The relationship between granulocyte elastase-like activity of cervical mucus and cervical maturation. Acta Obstet. Gynecol. Scand. 1991, 70, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Iwahashi, M.; Muragaki, Y.; Ooshima, A.; Umesaki, N. Decreased type I collagen expression in human uterine cervix during pregnancy. J. Clin. Endocrinol. Metab. 2003, 88, 2231–2235. [Google Scholar] [CrossRef] [Green Version]
- Van Engelen, E.; De Groot, M.W.; Breeveld-Dwarkasing, V.N.A.; Everts, M.E.; Van Der Weyden, G.C.; Taverne, M.A.M.; Rutten, V.P.M.G. Cervical ripening and parturition in cows are driven by a cascade of pro-inflammatory cytokines. Reprod. Domest. Anim. 2009, 44, 834–841. [Google Scholar] [CrossRef]
- Ahmadi, M.R.; Kafi, M.; Ghodrat, M. Crystallization and the number of neutrophils increase in the cervical mucus as parturition approaches in dairy cows. Comp. Clin. Pathol. 2005, 14, 72–75. [Google Scholar] [CrossRef]
- Bishop, E.H. Pelvic Scoring for Elective Induction. Obstet. Gynecol. 1964, 24, 266–268. [Google Scholar]
- Chanrot, M.; Blomqvist, G.; Guo, Y.; Ullman, K.; Juremalm, M.; Bage, R.; Donofrio, G.; Valarcher, J.F.; Humblot, P. Bovine herpes virus type 4 alters TNF-α and IL-8 profiles and impairs the survival of bovine endometrial epithelial cells. Reprod. Biol. 2017, 17, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Breeveld-Dwarkasing, V.N.A.; Te Koppele, J.M.; Bank, R.A.; Van Der Weijden, G.C.; Taverne, M.A.M.; van Dissel-Emiliani, F.M.F. Changes in water content, collagen degradation, collagen content, and concentration in repeated biopsies of the cervix of pregnant cows. Biol. Reprod. 2003, 69, 1608–1614. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, L.C.U.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Rich, L.; Whittaker, P. Collagen and picrosirius red staining: A polarized light assessment of fibrillar hue and spatial distribution. J. Morphol. Sci. 2005, 22, 97–104. [Google Scholar]
- Short, C.S.; Quinlan, R.; Bennett, P.; Shattock, R.J.; Taylor, G.P. Optimising the collection of female genital tract fluid for cytokine analysis in pregnant women. J. Immunol. Methods 2018, 458, 15–20. [Google Scholar] [CrossRef]
- Mi, Y.; Wang, W.; Lu, J.; Zhang, C.; Wang, Y.; Ying, H.; Sun, K. Proteasome-mediated degradation of collagen III by cortisol in amnion fibroblasts. J. Mol. Endocrinol. 2018, 60, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Read, C.P.; Word, R.A.; Ruscheinsky, M.A.; Timmons, B.C.; Mahendroo, M.S. Cervical remodeling during pregnancy and parturition: Molecular characterization of the softening phase in mice. Reproduction 2007, 134, 327–340. [Google Scholar] [CrossRef] [Green Version]
- Akgul, Y.; Holt, R.; Mummert, M.; Word, A.; Mahendroo, M. Dynamic changes in cervical glycosaminoglycan composition during normal pregnancy and preterm birth. Endocrinology 2012, 153, 3493–3503. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.R.; Cleveland, E.M.; Deeken, C.R.; Huitron, S.S.; Aluka, K.J.; Davis, K.G. Type I/type III collagen ratio associated with diverticulitis of the colon in young patients. J. Surg. Res. 2017, 207, 229–234. [Google Scholar] [CrossRef]
- Vink, J.; Mourad, M. The pathophysiology of human premature cervical remodeling resulting in spontaneous preterm birth: Where are we now? Semin. Perinatol. 2017, 41, 427–437. [Google Scholar] [CrossRef]
- Shukla, V.; Barnhouse, V.; Ackerman, W.E.; Summerfield, T.L.; Powell, H.M.; Leight, J.L.; Kniss, D.A.; Ghadiali, S.N. Cellular mechanics of primary human cervical fibroblasts: Influence of progesterone and a pro-inflammatory cytokine. Ann. Biomed. Eng. 2018, 46, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Wang, H.; Ding, Y.; Li, J.; Li, J. Changes in the blood routine, biochemical indexes and the pro-inflammatory cytokine expressions of peripheral leukocytes in postpartum dairy cows with metritis. BMC Vet. Res. 2019, 15, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateus, L.; Da Costa, L.L.; Carvalho, H.; Serra, P.; Robalo Silva, J. Blood and intrauterine leukocyte profile and function in dairy cows that spontaneously recovered from postpartum endometritis. Reprod. Domest. Anim. 2002, 37, 176–180. [Google Scholar] [CrossRef]
- Preisler, M.T.; Weber, P.S.; Tempelman, R.J.; Erskine, R.J.; Hunt, H.; Burton, J.L. Glucocorticoid receptor down-regulation in neutrophils of periparturient cows. Am. J. Vet. Res. 2000, 61, 14–19. [Google Scholar] [CrossRef]
- Monsanto, S.P.; Daher, S.; Ono, E.; Pendeloski, K.P.T.; Trainá, É.; Mattar, R.; Tayade, C. Cervical cerclage placement decreases local levels of proinflammatory cytokines in patients with cervical insufficiency. Am. J. Obstet. Gynecol. 2017, 217, e1–e455. [Google Scholar] [CrossRef] [Green Version]
- Ménard, L. The use of clenbuterol in large animal obstetrics: Manual correction of bovine dystocias. Can. Vet. J. 1994, 35, 289–292. [Google Scholar] [PubMed]
- Johanson, J.M.; Berger, P.J. Birth weight as a predictor of calving ease and perinatal mortality in Holstein cattle. J. Dairy Sci. 2003, 86, 3745–3755. [Google Scholar] [CrossRef] [Green Version]
- Kodama, S. Prolonged gestation of Japanese black calves in Miyazaki prefecture, Japan. J. Jpn. Vet. Med. Assoc. 2005, 58, 395–397. [Google Scholar] [CrossRef] [Green Version]
- Namihira, T.; Takahashi, K.; Nakamura, I.; Kanemine, H.; Hossain, M.A.; Tamaki, M. Transition of gestation length of Japanese black breeding cows in Okinawa. J. Warm Soc. Anim. Sci. Jpn. 2011, 54, 189–194. (In Japanese) [Google Scholar]
- Tenore, J.L. Methods for cervical ripening and induction of labor. Am. Fam. Phys. 2003, 67, 2123–2128. [Google Scholar]
- Malone, L.; Opazo, J.C.; Ryan, P.L. Hoffmann FG. Progressive erosion of the Relaxin1 gene in bovids. Gen. Comp. Endocrinol. 2017, 252, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Jelinic, M.; Marshall, S.A.; Stewart, D.; Unemori, E.; Parry, L.J.; Leo, C.H. Peptide hormone relaxin: From bench to bedside. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R753–R760. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.H.; Musah, A.I.; Cho, S.J.; Schwabe, C.; Anderson, L.L. Continuous infusion of relaxin on periparturient progesterone, oxytocin and relaxin plasma concentrations and time of parturition in beef heifers. Anim. Reprod. Sci. 1997, 46, 15–25. [Google Scholar] [CrossRef]
- Feltovich, H. Labour and delivery: A clinician’s perspective on a biomechanics problem. Interface Focus 2019, 9, 20190032. [Google Scholar] [CrossRef] [Green Version]
- Sisti, G.; Paccosi, S.; Parenti, A.; Seravalli, V.; Linari, C.; Di Tommaso, M.; Witkin, S. Pro-inflammatory mediators in vaginal fluid and short cervical length in pregnancy. Bratisl. Lek. Listy. 2020, 121, 278–281. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.R.; Hong, S.S.; Kim, J.; Lee, K.Y. Neutrophil elastase in cervical fluid in women with short cervical length. Taiwan J. Obstet. Gynecol. 2018, 57, 407–410. [Google Scholar] [CrossRef]
- Owen, J.; Yost, N.; Berghella, V.; Thom, E.; Swain, M.; Dildy, G.A., III; Miodovnik, M.; Langer, O.; Sibai, B.; McNellis, D. Mid-trimester endovaginal sonography in women at high risk for spontaneous preterm birth. J. Am. Med. Assoc. 2001, 286, 1340–1348. [Google Scholar] [CrossRef] [Green Version]
- Nakai, A.; Taniuchi, Y.; Miyake, H.; Nakai, M.; Yokota, A.; Takeshita, T. Increased level of granulocyte elastase in cervical secretion is an independent predictive factor for preterm delivery. Gynecol. Obstet. Investig. 2005, 60, 87–91. [Google Scholar] [CrossRef]
- Jung, E.Y.; Park, J.W.; Ryu, A.; Lee, S.Y.; Cho, S.H.; Park, K.H. Prediction of impending preterm delivery based on sonographic cervical length and different cytokine levels in cervicovaginal fluid in preterm labor. J. Obstet. Gynaecol. Res. 2016, 42, 158–165. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamanokuchi, E.; Kitahara, G.; Kanemaru, K.; Hemmi, K.; Kobayashi, I.; Yamaguchi, R.; Osawa, T. Inflammatory Changes and Composition of Collagen during Cervical Ripening in Cows. Animals 2022, 12, 2646. https://doi.org/10.3390/ani12192646
Yamanokuchi E, Kitahara G, Kanemaru K, Hemmi K, Kobayashi I, Yamaguchi R, Osawa T. Inflammatory Changes and Composition of Collagen during Cervical Ripening in Cows. Animals. 2022; 12(19):2646. https://doi.org/10.3390/ani12192646
Chicago/Turabian StyleYamanokuchi, Eigo, Go Kitahara, Kazuyuki Kanemaru, Koichiro Hemmi, Ikuo Kobayashi, Ryoji Yamaguchi, and Takeshi Osawa. 2022. "Inflammatory Changes and Composition of Collagen during Cervical Ripening in Cows" Animals 12, no. 19: 2646. https://doi.org/10.3390/ani12192646
APA StyleYamanokuchi, E., Kitahara, G., Kanemaru, K., Hemmi, K., Kobayashi, I., Yamaguchi, R., & Osawa, T. (2022). Inflammatory Changes and Composition of Collagen during Cervical Ripening in Cows. Animals, 12(19), 2646. https://doi.org/10.3390/ani12192646