
Spatial Factors Outperform Local Environmental and Geo-Climatic Variables in Structuring Multiple Facets of Stream Macroinvertebrates’ β-Diversity


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling and Processing
2.3. Geo-Climatic Variables
2.4. Species Traits
2.5. Data Analysis
3. Results
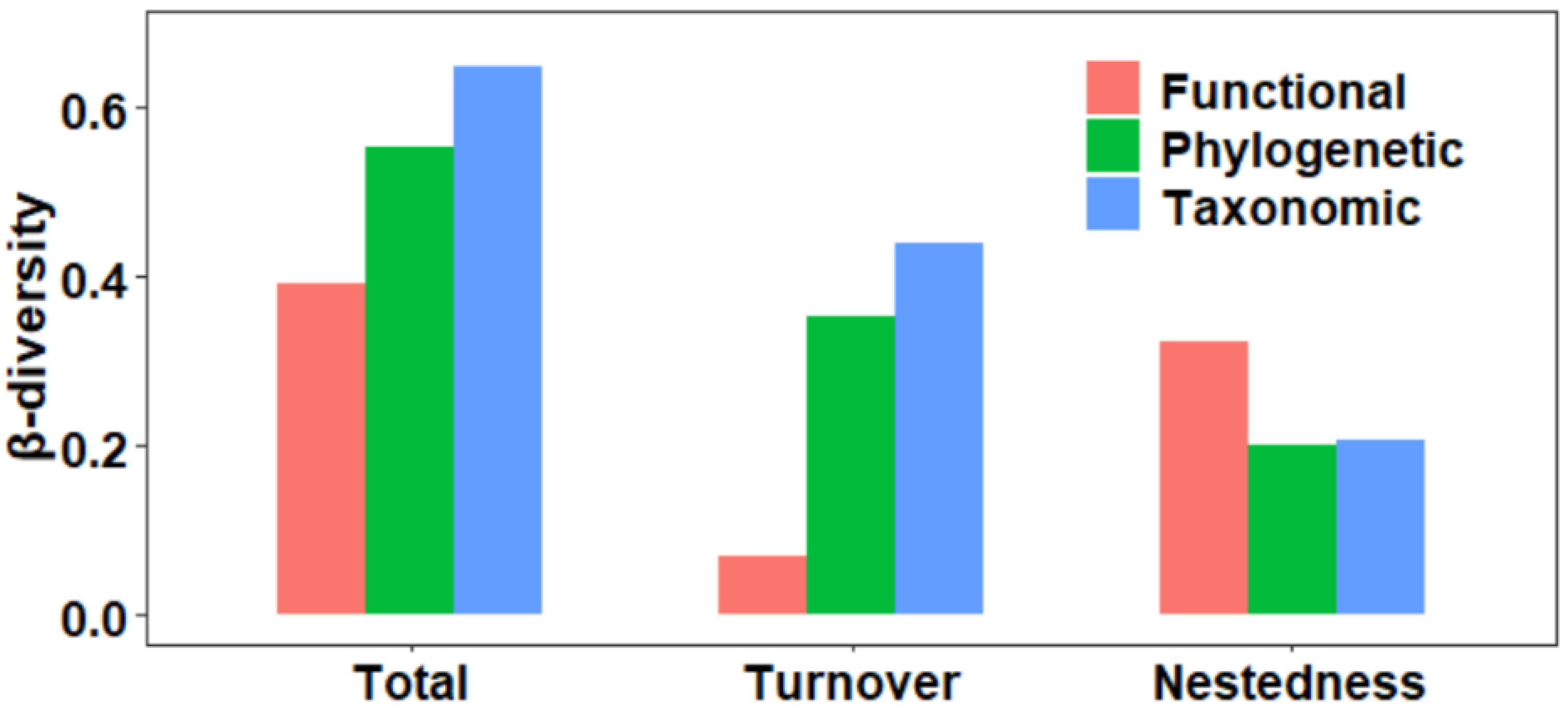
3.1. Taxonomic, Functional, and Phylogenetic β-Diversity Components
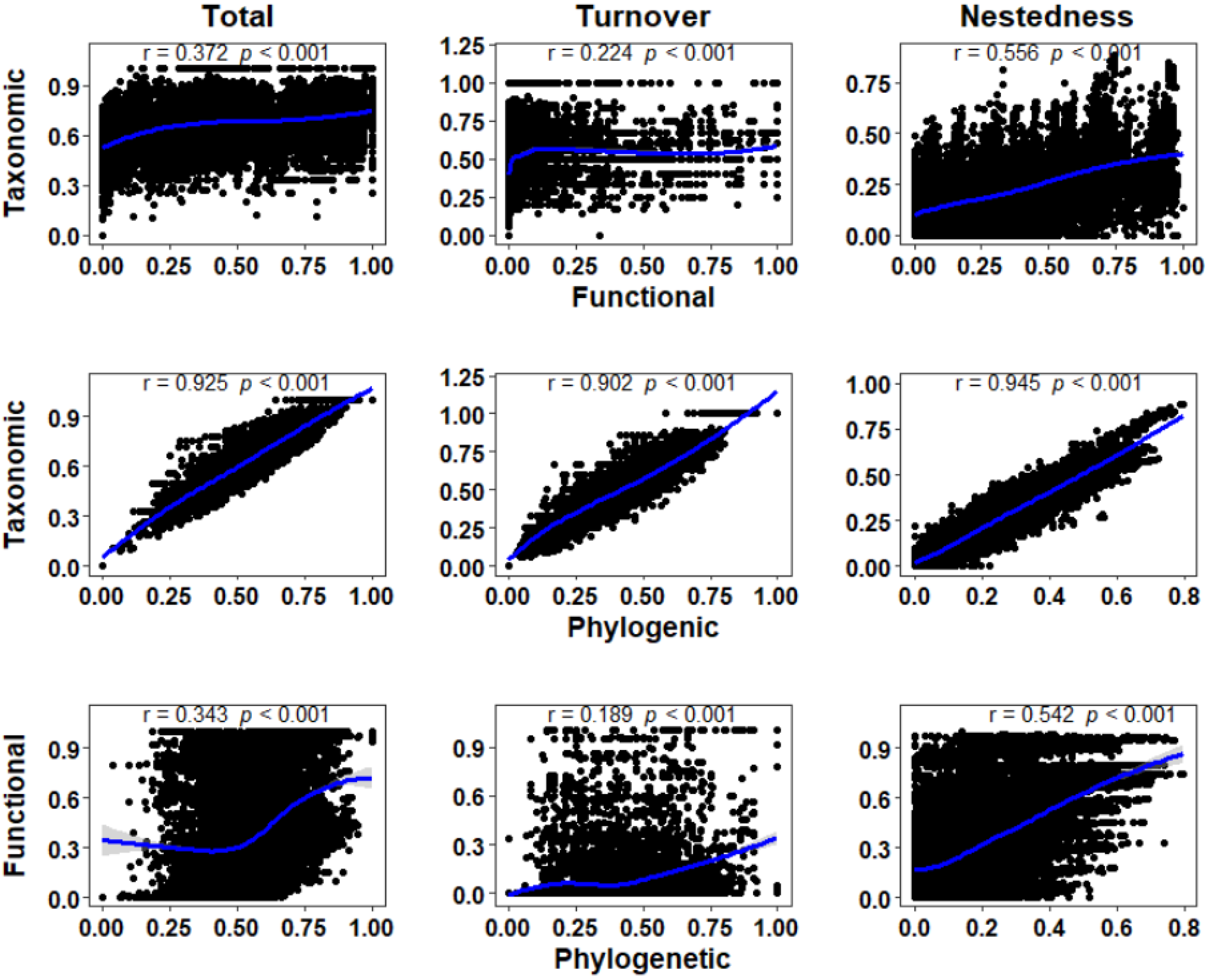
3.2. Correlation of Taxonomic, Functional, and Phylogenetic β-Diversity
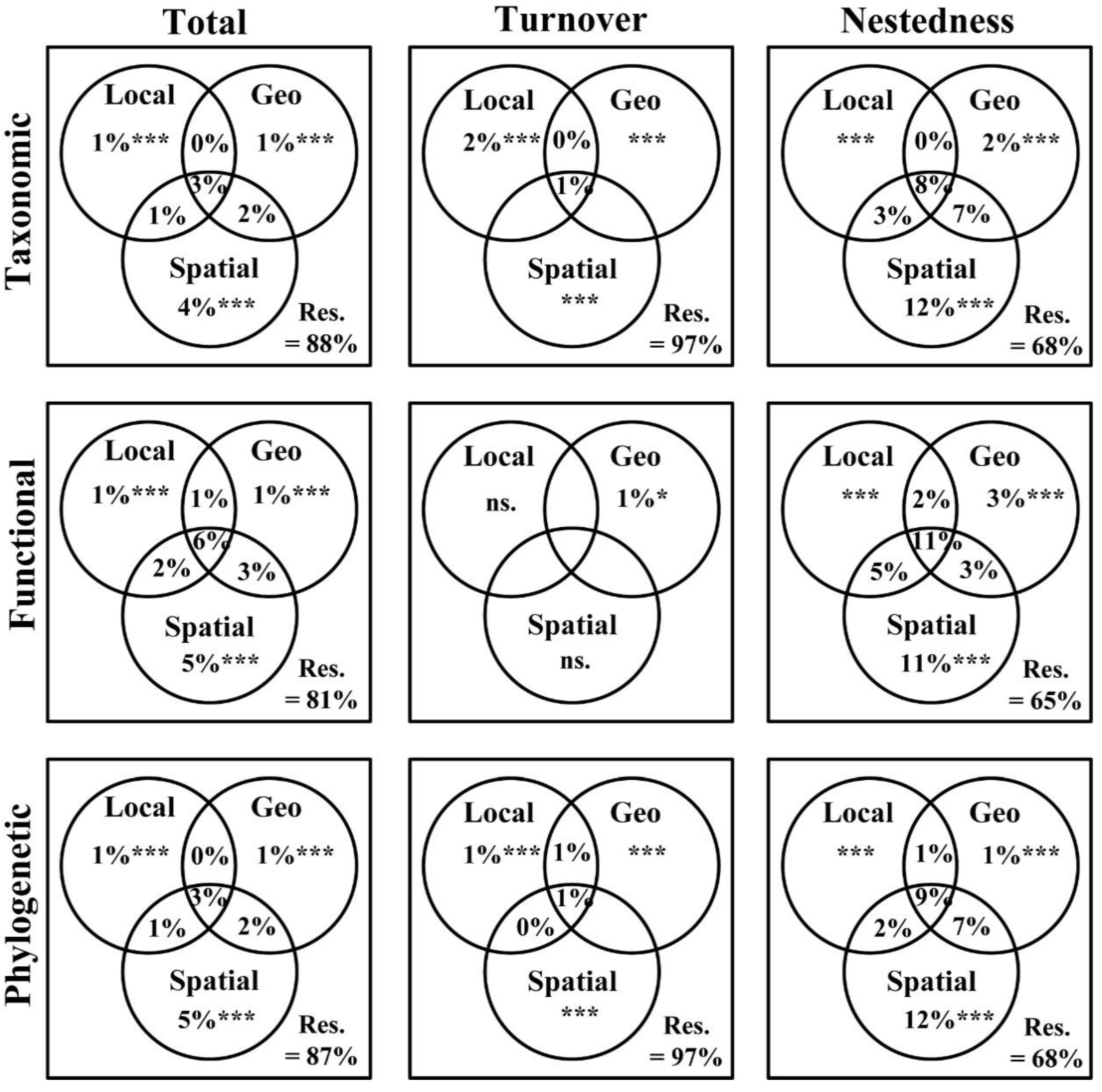
3.3. Main Drivers of Taxonomic, Functional, and Phylogenetic β-Diversity Components
4. Discussion
4.1. Contribution of Turnover and Nestedness
4.2. Relationships between Taxonomic, Functional, and Phylogenetic β-Diversity
4.3. Main Drivers of Taxonomic, Functional, and Phylogenetic β-Diversity
4.4. Management Implications and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Ellingsen, K.E.; Gray, J.S. Spatial patterns of benthic diversity: Is there a latitudinal gradient along the Norwegian continental shelf? J. Anim. Ecol. 2002, 71, 373–389. [Google Scholar] [CrossRef]
- Moss, W.E.; McDevitt-Galles, T.; Calhoun, D.M.; Johnson, P.T.J. Tracking the assembly of nested parasite communities: Using beta-diversity to understand variation in parasite richness and composition over time and scale. J. Anim. Ecol. 2020, 89, 1532–1542. [Google Scholar] [CrossRef]
- Svensson-Coelho, M.; Ricklefs, R.E. Host phylogeography and beta diversity in avian haemosporidian (Plasmodiidae) assemblages of the Lesser Antilles. J. Anim. Ecol. 2011, 80, 938–946. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Kozak, K.H.; Fine, P.V.A.; Kembel, S.W. The merging of community ecology and phylogenetic biology. Ecol. Lett. 2009, 12, 693–715. [Google Scholar] [CrossRef]
- Gianuca, A.T.; Declerck, S.A.J.; Cadotte, M.W.; Souffreau, C.; De Bie, T.; De Meester, L. Integrating trait and phylogenetic distances to assess scale-dependent community assembly processes. Ecography 2017, 40, 742–752. [Google Scholar] [CrossRef] [Green Version]
- Heino, J.; Tolonen, K.T. Ecological drivers of multiple facets of beta diversity in a lentic macroinvertebrate metacommunity. Limnol. Oceanogr. 2017, 62, 2431–2444. [Google Scholar] [CrossRef]
- Gianuca, A.T.; Engelen, J.; Brans, K.I.; Hanashiro, F.T.T.; Vanhamel, M.; van den Berg, E.M.; Souffreau, C.; De Meester, L. Taxonomic, functional and phylogenetic metacommunity ecology of cladoceran zooplankton along urbanization gradients. Ecography 2018, 41, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Pool, T.K.; Grenouillet, G.; Villeger, S. Species contribute differently to the taxonomic, functional, and phylogenetic alpha and beta diversity of freshwater fish communities. Divers. Distrib. 2014, 20, 1235–1244. [Google Scholar] [CrossRef]
- Wu, N.; Wang, Y.; Wang, Y.; Sun, X.; Faber, C.; Fohrer, N. Environment regimes play an important role in structuring trait- and taxonomy-based temporal beta diversity of riverine diatoms. J. Ecol. 2022, 110, 1442–1454. [Google Scholar] [CrossRef]
- Meynard, C.N.; Devictor, V.; Mouillot, D.; Thuiller, W.; Jiguet, F.; Mouquet, N. Beyond taxonomic diversity patterns: How do alpha, beta and gamma components of bird functional and phylogenetic diversity respond to environmental gradients across France? Glob. Ecol. Biogeogr. 2011, 20, 893–903. [Google Scholar] [CrossRef]
- Diaz, S.; Cabido, M. Vive la difference: Plant functional diversity matters to ecosystem processes. Trends Ecol. Evol. 2001, 16, 646–655. [Google Scholar] [CrossRef]
- Soininen, J.; Jamoneau, A.; Rosebery, J.; Passy, S.I. Global patterns and drivers of species and trait composition in diatoms. Glob. Ecol. Biogeogr. 2016, 25, 940–950. [Google Scholar] [CrossRef]
- Verberk, W.C.E.P.; van Noordwijk, C.G.E.; Hildrew, A.G. Delivering on a promise: Integrating species traits to transform descriptive community ecology into a predictive science. Freshw. Sci. 2013, 32, 531–547. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, M.; Alahuhta, J.; Heino, J.; Toivonen, H. No biotic homogenisation across decades but consistent effects of landscape position and pH on macrophyte communities in boreal lakes. Ecography 2020, 43, 294–305. [Google Scholar] [CrossRef]
- Si, X.; Baselga, A.; Leprieur, F.; Song, X.; Ding, P. Selective extinction drives taxonomic and functional alpha and beta diversities in island bird assemblages. J. Anim. Ecol. 2016, 85, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Zhou, S.; Zhang, M.; Peng, W.; Guo, K.; Qu, X.; He, F. Spatial and local environmental factors outweigh geo-climatic gradients in structuring taxonomically and trait-based b-diversity of benthic algae. J. Biogeogr. 2021, 48, 1842–1857. [Google Scholar] [CrossRef]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Veron, S.; Davies, T.J.; Cadotte, M.W.; Clergeau, P.; Pavoine, S. Predicting loss of evolutionary history: Where are we? Biol. Rev. 2017, 92, 271–291. [Google Scholar] [CrossRef]
- Morlon, H.; Schwilk, D.W.; Bryant, J.A.; Marquet, P.A.; Rebelo, A.G.; Tauss, C.; Bohannan, B.J.M.; Green, J.L. Spatial patterns of phylogenetic diversity. Ecol. Lett. 2011, 14, 141–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricklefs, R.E. Community Diversity: Relative Roles of Local and Regional Processes. Science 1987, 235, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Baselga, A.; Gomez-Rodriguez, C.; Lobo, J.M. Historical Legacies in World Amphibian Diversity Revealed by the Turnover and Nestedness Components of Beta Diversity. PLoS ONE 2012, 7, e32341. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Glob. Ecol. Biogeogr. 2014, 23, 1324–1334. [Google Scholar] [CrossRef]
- Cardoso, P.; Rigal, F.; Carvalho, J.C.; Fortelius, M.; Borges, P.A.V.; Podani, J.; Schmera, D. Partitioning taxon, phylogenetic and functional beta diversity into replacement and richness difference components. J. Biogeogr. 2014, 41, 749–761. [Google Scholar] [CrossRef] [Green Version]
- Devictor, V.; Mouillot, D.; Meynard, C.; Jiguet, F.; Thuiller, W.; Mouquet, N. Spatial mismatch and congruence between taxonomic, phylogenetic and functional diversity: The need for integrative conservation strategies in a changing world. Ecol. Lett. 2010, 13, 1030–1040. [Google Scholar] [CrossRef]
- Jiang, X.; Pan, B.; Jiang, W.; Hou, Y.; Yang, H.; Zhu, P.; Heino, J. The role of environmental conditions, climatic factors and spatial processes in driving multiple facets of stream macroinvertebrate beta diversity in a climatically heterogeneous mountain region. Ecol. Indic. 2021, 124, 107407. [Google Scholar] [CrossRef]
- Tapolczai, K.; Bouchez, A.; Stenger-Kovacs, C.; Padisak, J.; Rimet, F. Trait-based ecological classifications for benthic algae: Review and perspectives. Hydrobiologia 2016, 776, 1–17. [Google Scholar] [CrossRef]
- Qu, X.; Peng, W.; Liu, Y.; Zhang, M.; Ren, Z.; Wu, N.; Liu, X. Networks and ordination analyses reveal the stream community structures of fish, macroinvertebrate and benthic algae, and their responses to nutrient enrichment. Ecol. Indic. 2019, 101, 501–511. [Google Scholar] [CrossRef]
- Zhou, S.; Wu, N.; Zhang, M.; Peng, W.; He, F.; Guo, K.; Yan, S.; Zou, Y.; Qu, X. Local environmental, geo-climatic and spatial factors interact to drive community distributions and diversity patterns of stream benthic algae, macroinvertebrates and fishes in a large basin, Northeast China. Ecol. Indic. 2020, 117, 106673. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, R.; Kong, W.; Geng, S.; Bentsen, C.N.; Qu, X. Relationships between macroinvertebrate communities and land use types within different riparian widths in three headwater streams of Taizi River, China. J. Freshw. Ecol. 2013, 28, 307–328. [Google Scholar] [CrossRef]
- Li, Z.; Heino, J.; Chen, X.; Liu, Z.; Meng, X.; Jiang, X.; Ge, Y.; Chen, J.; Xie, Z. Understanding macroinvertebrate metacommunity organization using a nested study design across a mountainous river network. Ecol. Indic. 2021, 121, 107188. [Google Scholar] [CrossRef]
- Strahler, A.N. Quantitative analysis of watershed geomorphology. Eos Trans. Am. Geophys. Union 1957, 38, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Taft, B.; Koncelik, J.P. Methods for Assessing Habitat in Flowing Waters: Using the Qualitative Habitat Evaluation Index (QHEI); OHIO EPA Technical Bulletin EAS; State of Ohio Environmental: Chicago, IL, USA, 2006.
- Merritt, R.W.; Cummins, K.W.; Berg, M.B. Trophic Relationships of Macroinvertebrates. In Methods in Stream Ecology; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Morse, J.C.; Yang, L.; Tian, L. Aquatic Insects of China Useful for Monitoring Water Quality; Hohai University Press: Nanjing, China, 1994. [Google Scholar]
- Tuanmu, M.-N.; Jetz, W. A global 1-km consensus land-cover product for biodiversity and ecosystem modelling. Glob. Ecol. Biogeogr. 2014, 23, 1031–1045. [Google Scholar] [CrossRef]
- Amatulli, G.; Domisch, S.; Tuanmu, M.-N.; Parmentier, B.; Ranipeta, A.; Malczyk, J.; Jetz, W. A suite of global, cross-scale topographic variables for environmental and biodiversity modeling. Sci. Data 2018, 5, 180040. [Google Scholar] [CrossRef] [Green Version]
- Domisch, S.; Amatulli, G.; Jetz, W. Near-global freshwater-specific environmental variables for biodiversity analyses in 1 km resolution. Sci. Data 2015, 2, 150073. [Google Scholar] [CrossRef] [Green Version]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Addo-Bediako, A. Spatial distribution patterns of benthic macroinvertebrate functional feeding groups in two rivers of the olifants river system, South Africa. J. Freshw. Ecol. 2021, 36, 97–109. [Google Scholar] [CrossRef]
- Heino, J. Functional biodiversity of macroinvertebrate assemblages along major ecological gradients of boreal headwater streams. Freshw. Biol. 2005, 50, 1578–1587. [Google Scholar] [CrossRef]
- Merritt, R.; Cummins, K.; Berg, M.B.; Adler, P.H.; Allen, M.; Batzer, D.P.; Bland, R.; Buchwalter, D.; Burian, S.K.; Byers, G.W. An introduction to the aquatic insects of North America. Rev. Mex. Biodivers. 2010, 81, 593–595. [Google Scholar]
- Ramirez, A.; Gutierrez-Fonseca, P.E. Functional feeding groups of aquatic insect families in Latin America: A critical analysis and review of existing literature. Rev. Biol. Trop. 2014, 62, 155–167. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. vegan: Community Ecology Package, R Package Version 2.5-7; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Dray, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Clappe, S.; Guenard, G.; Jombart, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. adespatial: Multivariate Multiscale Spatial Analysis, R Package Version 0.3-8; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Baselga, A.; Orme, C.D.L. betapart: An R package for the study of beta diversity. Methods Ecol. Evol. 2012, 3, 808–812. [Google Scholar] [CrossRef]
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: Testing multispecies responses in multifactorial ecological experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- Naimi, B.; Hamm, N.A.S.; Groen, T.A.; Skidmore, A.K.; Toxopeus, A.G. Where is positional uncertainty a problem for species distribution modelling? Ecography 2014, 37, 191–203. [Google Scholar] [CrossRef]
- Perez Rocha, M.; Bini, L.M.; Domisch, S.; Tolonen, K.T.; Jyrkankallio-Mikkola, J.; Soininen, J.; Hjort, J.; Heino, J. Local environment and space drive multiple facets of stream macroinvertebrate beta diversity. J. Biogeogr. 2018, 45, 2744–2754. [Google Scholar] [CrossRef] [Green Version]
- Borcard, D.; Legendre, P.; Drapeau, P. Partialling out the spatial component of ecological variation. Ecology 1992, 73, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Delgado, E.O.; Winemiller, K.O.; Villa-Navarro, F.A. Local environmental factors influence beta-diversity patterns of tropical fish assemblages more than spatial factors. Ecology 2020, 101, e02940. [Google Scholar] [CrossRef]
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward Selection of Explanatory Variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; He, L.; Lu, Y.; Su, J.; Song, H.; Zeng, X.; Yu, Z. Distribution, source, and ecological risk assessment of polycyclic aromatic hydrocarbons (PAHs) in surface sediments from the Hun River, northeast China. Environ. Monit. Assess. 2015, 187, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wang, J.; Liu, Z.; Meng, X.; Heino, J.; Jiang, X.; Xiong, X.; Jiang, X.; Xie, Z. Different responses of taxonomic and functional structures of stream macroinvertebrate communities to local stressors and regional factors in a subtropical biodiversity hotspot. Sci. Total Environ. 2019, 655, 1288–1300. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tan, X. Macroinvertebrate community in relation to water quality and riparian land use in a substropical mountain stream, China. Environ. Sci. Pollut. Res. 2017, 24, 14682–14689. [Google Scholar] [CrossRef]
- Zhang, Y.; Peng, C.; Huang, S.; Wang, J.; Xiong, X.; Li, D. The relative role of spatial and environmental processes on seasonal variations of phytoplankton beta diversity along different anthropogenic disturbances of subtropical rivers in China. Environ. Sci. Pollut. Res. 2019, 26, 1422–1434. [Google Scholar] [CrossRef]
- Perez Rocha, M.; Bini, L.M.; Gronroos, M.; Hjort, J.; Lindholm, M.; Karjalainen, S.-M.; Tolonen, K.E.; Heino, J. Correlates of different facets and components of beta diversity in stream organisms. Oecologia 2019, 191, 919–929. [Google Scholar] [CrossRef] [Green Version]
- Villeger, S.; Grenouillet, G.; Brosse, S. Decomposing functional -diversity reveals that low functional -diversity is driven by low functional turnover in European fish assemblages. Glob. Ecol. Biogeogr. 2013, 22, 671–681. [Google Scholar] [CrossRef]
- Vilar, A.G.; van Dam, H.; van Loon, E.E.; Vonk, J.A.; van Der Geest, H.G.; Admiraal, W. Eutrophication decreases distance decay of similarity in diatom communities. Freshw. Biol. 2014, 59, 1522–1531. [Google Scholar] [CrossRef]
- Branco, C.C.Z.; Bispo, P.C.; Peres, C.K.; Tonetto, A.F.; Krupek, R.A.; Barfield, M.; Holt, R.D. Partitioning multiple facets of beta diversity in a tropical stream macroalgal metacommunity. J. Biogeogr. 2020, 47, 1765–1780. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Cavender-Bares, J.; Tilman, D.; Oakley, T.H. Using Phylogenetic, Functional and Trait Diversity to Understand Patterns of Plant Community Productivity. PLoS ONE 2009, 4, e5695. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; McPeek, M.A.; Donoghue, M.J. Phylogenies and community ecology. Annu. Rev. Ecol. Syst. 2002, 33, 475–505. [Google Scholar] [CrossRef] [Green Version]
- Heino, J.; Mykra, H.; Kotanen, J.; Muotka, T. Ecological filters and variability in stream macroinvertebrate communities: Do taxonomic and functional structure follow the same path? Ecography 2007, 30, 217–230. [Google Scholar] [CrossRef]
- Mykra, H.; Heino, J.; Muotka, T. Scale-related patterns in the spatial and environmental components of stream macroinvertebrate assemblage variation. Glob. Ecol. Biogeogr. 2007, 16, 149–159. [Google Scholar] [CrossRef]
- Poff, N.L. Landscape filters and species traits: Towards mechanistic understanding and prediction in stream ecology. J. N. Am. Benthol. Soc. 1997, 16, 391–409. [Google Scholar] [CrossRef]
- Heino, J. A macroecological perspective of diversity patterns in the freshwater realm. Freshw. Biol. 2011, 56, 1703–1722. [Google Scholar] [CrossRef]
- Cai, Y.; Xu, H.; Vilmi, A.; Tolonen, K.T.; Tang, X.; Qin, B.; Gong, Z.; Heino, J. Relative roles of spatial processes, natural factors and anthropogenic stressors in structuring a lake macroinvertebrate metacommunity. Sci. Total Environ. 2017, 601, 1702–1711. [Google Scholar] [CrossRef]
- Mouquet, N.; Loreau, M. Community patterns in source-sink metacommunities. Am. Nat. 2003, 162, 544–557. [Google Scholar] [CrossRef] [Green Version]
- Heino, J.; Melo, A.S.; Siqueira, T.; Soininen, J.; Valanko, S.; Bini, L.M. Metacommunity organisation, spatial extent and dispersal in aquatic systems: Patterns, processes and prospects. Freshw. Biol. 2015, 60, 845–869. [Google Scholar] [CrossRef]
- Sandin, L. Benthic macroinvertebrates in Swedish streams: Community structure, taxon richness, and environmental relations. Ecography 2003, 26, 269–282. [Google Scholar] [CrossRef]
- Hill, M.J.; Heino, J.; White, J.C.; Ryves, D.B.; Wood, P.J. Environmental factors are primary determinants of different facets of pond macroinvertebrate alpha and beta diversity in a human-modified landscape. Biol. Conserv. 2019, 237, 348–357. [Google Scholar] [CrossRef]
- Qu, X.; Zhang, H.; Zhang, M.; Liu, M.; Yu, Y.; Xie, Y.; Peng, W. Application of multiple biological indices for river health assessment in northeastern China. Ann. Limnol. Int. J. Limnol. 2016, 52, 75–89. [Google Scholar] [CrossRef]
- Qu, X.; Ren, Z.; Zhang, M.; Liu, X.; Peng, W. Sediment heavy metals and benthic diversities in Hun-Tai River, northeast of China. Environ. Sci. Pollut. Res. 2017, 24, 10662–10673. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, Z.; Heino, J.; Jiang, X.; Wang, J.; Tang, T.; Xie, Z. Discriminating the effects of local stressors from climatic factors and dispersal processes on multiple biodiversity dimensions of macroinvertebrate communities across subtropical drainage basins. Sci. Total Environ. 2020, 711, 134750. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cai, Q.; Jiang, W.; Qu, X. The response of benthic macroinvertebrate communities to climate change: Evidence from subtropical mountain streams in Central China. Int. Rev. Hydrobiol. 2012, 97, 200–214. [Google Scholar] [CrossRef]
- Karna, O.-M.; Gronroos, M.; Antikainen, H.; Hjort, J.; Ilmonen, J.; Paasivirta, L.; Heino, J. Inferring the effects of potential dispersal routes on the metacommunity structure of stream insects: As the crow flies, as the fish swims or as the fox runs? J. Anim. Ecol. 2015, 84, 1342–1353. [Google Scholar] [CrossRef] [Green Version]
- Morante-Filho, J.C.; Arroyo-Rodriguez, V.; Faria, D. Patterns and predictors of beta-diversity in the fragmented Brazilian Atlantic forest: A multiscale analysis of forest specialist and generalist birds. J. Anim. Ecol. 2016, 85, 240–250. [Google Scholar] [CrossRef] [Green Version]
- Wearn, O.R.; Carbone, C.; Rowcliffe, J.M.; Pfeifer, M.; Bernard, H.; Ewers, R.M. Land-use change alters the mechanisms assembling rainforest mammal communities in Borneo. J. Anim. Ecol. 2019, 88, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Xiong, D.; Wei, C.; Wang, X.; Lu, X.; Fang, S.; Li, Y.; Wang, X.; Liang, W.; Han, X.; Bezemer, T.M.; et al. Spatial patterns and ecological drivers of soil nematode beta-diversity in natural grasslands vary among vegetation types and trophic position. J. Anim. Ecol. 2021, 90, 1367–1378. [Google Scholar] [CrossRef]
- Beck, J.; Khen, C.V. Beta-diversity of geometrid moths from northern Borneo: Effects of habitat, time and space. J. Anim. Ecol. 2007, 76, 230–237. [Google Scholar] [CrossRef]
- Ellingsen, K.E.; Anderson, M.J.; Shackell, N.L.; Tveraa, T.; Yoccoz, N.G.; Frank, K.T. The role of a dominant predator in shaping biodiversity over space and time in a marine ecosystem. J. Anim. Ecol. 2015, 84, 1242–1252. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, M.; Peng, W.; Qu, X.; Zhang, Y.; Du, L.; Wu, N. Phylogenetic and functional diversity could be better indicators of macroinvertebrate community stability. Ecol. Indic. 2021, 129, 107892. [Google Scholar] [CrossRef]
- Cai, Y.; Xu, J.; Zhang, M.; Wang, J.; Heino, J. Different roles for geography, energy and environment in determining three facets of freshwater molluscan beta diversity at broad spatial scales. Sci. Total Environ. 2019, 659, 451–462. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Categories | Codes | Descriptions | |
|---|---|---|---|
| 1. Functional feeding groups [43] | Filter-collector | FFG_Fil | The FFG classification is based on the food consumed and also considers the morphological and behavioral characteristics used in the food acquisition [45] |
| Gather-collector | FFG_Gat | ||
| Shredder | FFG_Shr | ||
| Scraper | FFG_Scr | ||
| Predator | FFG_Pre | ||
| 2. Habit trait groups [44] | Burrower | HTG_Bur | The HTG describe the mobility and microhabitat use of the macroinvertebrates [44] |
| Climber | HTG_Clim | ||
| Clinger | HTG_Clin | ||
| Sprawler | HTG_Spr | ||
| Swimmer | HTG_Swi |
| Total | Turnover | Nestedness | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variables | AdjR2Cum | F | p | Variables | AdjR2Cum | F | p | Variables | AdjR2Cum | F | p |
| Local *** | Local *** | Local *** | |||||||||
| Depth | 0.073 | 13.981 | 0.001 | Depth | 0.018 | 4.059 | 0.009 | Depth | 0.080 | 15.372 | 0.001 |
| WT | 0.089 | 3.913 | 0.005 | WT | 0.032 | 3.387 | 0.012 | PO4 | 0.103 | 5.213 | 0.006 |
| Width | 0.103 | 3.626 | 0.008 | Width | 0.049 | 3.919 | 0.004 | NPR | 0.114 | 3.126 | 0.036 |
| PO4 | 0.114 | 3.090 | 0.014 | NPR | 0.058 | 2.497 | 0.031 | ||||
| NPR | 0.131 | 4.157 | 0.002 | ||||||||
| pH | 0.142 | 3.016 | 0.020 | ||||||||
| Velocity | 0.152 | 2.886 | 0.024 | ||||||||
| Geo *** | Geo *** | Geo *** | |||||||||
| Elevation | 0.094 | 18.125 | 0.001 | Elevation | 0.023 | 4.827 | 0.001 | Elevation | 0.118 | 23.133 | 0.001 |
| Bio15 | 0.127 | 7.319 | 0.001 | Bio15 | 0.045 | 4.800 | 0.001 | Bio15 | 0.166 | 10.516 | 0.001 |
| Shrubs | 0.139 | 3.335 | 0.010 | Shrubs | 0.183 | 4.435 | 0.011 | ||||
| Herbaceous | 0.150 | 3.016 | 0.022 | Snow.ice | 0.192 | 2.826 | 0.044 | ||||
| Urban | 0.167 | 4.365 | 0.009 | ||||||||
| Bio3 | 0.175 | 2.489 | 0.027 | ||||||||
| Spatial *** | Spatial *** | Spatial *** | |||||||||
| MEM3 | 0.037 | 7.449 | 0.001 | MEM40 | 0.023 | 4.974 | 0.001 | MEM3 | 0.076 | 14.710 | 0.001 |
| MEM2 | 0.067 | 6.149 | 0.001 | MEM6 | 0.040 | 3.810 | 0.002 | MEM6 | 0.122 | 9.581 | 0.001 |
| MEM4 | 0.091 | 5.371 | 0.001 | MEM3 | 0.054 | 3.503 | 0.007 | MEM8 | 0.157 | 7.854 | 0.003 |
| MEM7 | 0.114 | 5.210 | 0.002 | MEM4 | 0.064 | 2.731 | 0.023 | MEM11 | 0.181 | 5.737 | 0.004 |
| MEM9 | 0.136 | 5.201 | 0.001 | MEM9 | 0.074 | 2.693 | 0.019 | MEM5 | 0.202 | 5.333 | 0.003 |
| MEM15 | 0.158 | 5.130 | 0.001 | MEM24 | 0.082 | 2.400 | 0.029 | MEM1 | 0.223 | 5.200 | 0.005 |
| MEM1 | 0.178 | 4.876 | 0.002 | MEM8 | 0.089 | 2.205 | 0.044 | MEM15 | 0.240 | 4.757 | 0.005 |
| MEM5 | 0.194 | 4.225 | 0.004 | MEM35 | 0.095 | 2.144 | 0.046 | MEM7 | 0.255 | 4.164 | 0.013 |
| MEM6 | 0.209 | 4.035 | 0.008 | MEM1 | 0.102 | 2.151 | 0.036 | MEM2 | 0.270 | 4.247 | 0.010 |
| MEM16 | 0.223 | 3.883 | 0.004 | MEM9 | 0.283 | 3.688 | 0.022 | ||||
| MEM8 | 0.235 | 3.414 | 0.010 | MEM16 | 0.294 | 3.435 | 0.028 | ||||
| MEM18 | 0.246 | 3.249 | 0.010 | MEM18 | 0.304 | 3.263 | 0.033 | ||||
| MEM11 | 0.256 | 3.015 | 0.005 | MEM4 | 0.313 | 3.189 | 0.032 | ||||
| MEM28 | 0.263 | 2.452 | 0.029 | MEM29 | 0.323 | 3.121 | 0.023 | ||||
| MEM31 | 0.269 | 2.340 | 0.041 | MEM35 | 0.330 | 2.728 | 0.037 | ||||
| MEM40 | 0.275 | 2.247 | 0.035 | ||||||||
| Total | Turnover | Nestedness | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variables | AdjR2Cum | F | p | Variables | AdjR2Cum | F | p | Variables | AdjR2Cum | F | p |
| Local *** | Local | Local *** | |||||||||
| Depth | 0.095 | 18.413 | 0.001 | Depth | 0.041 | 8.048 | 0.001 | Depth | 0.097 | 18.860 | 0.001 |
| PO4 | 0.162 | 14.293 | 0.001 | PO4 | 0.065 | 5.348 | 0.006 | PO4 | 0.161 | 13.550 | 0.001 |
| SS | 0.190 | 6.684 | 0.011 | Width | 0.091 | 5.617 | 0.002 | SS | 0.190 | 6.903 | 0.004 |
| Velocity | 0.208 | 4.608 | 0.017 | Velocity | 0.208 | 4.684 | 0.017 | ||||
| NPR | 0.224 | 4.426 | 0.031 | NPR | 0.224 | 4.401 | 0.027 | ||||
| pH | 0.246 | 5.575 | 0.006 | pH | 0.246 | 5.588 | 0.007 | ||||
| Geo *** | Geo * | Geo *** | |||||||||
| Elevation | 0.132 | 26.217 | 0.001 | Elevation | 0.058 | 11.223 | 0.001 | Elevation | 0.132 | 26.185 | 0.001 |
| Bio15 | 0.185 | 11.764 | 0.001 | Bio15 | 0.110 | 10.589 | 0.001 | Bio15 | 0.188 | 12.388 | 0.001 |
| Urban | 0.210 | 6.132 | 0.011 | Urban | 0.212 | 6.145 | 0.009 | ||||
| Barrenlands | 0.222 | 3.523 | 0.030 | Barrenlands | 0.225 | 3.708 | 0.034 | ||||
| Spatial *** | Spatial | Spatial *** | |||||||||
| MEM6 | 0.057 | 10.946 | 0.001 | MEM3 | 0.037 | 7.457 | 0.001 | MEM3 | 0.061 | 11.706 | 0.001 |
| MEM3 | 0.112 | 11.363 | 0.001 | MEM9 | 0.060 | 5.035 | 0.003 | MEM6 | 0.117 | 11.520 | 0.001 |
| MEM5 | 0.151 | 8.413 | 0.001 | MEM1 | 0.081 | 4.674 | 0.009 | MEM9 | 0.154 | 8.279 | 0.002 |
| MEM9 | 0.186 | 8.063 | 0.005 | MEM5 | 0.100 | 4.520 | 0.005 | MEM5 | 0.192 | 8.569 | 0.002 |
| MEM1 | 0.217 | 7.350 | 0.004 | MEM24 | 0.112 | 3.158 | 0.029 | MEM1 | 0.226 | 8.059 | 0.001 |
| MEM10 | 0.235 | 4.853 | 0.015 | MEM6 | 0.124 | 3.176 | 0.022 | MEM10 | 0.244 | 4.938 | 0.018 |
| MEM15 | 0.251 | 4.359 | 0.031 | MEM8 | 0.135 | 2.934 | 0.035 | MEM29 | 0.262 | 4.934 | 0.014 |
| MEM7 | 0.265 | 4.087 | 0.028 | MEM29 | 0.144 | 2.675 | 0.035 | MEM7 | 0.277 | 4.318 | 0.028 |
| MEM29 | 0.278 | 3.949 | 0.032 | MEM31 | 0.151 | 2.461 | 0.039 | MEM15 | 0.292 | 4.301 | 0.029 |
| MEM31 | 0.292 | 3.946 | 0.031 | MEM8 | 0.307 | 4.309 | 0.024 | ||||
| MEM24 | 0.305 | 3.992 | 0.027 | MEM31 | 0.319 | 3.822 | 0.031 | ||||
| MEM8 | 0.318 | 4.054 | 0.032 | MEM4 | 0.331 | 3.832 | 0.034 | ||||
| MEM4 | 0.332 | 4.044 | 0.033 | MEM16 | 0.343 | 3.820 | 0.027 | ||||
| MEM16 | 0.344 | 3.821 | 0.033 | MEM11 | 0.355 | 3.705 | 0.032 | ||||
| MEM11 | 0.356 | 3.800 | 0.033 | ||||||||
| Total | Turnover | Nestedness | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Variables | AdjR2Cum | F | p | Variables | AdjR2Cum | F | p | Variables | AdjR2Cum | F | p |
| Local *** | Local *** | Local *** | |||||||||
| Depth | 0.079 | 15.278 | 0.001 | Depth | 0.024 | 5.150 | 0.003 | Depth | 0.077 | 14.813 | 0.001 |
| PO4 | 0.094 | 3.731 | 0.012 | Width | 0.039 | 3.595 | 0.007 | PO4 | 0.107 | 6.654 | 0.002 |
| NPR | 0.113 | 4.476 | 0.005 | WT | 0.050 | 2.827 | 0.015 | NPR | 0.128 | 4.819 | 0.014 |
| Width | 0.129 | 3.985 | 0.004 | pH | 0.138 | 2.998 | 0.048 | ||||
| pH | 0.137 | 2.533 | 0.050 | ||||||||
| Geo *** | Geo *** | Geo *** | |||||||||
| Elevation | 0.110 | 21.563 | 0.001 | Elevation | 0.031 | 6.252 | 0.001 | Elevation | 0.122 | 24.071 | 0.001 |
| Bio15 | 0.140 | 6.766 | 0.001 | Bio15 | 0.050 | 4.400 | 0.004 | Bio15 | 0.174 | 11.407 | 0.001 |
| Herbaceous | 0.153 | 3.445 | 0.007 | Herbaceous | 0.060 | 2.654 | 0.029 | Shrubs | 0.189 | 3.976 | 0.014 |
| Urban | 0.166 | 3.519 | 0.011 | Urban | 0.200 | 3.249 | 0.038 | ||||
| Shrubs | 0.181 | 4.093 | 0.007 | Bio3 | 0.209 | 2.839 | 0.050 | ||||
| Bio3 | 0.190 | 2.661 | 0.033 | ||||||||
| Spatial *** | Spatial *** | Spatial *** | |||||||||
| MEM3 | 0.042 | 8.284 | 0.001 | MEM40 | 0.030 | 6.201 | 0.001 | MEM3 | 0.086 | 16.541 | 0.001 |
| MEM4 | 0.066 | 5.276 | 0.004 | MEM3 | 0.047 | 3.816 | 0.004 | MEM6 | 0.133 | 10.035 | 0.001 |
| MEM2 | 0.090 | 5.323 | 0.003 | MEM6 | 0.063 | 3.819 | 0.003 | MEM11 | 0.163 | 6.948 | 0.004 |
| MEM7 | 0.114 | 5.448 | 0.002 | MEM4 | 0.073 | 2.795 | 0.014 | MEM8 | 0.194 | 7.115 | 0.003 |
| MEM15 | 0.137 | 5.162 | 0.002 | MEM35 | 0.082 | 2.514 | 0.028 | MEM5 | 0.223 | 7.137 | 0.003 |
| MEM6 | 0.156 | 4.628 | 0.001 | MEM7 | 0.089 | 2.358 | 0.023 | MEM1 | 0.243 | 5.305 | 0.007 |
| MEM1 | 0.173 | 4.403 | 0.002 | MEM15 | 0.261 | 4.868 | 0.009 | ||||
| MEM9 | 0.189 | 4.151 | 0.002 | MEM2 | 0.279 | 4.948 | 0.011 | ||||
| MEM16 | 0.204 | 3.933 | 0.009 | MEM9 | 0.293 | 4.210 | 0.019 | ||||
| MEM5 | 0.219 | 4.004 | 0.004 | MEM7 | 0.307 | 4.124 | 0.013 | ||||
| MEM11 | 0.232 | 3.657 | 0.007 | MEM18 | 0.321 | 4.094 | 0.024 | ||||
| MEM18 | 0.244 | 3.533 | 0.010 | MEM16 | 0.334 | 4.010 | 0.020 | ||||
| MEM8 | 0.255 | 3.090 | 0.015 | MEM10 | 0.344 | 3.428 | 0.033 | ||||
| MEM40 | 0.263 | 2.712 | 0.016 | MEM40 | 0.352 | 2.941 | 0.046 | ||||
| MEM29 | 0.271 | 2.681 | 0.023 | ||||||||
| MEM28 | 0.277 | 2.342 | 0.030 | ||||||||
| MEM31 | 0.284 | 2.358 | 0.043 | ||||||||
| MEM37 | 0.290 | 2.336 | 0.049 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, N.; Liu, G.; Zhang, M.; Wang, Y.; Peng, W.; Qu, X. Spatial Factors Outperform Local Environmental and Geo-Climatic Variables in Structuring Multiple Facets of Stream Macroinvertebrates’ β-Diversity. Animals 2022, 12, 2648. https://doi.org/10.3390/ani12192648
Wu N, Liu G, Zhang M, Wang Y, Peng W, Qu X. Spatial Factors Outperform Local Environmental and Geo-Climatic Variables in Structuring Multiple Facets of Stream Macroinvertebrates’ β-Diversity. Animals. 2022; 12(19):2648. https://doi.org/10.3390/ani12192648
Chicago/Turabian StyleWu, Naicheng, Guohao Liu, Min Zhang, Yixia Wang, Wenqi Peng, and Xiaodong Qu. 2022. "Spatial Factors Outperform Local Environmental and Geo-Climatic Variables in Structuring Multiple Facets of Stream Macroinvertebrates’ β-Diversity" Animals 12, no. 19: 2648. https://doi.org/10.3390/ani12192648
APA StyleWu, N., Liu, G., Zhang, M., Wang, Y., Peng, W., & Qu, X. (2022). Spatial Factors Outperform Local Environmental and Geo-Climatic Variables in Structuring Multiple Facets of Stream Macroinvertebrates’ β-Diversity. Animals, 12(19), 2648. https://doi.org/10.3390/ani12192648