
Species Composition, Growth, and Trophic Traits of Hairtail (Trichiuridae), the Most Productive Fish in Chinese Marine Fishery


 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection
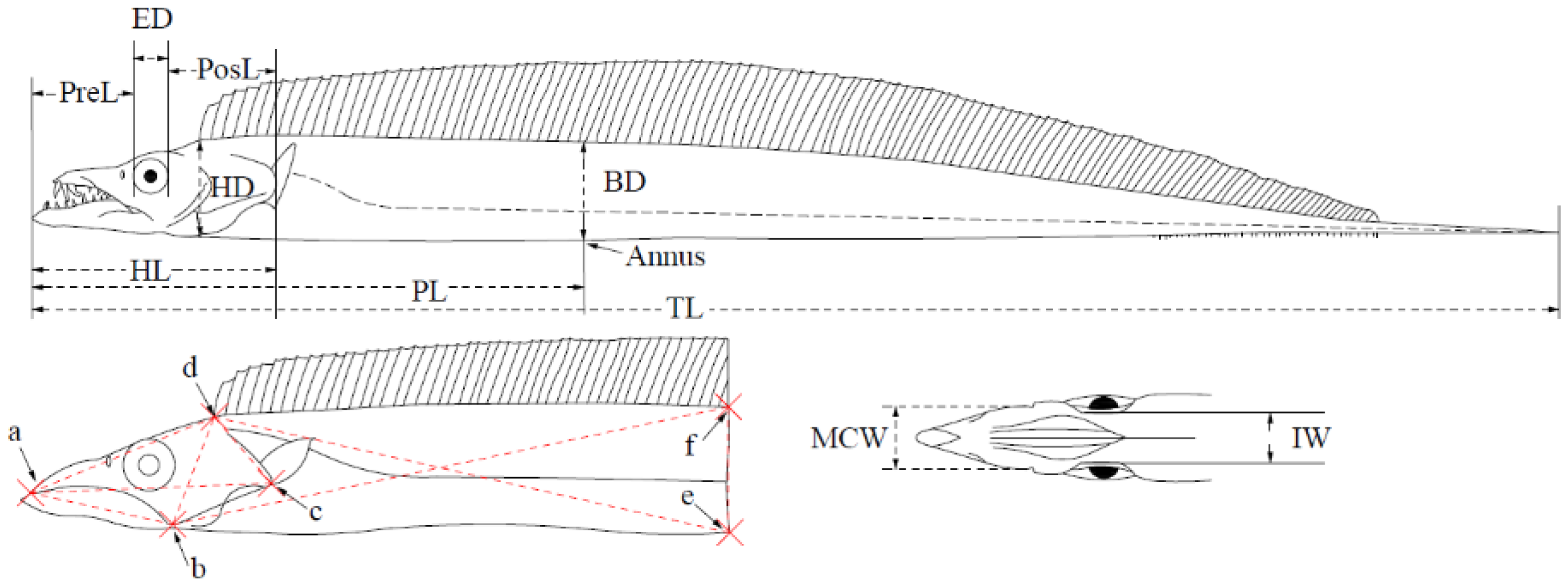
2.2. Species Morphometry and Identification
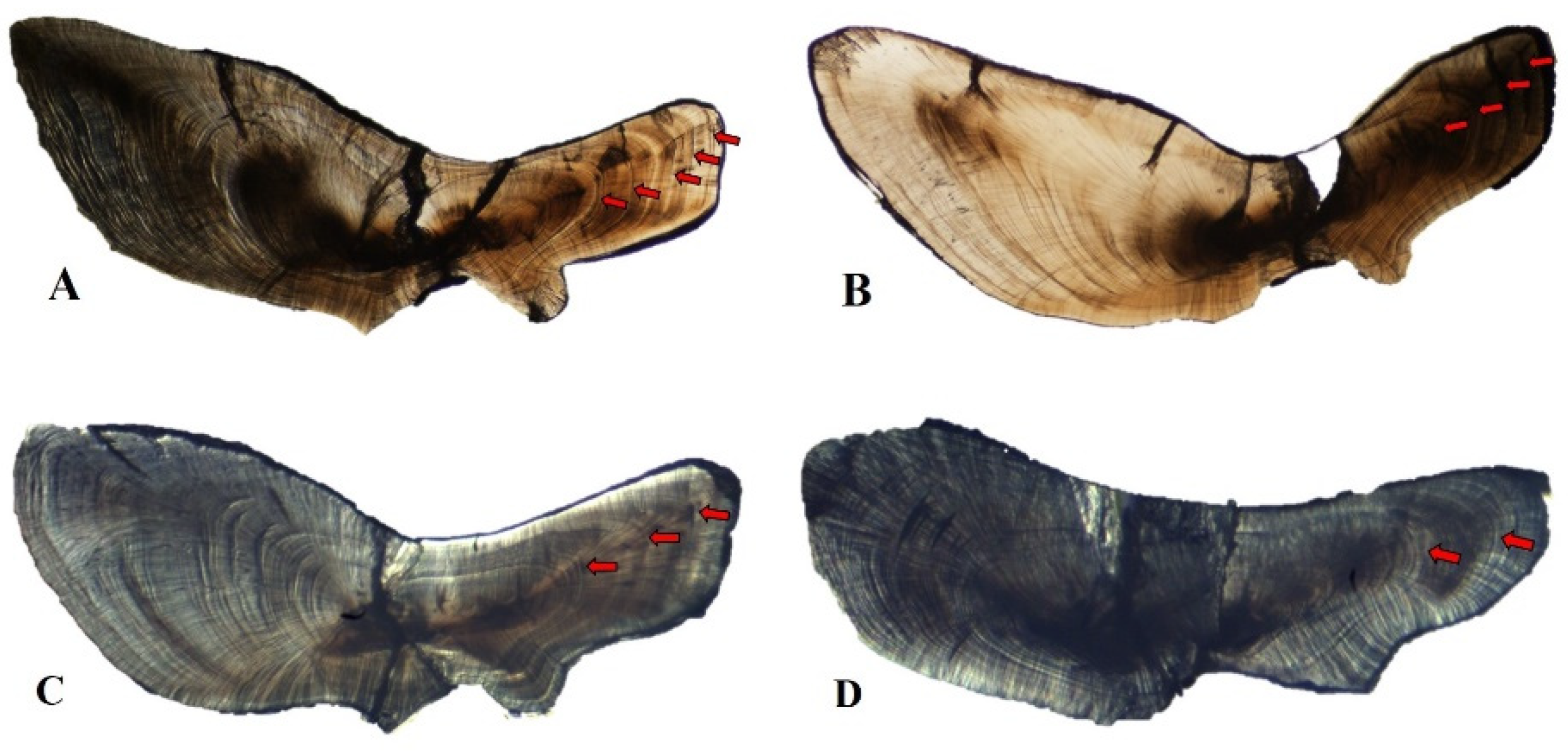
2.3. Age Determination
2.4. Stable Isotope Measurement
2.5. Data Analysis
3. Results
3.1. Common Species Composition of Trichiuridae in Chinese Seas
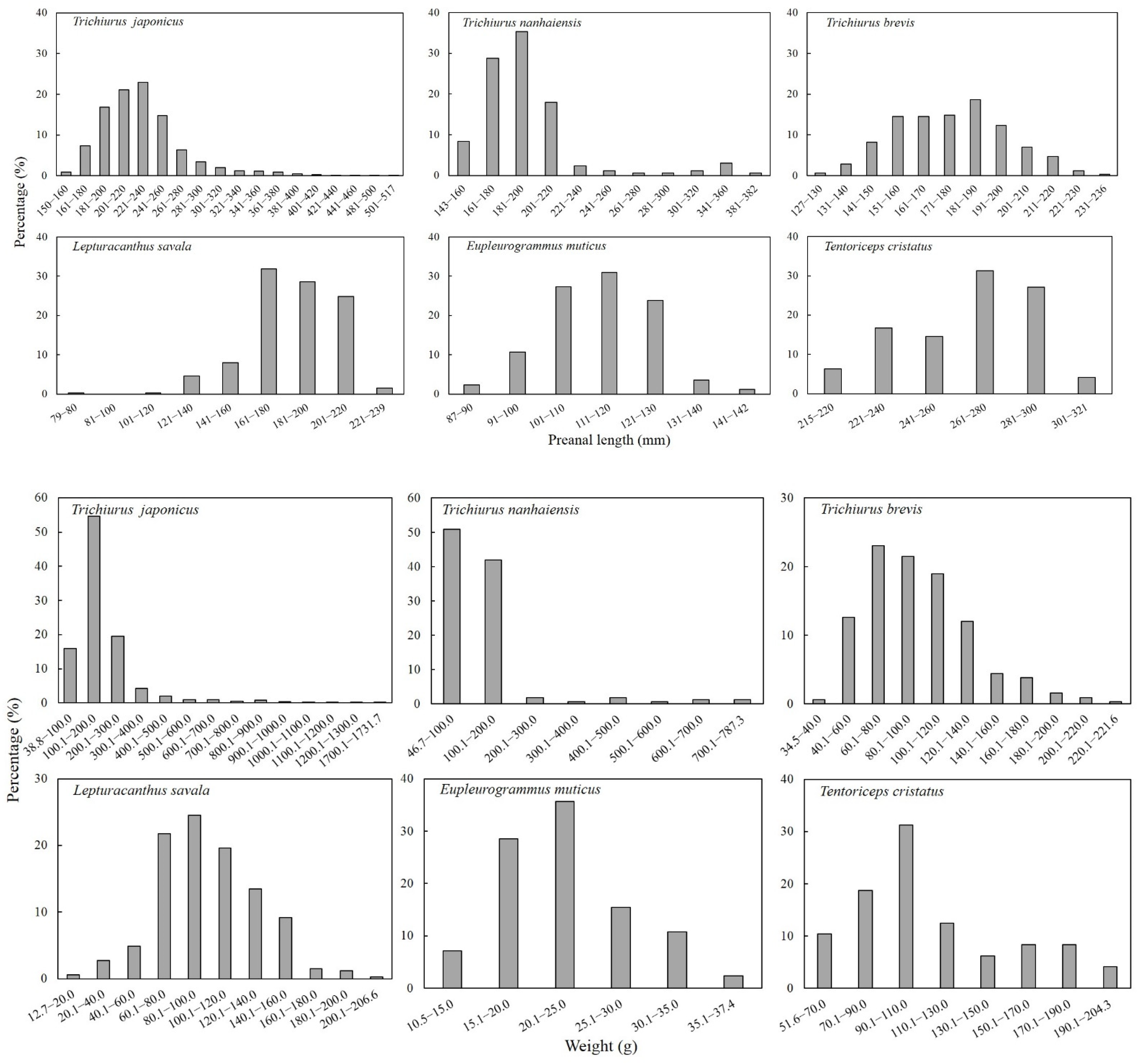
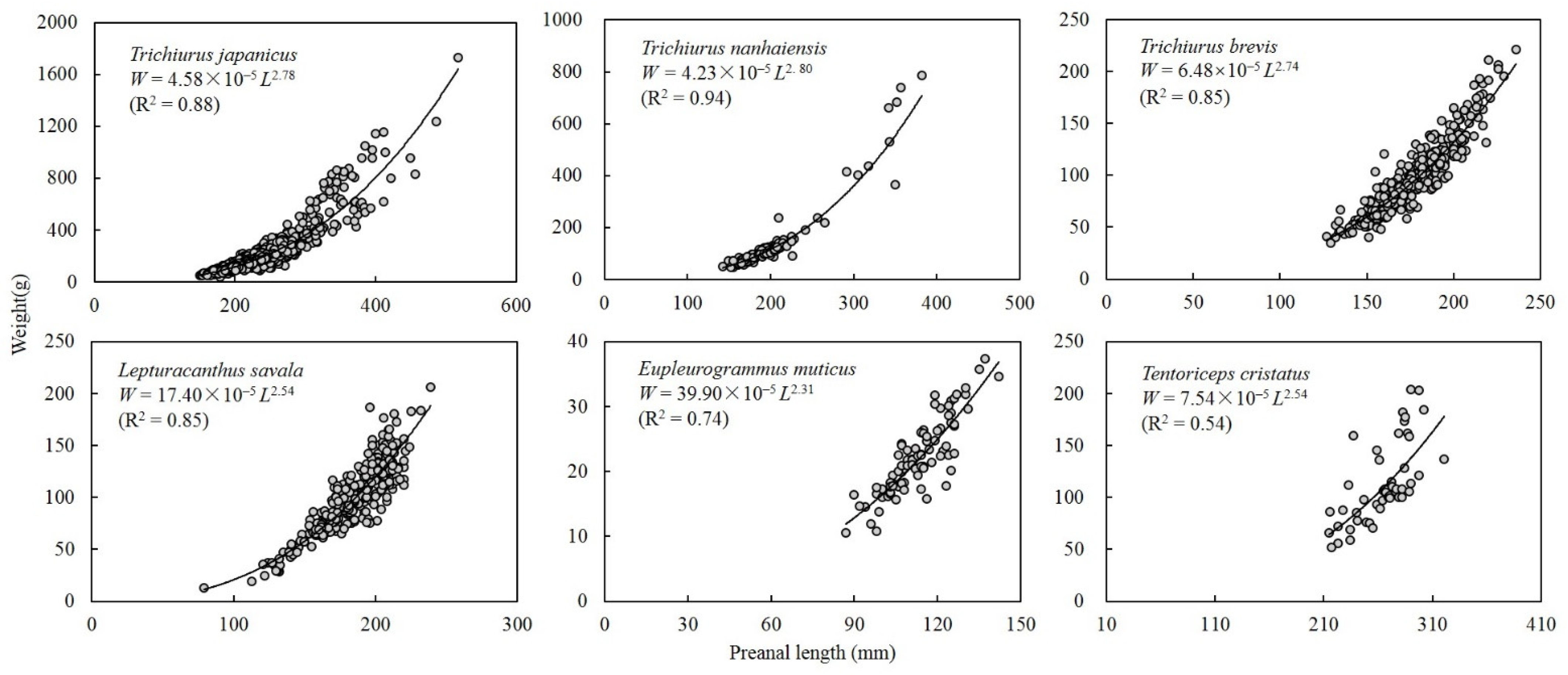
3.2. Species Morphometry
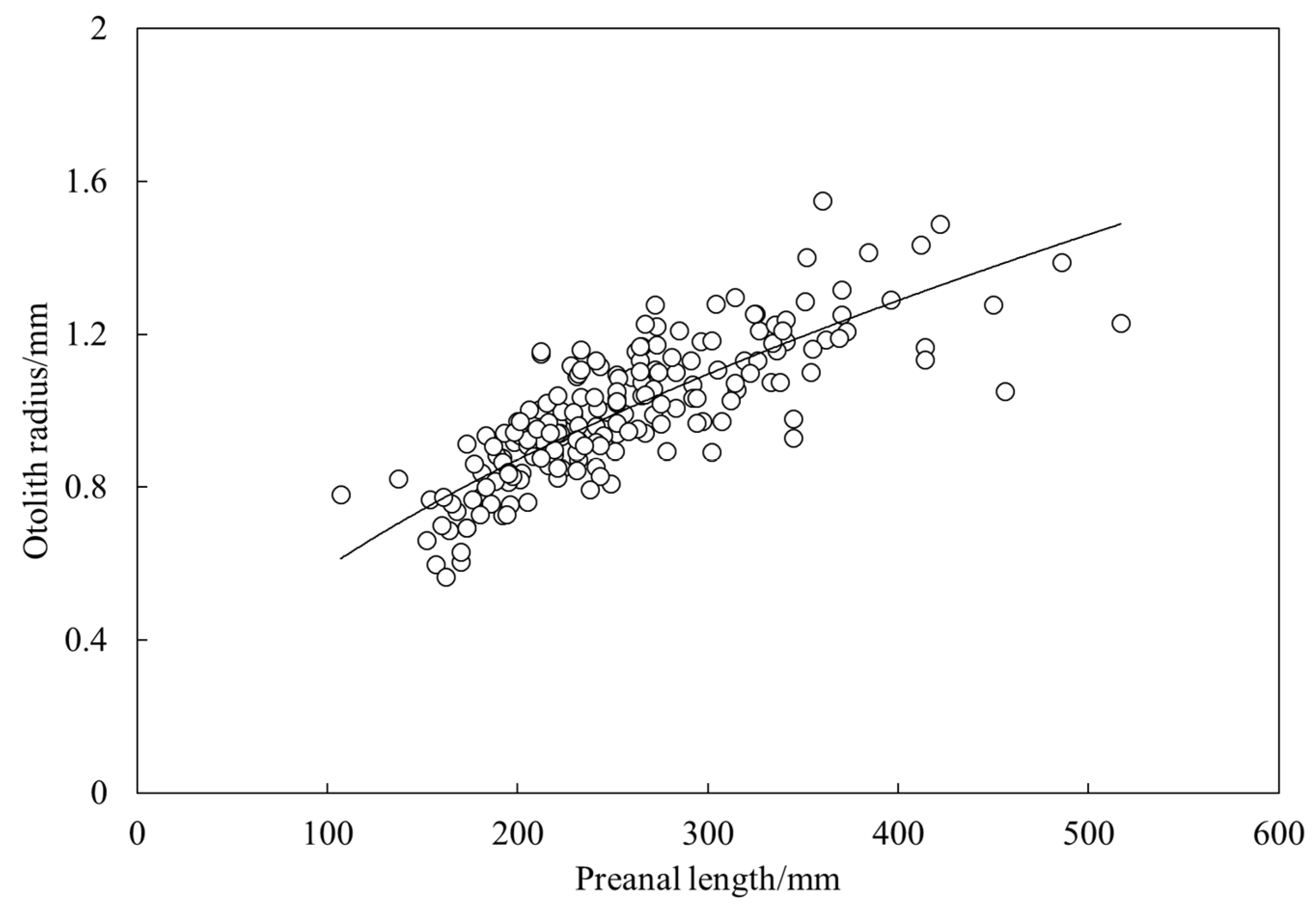
3.3. Age and Growth
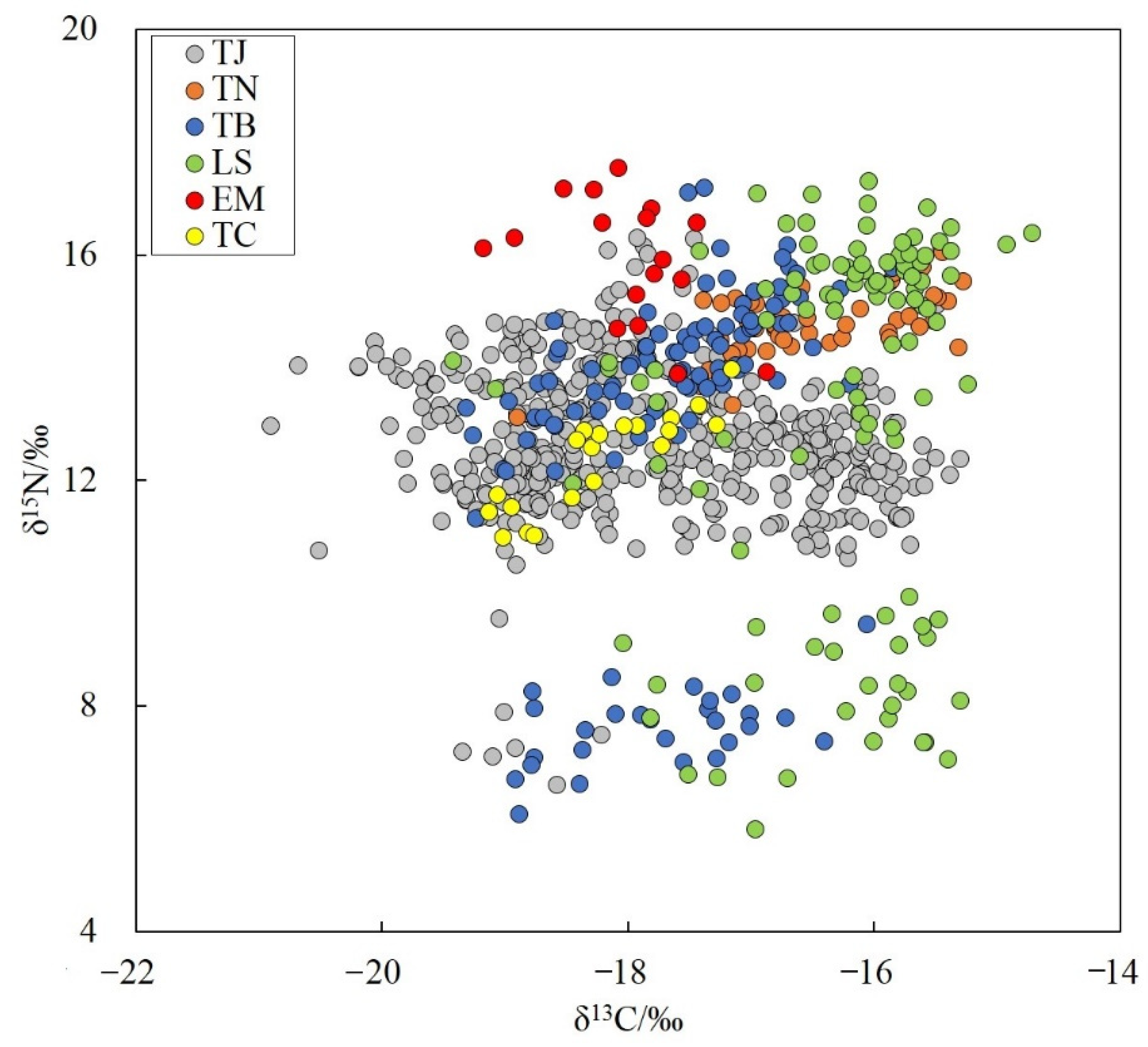
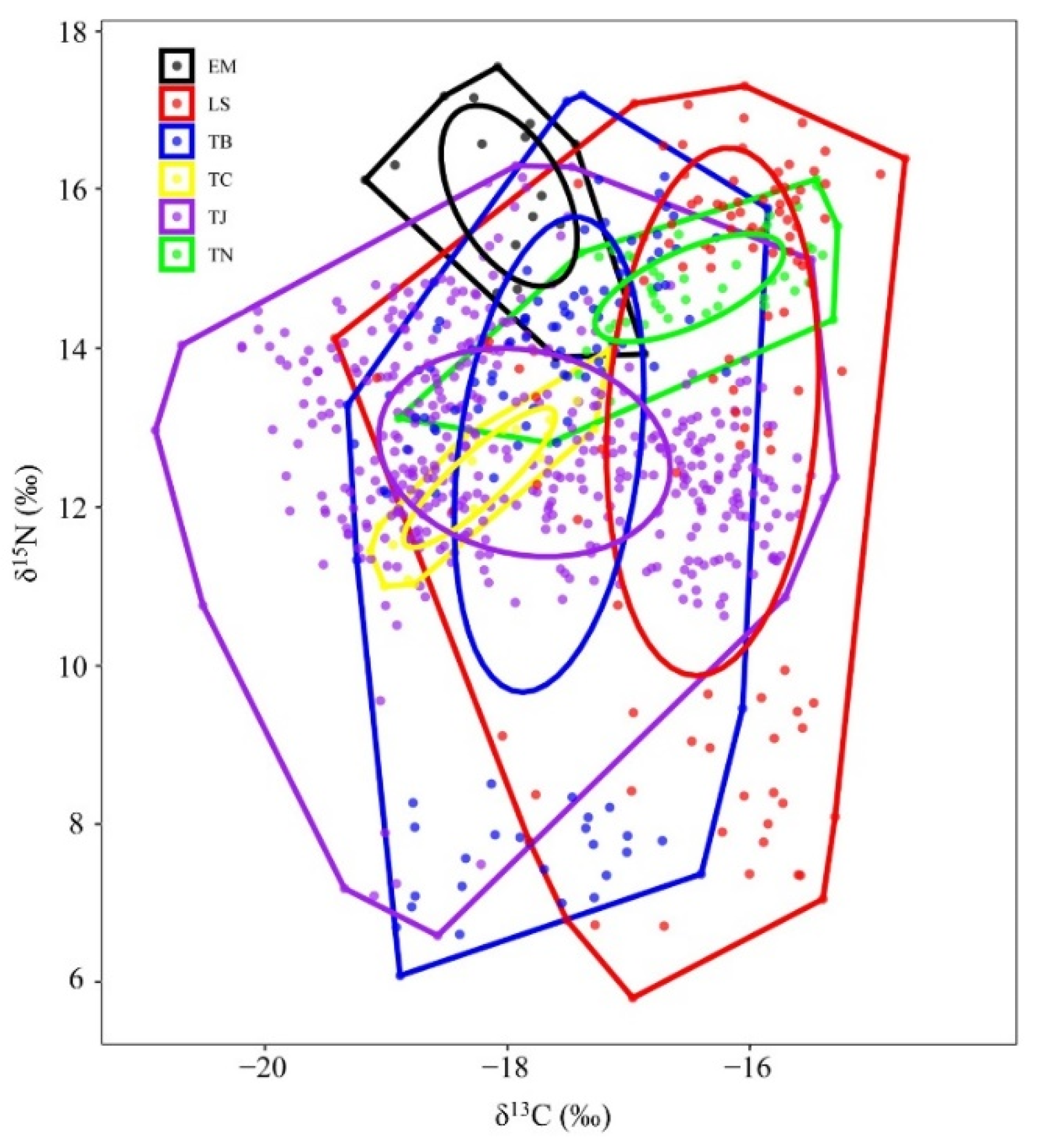
3.4. Trophic Structure and Trophic Level
4. Discussion
4.1. Composition and Distribution of Hairtails
4.2. Age and Growth
4.3. Trophic Characteristics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B


Appendix C
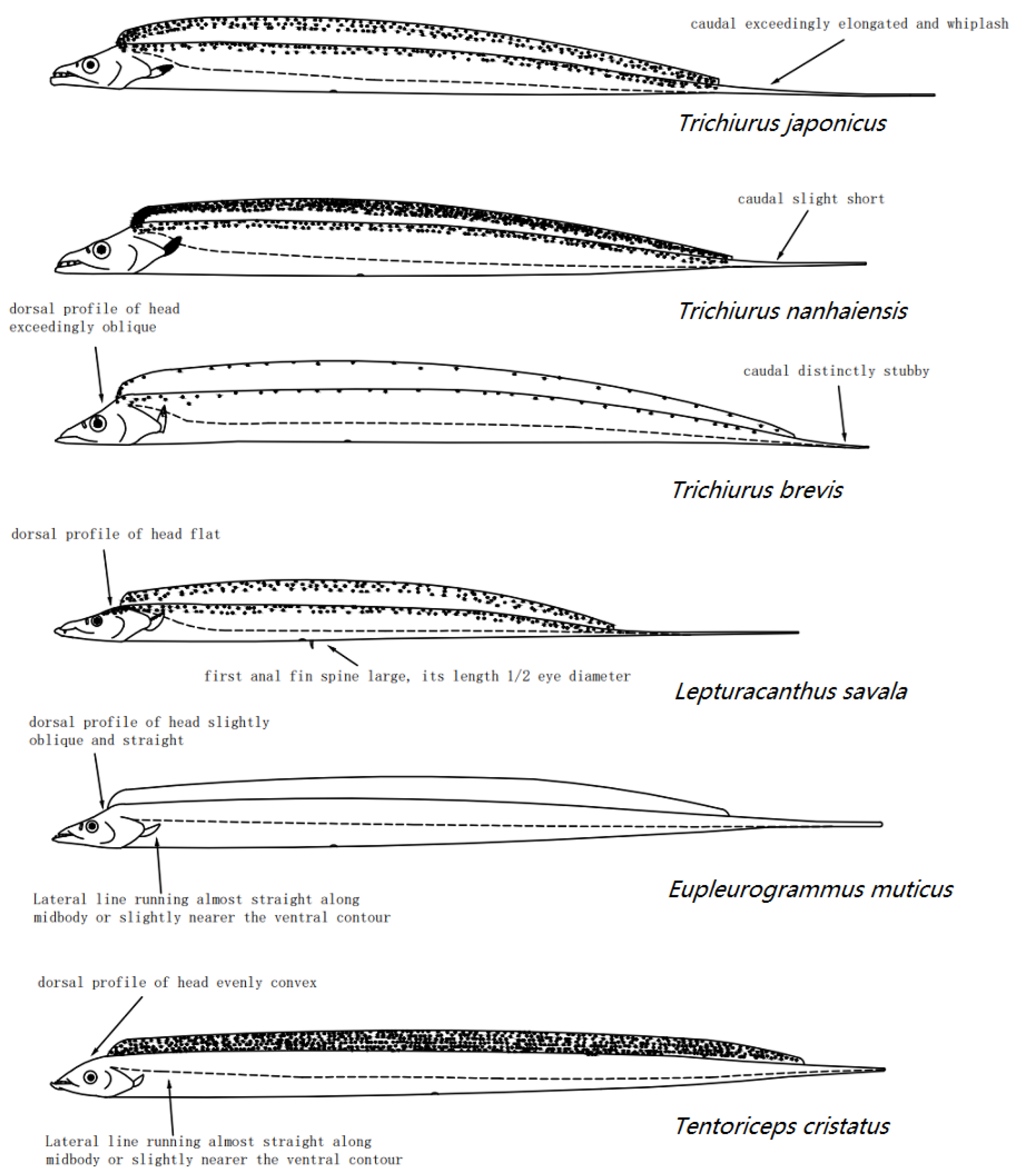
- 1(2) Lacrymal bone large; two-thirds of premaxilla sheathed by lacrymal bone. Black blotch in pectoral fin base present. Pelvic girdle and fin present, but fin reduced to a scale-like process. Dorsal soft rays 96~134, anal soft rays 73~106, pectoral soft rays 8~11, gill rakers 5~12 + 2~9……………………………………Eupleurogrammus muticus (Gray, 1831).
- 2(1) Lacrymal bone small; only one-third of premaxilla sheathed by lacrymal bone. Black blotch in pectoral fin base absent.
- 3(4) Pectoral fins short, not reaching lateral line. Dorsal profile of head evenly convex. Dorsal soft rays 129~145, anal soft rays 71~93, pectoral soft rays 10~11, gill rakers 8~11 + 4~5……………………………………………….Tentoriceps cristatus (Klunzinger, 1884).
- 4(3) Pectoral fins extending beyond lateral line. Dorsal profile of head flat.
- 5(6) First anal fin spine large, with length one-half of eye diameter; soft anal fin ray pungent spinules breaking through ventral skin. Pectoral fins yellow. Iridescence of eyes dose not turn yellow after death. Dorsal soft rays 91~134, anal soft rays 64~97, pectoral soft rays 10~11, gill rakers 0~8 + 0~45………………………………Lepturacanthus savala (Cuvier, 1829).
- 6(5) First anal fin spine small, with length less than pupil diameter; soft anal fin rays slightly breaking through ventral skin in smaller specimens. Pectoral fins grey or white.
- 7(8) Preanal length more than three times total length. Dorsal fin silver with transparency. Eye iridescence not turning yellow after death. Dorsal soft rays 123~172, anal soft rays 73~142, pectoral soft rays 10~11, gill rakers 6~24 + 4~11……………………….…….……..Trichiurus japanicus Temminck & Schlegel, 1844.
- 8(7) Preanal length less than three times total length.
- 9(10) Iridescence of eyes turning yellow after death. Dorsal fin yellowish; pectoral fins grey. Osteoma in posterior of supraoccipital present. Dorsal soft rays 129~143, anal soft rays 83~110, pectoral soft rays 11, gill rakers 5~13 + 4~8…………………………………………..….Trichiurus nanhaiensis Wang & Xu, 1992.
- 10(9) Caudal distinctly stubby. Iridescence of eyes turning yellow after death. Dorsal fin silver. Osteoma in posterior of supraoccipital absent. Dorsal soft rays 106~140, anal soft rays 60~110, pectoral soft rays 10~11, gill rakers 4~11 + 2~9………………………………………………….Trichiurus brevis Wang & You, 1992.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | TJ | TN | LS | TB | EM | TC |
|---|---|---|---|---|---|---|
| TL/PL | 3.1 | 2.7 | 3.1 | 2.9 | 3.2 | 2.7 |
| PL/HL | 2. 8 | 2.6 | 2.6 | 2.7 | 3.0 | 3.6 |
| HL/ED | 6.9 | 6.8 | 9.6 | 6.4 | 7.3 | 6.3 |
| HL/IW | 6.8 | 7.4 | 6.5 | 6.9 | 8.6 | 8.6 |
| HL/PreL | 2.9 | 3.0 | 2.7 | 3.0 | 2.7 | 2.6 |
| HL/PosL | 2.0 | 1.9 | 1.9 | 2.0 | 2.0 | 2.2 |
| BD/HD | 1.1 | 1.1 | 1.1 | 1.1 | 1.2 | 1.0 |
| HD/ED | 3.2 | 3.6 | 4.2 | 3.3 | 3.5 | 3.0 |
| HD/IW | 3.2 | 3.9 | 2.8 | 3.6 | 4.1 | 4.1 |
| PosL/PreL | 1.5 | 1.6 | 1.4 | 1.5 | 1.4 | 1.2 |
| MCL/MCW | 4.0 | 4.5 | 4.7 | 4.3 | 4.0 | 5.8 |
| Number of dorsal soft rays | 123~172 | 129~143 | 91~134 | 106~140 | 96~134 | 129–145 |
| Number of anal fin spine | 73~142 | 83~110 | 64~97 | 60~110 | 73~106 | 71–93 |
| Number of pectoral soft rays | 10~12 | 11 | 10~11 | 10~13 | 8~11 | 10~11 |
| Number of canine teeth of upper jaw | 2~6 | 3~6 | 0~6 | 0~6 | 2~5 | 0–6 |
| Number of canine teeth of lower jaw | 0~3 | 1~2 | 0~3 | 0~3 | 0~2 | 0–3 |
| Gill raker | 6~24 + 4~11 | 5~13 + 4~8 | 0~8 + 0~4 | 4~11 + 2~9 | 5~12 + 2~9 | 8~11 + 4~5 |


References
- Pauly, D.; Christensen, V.; Dalsgaard, J.; Froese, R.; Torres, F. Fishing down marine food webs. Science 1998, 279, 860–863. [Google Scholar] [CrossRef] [PubMed]
- FAO. The State of World Fisheries and Aquaculture 2018; Food and Agriculture Organization of the United Nations, Fisheries and Aquaculture Department: Rome, Italy, 2018. [Google Scholar]
- Fishbase. Available online: http://www.fishbase.org/search.php. (accessed on 10 August 2022).
- China Fishery Bureau. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2020; p. 39. [Google Scholar]
- Liu, M.; Sadovy de Mitcheson, Y. Profile of a fishery collapse: Why mariculture failed to save the large yellow croaker. Fish Fish. 2008, 9, 219–242. [Google Scholar] [CrossRef]
- Kang, B.; Liu, M.; Huang, X.; Li, J.; Yan, Y.R.; Han, C.C.; Chen, S.B. Fisheries in Chinese seas: What can we learn from controversial official fisheries statistics? Rev. Fish Biol. Fish. 2018, 28, 503–519. [Google Scholar] [CrossRef]
- Starks, E.C. The osteological characters of the scombroid fishes of the families Gempylidae, Lepidopidae and Trichiuridae; Stanford University Press: Redwood City, CA, USA, 1911. [Google Scholar]
- Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http//:researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. (accessed on 10 August 2022).
- Chen, D.G.; Zhang, M.Z. Chinese Marine Fish Records; China Ocean University Press: Qingdao, China, 2015; pp. 1902–1907. [Google Scholar]
- Hsu, K.C.; Yi, M.R.; Gu, S.; He, X.-B.; Luo, Z.-S.; Kang, B.; Lin, H.-D.; Yan, Y.-R. Composition, Demographic History, and Population Structures of Trichiurus. Front. Mar. Sci. 2022, 9, 875042. [Google Scholar] [CrossRef]
- Ricker, W.E. Computation and Interpretation of Biological Statistics of Fish Population; Bulletin of the Fisheries Research Board of Canada: Ottawa, MTL, Canada, 1975; p. 332. [Google Scholar]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Post, D.M. Using Stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Li, Z.Y.; Jin, X.S.; Zhuang, Z.M.; Tang, Q.S.; Su, Y.Q. Applications of stable isotope techniques in aquatic ecological studies. Acta Ecol. Sin. 2005, 25, 3052–3060. [Google Scholar]
- Layman, C.A.; Arrington, D.A.; Montana, C.G.; Post, D.M. Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 2007, 88, 42–48. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER–Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef]
- Committee of Age Reading Experts. Manual on Generalized Age Determination Procedures for Groundfish; Pacific States Marine Fisheries Commission: Portland, OR, USA, 1982; pp. 10–24. [Google Scholar]
- Wang, X.F.; Xu, L.X.; Zhu, G.P.; Wang, C.-L. Age identification and growth characteristics of Katsuwonus pelamis in western and central Pacific Ocean. Chin. J. Appl. Ecol. 2010, 21, 756–762. [Google Scholar]
- Du, S.Q.; Feng, B.; Hou, G.; Lu, H.-S.; Yan, Y.-R. Age and growth of moonfish, Mene maculata from mouth of the Beibu Gulf, South China Sea. J. Fish. China 2012, 36, 556–583. [Google Scholar] [CrossRef]
- Xu, J.; Xie, P.; Zhang, M.; Yang, H. Variation in stable isotope signatures of seston and a zooplanktivorous fish in a eutrophic Chinese lake. Hydrobiologia 2005, 541, 215–220. [Google Scholar] [CrossRef]
- He, X.B.; Zhu, D.W.; Zhao, C.X.; Yan, Y.R.; Kang, B. Feeding habit of Asian moon scallop (Amusium pleuronectes) and as an isotopic baseline indicator in the Beibu Gulf, South China Sea. J. Shellfish Res. 2019, 38, 245–252. [Google Scholar] [CrossRef]
- Zhan, B.Y. Fish stock Assessment; China Agriculture Press: Beijing, China, 1995; pp. 26–27. [Google Scholar]
- Coplen, T.B. Reporting of stable hydrogen, carbon, and oxygen isotopic abundances. Pure Appl. Chem. 1994, 66, 273–276. [Google Scholar] [CrossRef]
- DeNiro, M.J.; Epstein, S. Influence of diet on the distribution of carbon isotopes in animals. Geochim. Cosmochim. Acta 1978, 42, 495–506. [Google Scholar] [CrossRef]
- Mariotti, A. Atmospheric nitrogen is a reliable standard for natural 15N abundance measurements. Nature 1983, 303, 685–687. [Google Scholar] [CrossRef]
- Mariotti, A. Natural 15N abundance measurements and atmospheric nitrogen standard calibration. Nature 1984, 311, 251–252. [Google Scholar] [CrossRef]
- Zhu, Y.D. Index Piscium Sinensium; Department of Biology, Shanghai St. John’s University: Shanghai, China, 1931. [Google Scholar]
- Zhu, Y.D.; Wu, H.L. New genu and species of Chinese hairtail: Pseudoxymetopon sinensis gen. and sp.nov. Acta Zool. Sin. 1962, 14, 76–78. [Google Scholar]
- Wu, H.L.; Shao, G.Z.; Lai, C.F. Latin-Chinese Dictionary of Fish Names by Classification; China Ocean University Press: Qingdao, China, 2017; p. 322. [Google Scholar]
- Lee, S.C.; Chang, K.H.; Wu, W.L.; Yang, H.C. Taiwan Ribbonfishes (Perciformes, Trichiuridae). Bull. Inst. Zool. Acad. Sin. 1977, 16, 77–84. [Google Scholar]
- Lin, X.Y.; Shen, X.M. Preliminary research on species identification of largehead hairtail in the Donghai Sea and Huanghai Sea. J. Fish. China 1986, 10, 339–350. [Google Scholar]
- Li, C.S. Hairtail fishes from Chinese coastal waters (Trichiuridae). Mar. Sci. 1992, 4, 212–219. [Google Scholar]
- Hsu, K.C.; Shih, N.T.; Ni, I.; Shao, K.-T. Genetic variation in Trichiurus lepturus (Perciformes: Trichiuridae) in waters off Taiwan: Several species or cohort contribution? Raffles Bull. Zool. 2007, 14, 215–220. [Google Scholar]
- Wu, R.X.; Guo, L.J.; Liu, J. Genetic variation of Trichiurus lepturus and phylogenetic relationship among its closed species. Acta Zootaxonomica Sin. 2011, 36, 648–655. [Google Scholar]
- Wang, K.L.; Liu, L.Y.; You, F.; Xu, C. Studies on the genetic variation and systematics of the hairtails fishes from the South China Sea. Mar. Sci. 1992, 3, 69–72. [Google Scholar]
- Wang, K.L.; Zhang, P.J.; Liu, L.Y.; Xu, C. Classification of Hairtail in the Chinese Offshore. Acta Oceanol. Sinaca 1993, 15, 77–83. [Google Scholar]
- Cheng, Q.T.; Zheng, B.S. Systematic Retrieval of Chinese fish; Science Press: Beijing, China, 1987; pp. 415–416. [Google Scholar]
- The Fish Database of Taiwan. Available online: https://fishdb.sinica.edu.tw/chi/species.php?science=Trichiurus%20brevis&tree=y (accessed on 15 August 2022).
- Liu, Q.Y.; Chen, W.F.; Zhan, H.X. Discovery of Pseudoxymetopon sinensis in the Coasts of Fujian and Guangdong. Chin. J. Zool. 1965, 3, 108. [Google Scholar]
- Mi, C.D. A study on resources, stock structure and variation of reproductive habit of hairtail, Trichiurus haumela in east China sea. J. Fish. Sci. China 1997, 4, 8–15. [Google Scholar]
- Heino, M.; Diaz Pauli, B.; Dieckmann, U. Fisheries-induced evolution. Annu. Rev. Ecol. Evol. Syst. 2015, 46, 461–480. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.M. Study on the biology and resource variation of hairtail Trichiurus haumela (Forskal) in Fujian sea area. South China Fish. Sci. 2005, 1, 32–36. [Google Scholar]
- Jellyman, P.G.; Booker, D.J.; Crow, S.K.; Bonnett, M.L. Does one size fit all? An evaluation of length-weight relationships for New Zealand’s freshwater fish species. N. Z. J. Mar. Freshw. Res. 2013, 47, 450–468. [Google Scholar] [CrossRef]
- Froese, R.; Thorson, J.T.; Reyes, R.B. A Bayesian approach for estimating length-weight relationships in fishes. J. Appl. Ichthyol. 2014, 30, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Misu, H. Fisheries biology on the ribbonfish (Trichiurus lepturus) in the East China and Yellow Seas. Bull. Seikai Reg. 1964, 32, 1–58. [Google Scholar]
- Ye, Q.T.; Huang, P.M.; Zhang, Z.L. Study on the Utilization of Trichiurus nanhaiensis in the Adjacent Waters of Taiwan Bank. J. Fujian Fish. 1999, 21, 35–41. [Google Scholar]
- Martins, A.S.; Haimovici, M.; Palacios, R. Diet and feeding of the cutlassfish Trichiurus lepturus in the Subtropical Convergence Ecosystem of southern Brazil. J. Mar. Biol. Assoc. UK 2005, 85, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.R.; Chen, J.L.; Hou, G.; Lu, H.-S.; Jin, X.-S. Feeding habits of Trichiurus lepturus in Beibu Gulf of South China Sea. Chin. J. Appl. Ecol. 2010, 21, 749–755. [Google Scholar]
- Bittar, V.T.; Beneditto, A.P.M.D. Diet and potential feeding overlap between Trichiurus lepturus (Osteichthyes: Perciformes) and Pontoporia blainvillei (Mammalia: Cetacea) in Northern Rio de Janeiro, Brazil. Zoologia 2009, 26, 374–378. [Google Scholar] [CrossRef]
- Liu, B.Z. The Studies on the Food Web Structures of Northern China Sea by Stable Isotopes Analysis. Doctoral Thesis, Dalian Maritime University, Dalian, China, 2013. [Google Scholar]
- Ma, Q.Y.; Han, D.Y.; Liu, H.; Xue, Y.; Ji, Y.P.; Ren, Y.P. Construction of a continuous trophic spectrum for the food web in Jiaozhou Bay using stable isotope analyses. Acta Ecol. Sin. 2015, 35, 7207–7218. [Google Scholar]
- Yan, Y.R.; Zhang, W.K.; Lu, H.S.; Wang, X.; Lai, J. Using stable isotopes to analyze feeding habits trophic position of hairtail (Trichiurus lepturus) from the Beibu Gulf, South China Sea. Oceanol. Limnol. Sin. 2012, 43, 192–200. [Google Scholar]






| Sea Area | Site Number | Site Name | Longitude | Latitude |
|---|---|---|---|---|
| Bohai and Yellow Seas | 1 | Dandong (DD) | 123.90 | 40.00 |
| 2 | Dalian (DL) | 121.50 | 39.00 | |
| 3 | Weihai (WH) | 122.20 | 37.50 | |
| 4 | Qingdao (QD) | 120.20 | 36.00 | |
| East China Sea | 5 | Lianyungang (LY) | 119.40 | 34.70 |
| 6 | Nantong (NT) | 121.00 | 32.00 | |
| 7 | Zhoushan (ZS) | 122.50 | 30.20 | |
| 8 | Wenzhou (WZ) | 121.00 | 28.00 | |
| 9 | Ningde (ND) | 120.20 | 27.00 | |
| 10 | Quanzhou (QZ) | 118.70 | 24.80 | |
| South China Sea | 11 | Shantou (ST) | 116.50 | 23.25 |
| 12 | Shenzhen (SZ) | 114.00 | 22.50 | |
| 13 | Yangjiang (YJ) | 111.91 | 21.63 | |
| 14 | Zhanjiang (ZJ) | 110.50 | 21.00 | |
| 15 | Beihai (BH) | 109.20 | 21.60 | |
| 16 | Wenchang (WC) | 110.75 | 19.60 | |
| 17 | Sanya (SY) | 109.60 | 18.30 |
| Species | Preanal Length (mm) | Weight (g) | ||||
|---|---|---|---|---|---|---|
| Min | Max | Mean ± SD | Min | Max | Mean ± SD | |
| Trichiurus japonicus | 150 | 517 | 230 ± 44 | 38.9 | 1731.7 | 192.2 ± 143 |
| Trichiurus nanhaiensis | 143 | 382 | 195 ± 40 | 46.8 | 787.3 | 123.0 ± 114 |
| Trichiurus brevis | 127 | 236 | 177 ± 21 | 34.5 | 211.6 | 98.0 ± 35 |
| Lepturacanthus savala | 79 | 239 | 183 ± 23 | 12.8 | 206.6 | 99.5 ± 32 |
| Eupleurogrammus muticus | 87 | 142 | 113 ± 11 | 10.5 | 37.4 | 22.4 ± 6 |
| Tentoriceps cristatus | 215 | 321 | 264 ± 25 | 51.6 | 204.2 | 113.5 ± 39 |
| Species | CR | NR | TA | CD | MNND | SDNND | SEA |
|---|---|---|---|---|---|---|---|
| Trichiurus japonicus | 5.61 | 9.71 | 35.77 | 1.57 | 0.12 | 0.13 | 4.80 |
| Trichiurus nanhaiensis | 3.62 | 3.32 | 6.04 | 0.85 | 0.19 | 0.21 | 1.22 |
| Trichiurus brevis | 3.47 | 11.11 | 28.67 | 2.58 | 0.22 | 0.21 | 6.90 |
| Lepturacanthus savala | 4.70 | 11.50 | 35.34 | 3.01 | 0.28 | 0.22 | 8.82 |
| Eupleurogrammus muticus | 2.30 | 3.66 | 4.37 | 1.07 | 0.33 | 0.17 | 1.69 |
| Tentoriceps cristatus | 1.98 | 2.98 | 2.07 | 0.92 | 0.23 | 0.14 | 0.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Luo, Z.; Zhao, C.; Huang, L.; Yan, Y.; Kang, B. Species Composition, Growth, and Trophic Traits of Hairtail (Trichiuridae), the Most Productive Fish in Chinese Marine Fishery. Animals 2022, 12, 3078. https://doi.org/10.3390/ani12223078
He X, Luo Z, Zhao C, Huang L, Yan Y, Kang B. Species Composition, Growth, and Trophic Traits of Hairtail (Trichiuridae), the Most Productive Fish in Chinese Marine Fishery. Animals. 2022; 12(22):3078. https://doi.org/10.3390/ani12223078
Chicago/Turabian StyleHe, Xiongbo, Zhisen Luo, Chunxu Zhao, Liangliang Huang, Yunrong Yan, and Bin Kang. 2022. "Species Composition, Growth, and Trophic Traits of Hairtail (Trichiuridae), the Most Productive Fish in Chinese Marine Fishery" Animals 12, no. 22: 3078. https://doi.org/10.3390/ani12223078
APA StyleHe, X., Luo, Z., Zhao, C., Huang, L., Yan, Y., & Kang, B. (2022). Species Composition, Growth, and Trophic Traits of Hairtail (Trichiuridae), the Most Productive Fish in Chinese Marine Fishery. Animals, 12(22), 3078. https://doi.org/10.3390/ani12223078