
Effects of Eugenol on Water Quality and the Metabolism and Antioxidant Capacity of Juvenile Greater Amberjack (Seriola dumerili) under Simulated Transport Conditions



Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Ethics Statement
2.2. Experiment Design and Fish
2.3. Sampling
2.4. Measurement of Water Quality
2.5. Assay of Metabolic and Antioxidant-Related Enzymes
2.6. Relative Expression of Metabolic and Antioxidant-Related Genes
2.7. Statistical Analysis
3. Results
3.1. Water Quality after Simulated Transport Conditions
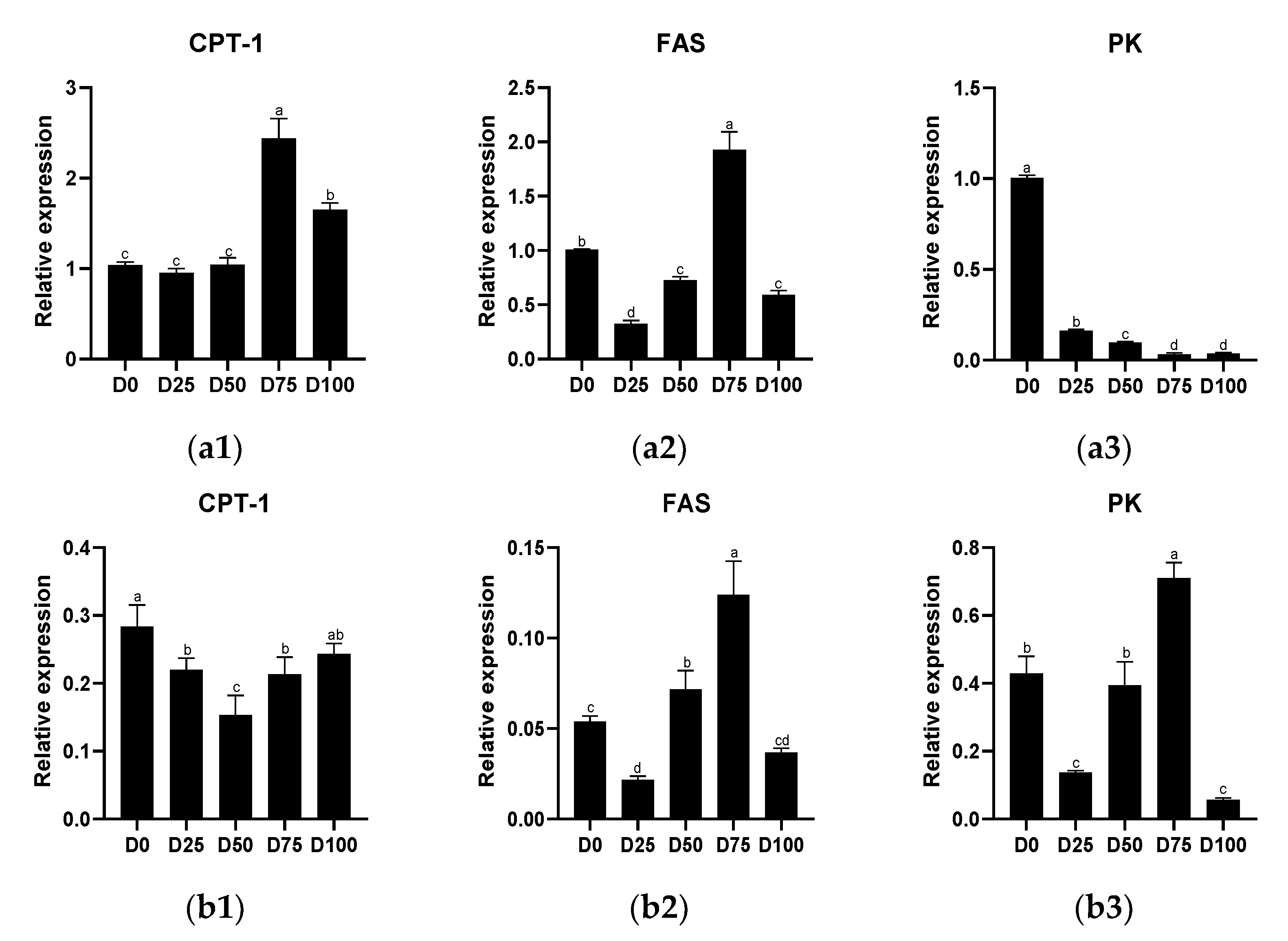
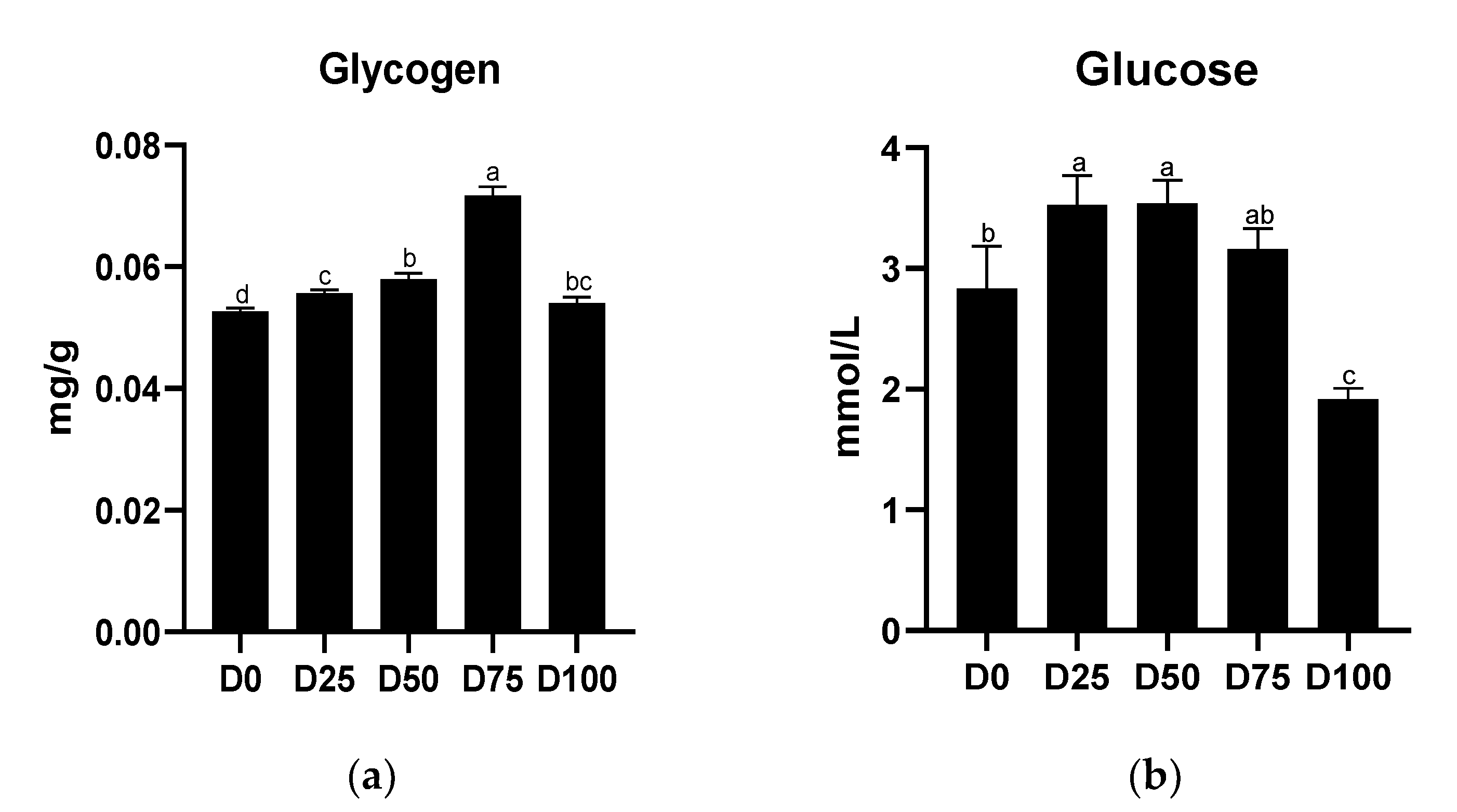
3.2. Metabolic Capacity after Simulated Transport Conditions
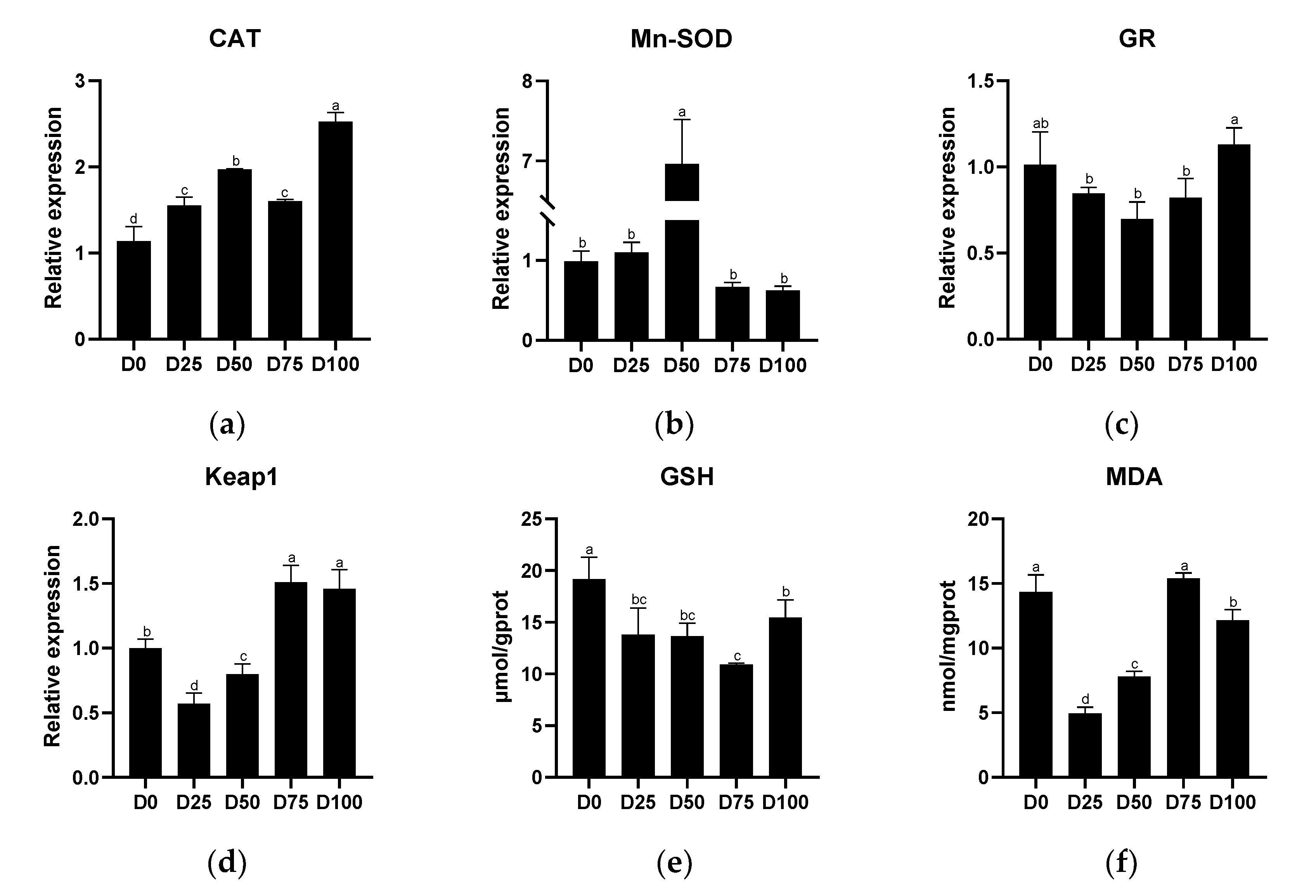
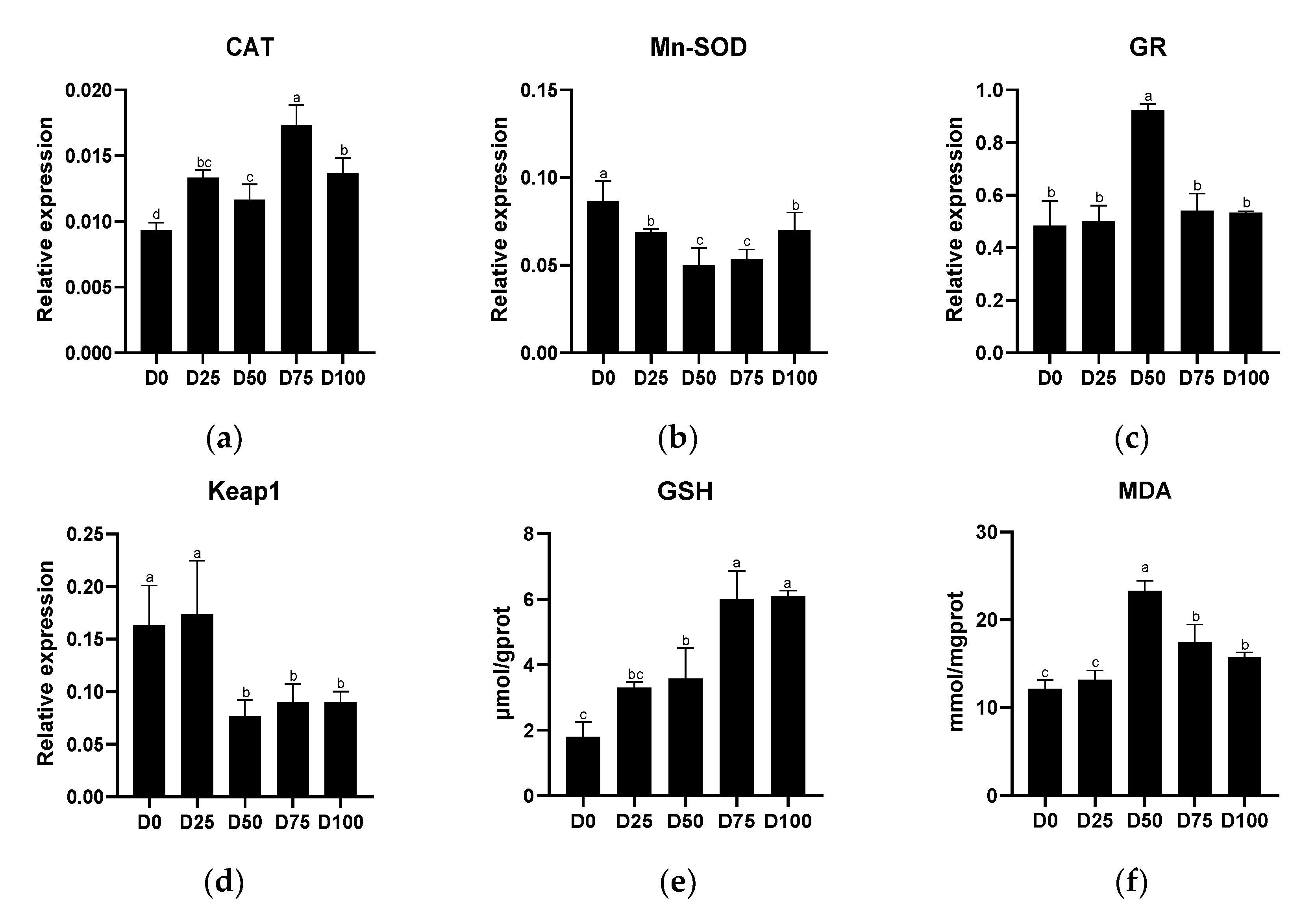
3.3. Antioxidant Capacity after Simulated Transport Conditions
4. Discussion
4.1. The Effects of Eugenol on Liver and Gill Metabolic Capacity after Simulated Transport
4.2. The Effects of Eugenol on Liver and Gill Antioxidant Capacity after Simulated Transport
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stankus, A. State of world aquaculture 2020 and regional reviews: FAO webinar series. FAO Aquac. Newsl. 2021, 63, 17–18. [Google Scholar]
- Wu, S.; Tseng, Y.; Lin, J.; Pan, B.S. Mitigation of stress and water deterioration with a root extract of Glycine tomentella during simulated transport of orange-spotted grouper (Epinephelus coioides). Aquaculture 2020, 514, 734485. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, Y.; Liu, Y.; Adányi, N.; Zhang, X. Effects of waterless live transportation on survivability, physiological responses and flesh quality in Chinese farmed sturgeon (Acipenser schrenckii). Aquaculture 2020, 518, 734834. [Google Scholar] [CrossRef]
- Gomes, L.C.; Roubach, R.; Araujo-Lima, C.A.R.M.; Chippari-Gomes, A.R.; Lopes, N.P.; Urbinati, E.C. Effect of Fish Density During Transportation on Stress and Mortality of Juvenile Tambaqui Colossoma macropomum. J. World Aquacult. Soc. 2003, 34, 76–84. [Google Scholar] [CrossRef]
- Pan, C.; Chien, Y.; Wang, Y. The antioxidant capacity response to hypoxia stress during transportation of characins (Hyphessobrycon callistus Boulenger) fed diets supplemented with carotenoids. Aquac. Res. 2010, 41, 973–981. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Monaghan, P.; Metcalfe, N.B.; Torres, R. Oxidative stress as a mediator of life history trade-offs: Mechanisms, measurements and interpretation. Ecol. Lett. 2009, 12, 75–92. [Google Scholar] [CrossRef]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2: INrf2 (Keap1) signaling in oxidative stress. Free Radical Bio. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1–Nrf2 system. Free Radic. Bio. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, H.; Yamamoto, M. Nrf2-Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef]
- Mazzola, A.; Favaloro, E.; Sarà, G. Cultivation of the Mediterranean amberjack, Seriola dumerili (Risso, 1810), in submerged cages in the Western Mediterranean Sea. Aquaculture 2000, 181, 257–268. [Google Scholar] [CrossRef]
- Jover, M.; García-Gómez, A.; Tomás, A.; De la Gándara, F.; Pérez, L. Growth of mediterranean yellowtail (Seriola dumerilii) fed extruded diets containing different levels of protein and lipid. Aquaculture 1999, 179, 25–33. [Google Scholar] [CrossRef]
- Singh, R.K.; Vartak, V.R.; Balange, A.K.; Ghughuskar, M.M. Water quality management during transportation of fry of Indian major carps, Catla catla (Hamilton), Labeo rohita (Hamilton) and Cirrhinus mrigala (Hamilton). Aquaculture 2004, 235, 297–302. [Google Scholar] [CrossRef]
- Park, I.S.; Park, M.O.; Hur, J.W.; Kim, D.S.; Chang, Y.J.; Kim, Y.J.; Park, J.Y.; Johnson, S.C. Anesthetic effects of lidocaine-hydrochloride on water parameters in simulated transport experiment of juvenile winter flounder, Pleuronectes americanus. Aquaculture 2009, 294, 76–79. [Google Scholar] [CrossRef]
- Harmon, T.S. Methods for reducing stressors and maintaining water quality associated with live fish transport in tanks: A review of the basics. Rev. Aquacult. 2009, 1, 58–66. [Google Scholar] [CrossRef]
- Barata, M.; Soares, F.; Aragão, C.; Almeida, A.C.; Pousão-Ferreira, P.; Ribeiro, L. Efficiency of 2-phenoxyethanol and Clove Oil for Reducing Handling Stress in Reared Meagre, Argyrosomus regius (Pisces: Sciaenidae). J. World Aquacult. Soc. 2016, 47, 82–92. [Google Scholar] [CrossRef]
- Kaiser, H.; Vine, N. The effect of 2-phenoxyethanol and transport packing density on the post-transport survival rate and metabolic activity in the goldfish, Carassius Auratus. Aquar. Sci. Conserv. 1998, 2, 1–7. [Google Scholar] [CrossRef]
- Baynes, R.; Riviere, J.E. Strategies for Reducing Drug and Chemical Residues in Food Animals: International Approaches to Residue Avoidance, Management, and Testing; Wiley: Hoboken, NJ, USA, 2014; ISBN 1118872819. [Google Scholar]
- Baynes, R.E.; Dedonder, K.; Kissell, L.; Mzyk, D.; Marmulak, T.; Smith, G.; Tell, L.; Gehring, R.; Davis, J.; Riviere, J.E. Health concerns and management of select veterinary drug residues. Food Chem. Toxicol. 2016, 88, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Netto, J.D.L.; Oliveira, R.S.; Copatti, C.E. Efficiency of essential oils of Ocimum basilicum and Cymbopogum flexuosus in the sedation and anaesthesia of Nile tilapia juveniles. An. Acad. Bras. Ciênc. 2017, 89, 2971–2974. [Google Scholar] [CrossRef]
- Correia, A.M.; Pedrazzani, A.S.; Mendonça, R.C.; Massucatto, A.; Ozório, R.A.; Tsuzuki, M.Y. Basil, tea tree and clove essential oils as analgesics and anaesthetics in Amphiprion clarkii (Bennett, 1830). Braz. J. Biol. 2017, 78, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Paradewi, M.; Saputri, D.M.; Sulmartiwi, L.; Suprapto, H.; Satapornvanit, K. The Effect of Rubber Seed (Havea brasiliensis) as Anesthetic on Asian Sea Bass (Lates calcarifer, Centropomidae) during Transport. J. Fish. Environ. 2021, 45, 78–84. [Google Scholar]
- Irawan, O.; Efendi, E.; Ali, M. Efek pelarut yang berbeda terhadap toksisitas ekstrak akar tuba (Derris elliptica). e-J. Rekayasa Dan Teknol. Budid. Perair. 2014, 2, 259–266. [Google Scholar]
- Hoskonen, P.; Pirhonen, J. The effect of clove oil sedation on oxygen consumption of six temperate-zone fish species. Aquac. Res. 2004, 35, 1002–1005. [Google Scholar] [CrossRef]
- Peake, S. Sodium Bicarbonate and Clove Oil as Potential Anesthetics for Nonsalmonid Fishes. N. Am. J. Fish. Manag. 1998, 18, 919–924. [Google Scholar] [CrossRef]
- Park, M.O.; Im, S.; Seol, D.; Park, I. Efficacy and physiological responses of rock bream, Oplegnathus fasciatus to anesthetization with clove oil. Aquaculture 2009, 287, 427–430. [Google Scholar] [CrossRef]
- Javahery, S.; Nekoubin, H.; Moradlu, A.H. Effect of anaesthesia with clove oil in fish (review). Fish Physiol. Biochem. 2012, 38, 1545–1552. [Google Scholar] [CrossRef]
- Balamurugan, J.; Ajith Kumar, T.T.; Prakash, S.; Meenakumari, B.; Balasundaram, C.; Harikrishnan, R. Clove extract: A potential source for stress free transport of fish. Aquaculture 2016, 454, 171–175. [Google Scholar] [CrossRef]
- Roubach, R.; Gomes, L.C.; Leao Fonseca, F.A.; Val, A.L. Eugenol as an efficacious anaesthetic for tambaqui, Colossoma macropomum (Cuvier). Aquac. Res. 2005, 36, 1056–1061. [Google Scholar] [CrossRef]
- Ke, C.; Liu, Q.; Huang, K.; Mo, M.; Chen, H.; Cheng, B. Effect of Water Temperature on the Depletion of Eugenol in Sea Bass under the Simulated Settings in Handling and Transport. Appl. Sci. 2021, 11, 10882. [Google Scholar] [CrossRef]
- Favero, G.C.; Silva, W.D.S.E.; Boaventura, T.P.; Paes Leme, F.D.O.; Luz, R.K. Eugenol or salt to mitigate stress during the transport of juvenile Lophiosilurus alexandri, a Neotropical carnivorous freshwater catfish. Aquaculture 2019, 512, 734321. [Google Scholar] [CrossRef]
- Becker, A.G.; Cunha, M.A.D.; Garcia, L.D.O.; Zeppenfeld, C.C.; Parodi, T.V.; Maldaner, G.; Morel, A.F.; Baldisserotto, B. Efficacy of eugenol and the methanolic extract of Condalia buxifolia during the transport of the silver catfish Rhamdia quelen. Neotrop. Ichthyol. 2013, 11, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Perry, S.F.; Laurent, P. Environmental effects on fish gill structure and function. In Fish Ecophysiology; Springer: Berlin/Heidelberg, Germany, 1993; pp. 231–264. [Google Scholar]
- Odei, D.K.; Hagen, Ø.; Peruzzi, S.; Falk-Petersen, I.; Fernandes, J.M. Transcriptome sequencing and histology reveal dosage compensation in the liver of triploid pre-smolt Atlantic salmon. Sci. Rep. 2020, 10, 16836. [Google Scholar] [CrossRef] [PubMed]
- Cupp, A.R.; Hartleb, C.F.; Fredricks, K.T.; Gaikowski, M.P. Effectiveness of eugenol sedation to reduce the metabolic rates of cool and warm water fish at high loading densities. Aquac. Res. 2016, 47, 234–242. [Google Scholar] [CrossRef]
- Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wang, Q.; Mei, J.; Cao, J.; Xie, J. Effects of Melissa officinalis L. Essential Oil in Comparison with Anaesthetics on Gill Tissue Damage, Liver Metabolism and Immune Parameters in Sea Bass (Lateolabrax maculatus) during Simulated Live Transport. Biology 2022, 11, 11. [Google Scholar] [CrossRef]
- Jiang, J.; Wu, X.; Zhou, X.; Feng, L.; Liu, Y.; Jiang, W.; Wu, P.; Zhao, Y. Effects of dietary curcumin supplementation on growth performance, intestinal digestive enzyme activities and antioxidant capacity of crucian carp Carassius auratus. Aquaculture 2016, 463, 174–180. [Google Scholar] [CrossRef]
- Zhao, H.; Peng, K.; Wang, G.; Mo, W.; Huang, Y.; Cao, J. Metabolic changes, antioxidant status, immune response and resistance to ammonia stress in juvenile yellow catfish (Pelteobagrus fulvidraco) fed diet supplemented with sodium butyrate. Aquaculture 2021, 536, 736441. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Smith, S. The animal fatty acid synthase: One gene, one polypeptide, seven enzymes. The FASEB journal 1994, 8, 1248–1259. [Google Scholar] [CrossRef]
- Calderon-Dominguez, M.; Sebastian, D.; Fucho, R.; Weber, M.; Mir, J.F.; García-Casarrubios, E.; Obregón, M.J.; Zorzano, A.; Valverde, Á.M.; Serra, D. Carnitine palmitoyltransferase 1 increases lipolysis, UCP1 protein expression and mitochondrial activity in brown adipocytes. PLoS ONE 2016, 11, e159399. [Google Scholar] [CrossRef] [Green Version]
- Kitayama, K.; Yashiro, M.; Morisaki, T.; Miki, Y.; Okuno, T.; Kinoshita, H.; Fukuoka, T.; Kasashima, H.; Masuda, G.; Hasegawa, T. Pyruvate kinase isozyme M2 and glutaminase might be promising molecular targets for the treatment of gastric cancer. Cancer Sci. 2017, 108, 2462–2469. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Li, W.; Shan, H.; Ma, S. Responses of energy homeostasis and lipid metabolism in Penaeus vannamei exposed to ammonia stress. Aquaculture 2021, 544, 737092. [Google Scholar] [CrossRef]
- Tirone, T.A.; Brunicardi, F.C. Overview of glucose regulation. World J. Surg. 2001, 25, 461. [Google Scholar] [CrossRef]
- Cao, J.; Wang, Q.; Qiu, W.; Mei, J.; Xie, J. Transport and Recovery of Turbot (Scophthalmus maximus) Sedated with MS-222 and Eugenol: Effects on Intermediary Metabolism and Osmoregulation. Animals 2021, 11, 2228. [Google Scholar] [CrossRef]
- Axelrod, J.; Reisine, T.D. Stress hormones: Their interaction and regulation. Science 1984, 224, 452–459. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira, C.P.B.; Lemos, C.H.D.P.; Vidal, L.V.O.; Couto, R.D.; Pereira, D.S.P.; Copatti, C.E. Anaesthesia with eugenol in hybrid Amazon catfish (Pseudoplatystoma reticulatum × Leiarius marmoratus) handling: Biochemical and haematological responses. Aquaculture 2019, 501, 255–259. [Google Scholar] [CrossRef]
- Zhang, L.; Yue, H.Y.; Zhang, H.J.; Xu, L.; Wu, S.G.; Yan, H.J.; Gong, Y.S.; Qi, G.H. Transport stress in broilers: I. Blood metabolism, glycolytic potential, and meat quality. Poultry Sci. 2009, 88, 2033–2041. [Google Scholar] [CrossRef]
- Halliwell, B.; Chirico, S. Lipid peroxidation: Its mechanism, measurement, and significance. Am. J. Clin. Nutr. 1993, 57, 715S–725S. [Google Scholar] [CrossRef] [Green Version]
- Mylonas, C.; Kouretas, D. Lipid peroxidation and tissue damage. In Vivo 1999, 13, 295–309. [Google Scholar]
- Shylaja, M.R.; Peter, K.V. The functional role of herbal spices. In Handbook of Herbs and Spices; CRC press: Boca Raton, FL, USA, 2004; Volume 2, pp. 26–45. [Google Scholar]
- Yogalakshmi, B.; Viswanathan, P.; Anuradha, C.V. Investigation of antioxidant, anti-inflammatory and DNA-protective properties of eugenol in thioacetamide-induced liver injury in rats. Toxicology 2010, 268, 204–212. [Google Scholar] [CrossRef]
- Anderson, W.G.; Mckinley, R.S.; Colavecchia, M. The use of clove oil as an anesthetic for rainbow trout and its effects on swimming performance. N. Am. J. Fish. Manag. 1997, 17, 301–307. [Google Scholar] [CrossRef]
- Dutra, F.M.; Rönnau, M.; Sponchiado, D.; Forneck, S.C.; Freire, C.A.; Ballester, E.L.C. Histological alterations in gills of Macrobrachium amazonicum juveniles exposed to ammonia and nitrite. Aquat. Toxicol. 2017, 187, 115–123. [Google Scholar] [CrossRef]
- Zhao, Q.; Feng, K.; Zhang, L.; Bai, Y.; Yao, W. Effects of Acute Ammonia Stress on Antioxidant Responses, Histopathology and Ammonia Detoxification Metabolism in Triangle Sail Mussels (Hyriopsis cumingii). Water 2021, 13, 425. [Google Scholar] [CrossRef]
- Yang, W.; Sun, H.; Xiang, F.; Yang, Z.; Chen, Y. Response of juvenile crucian carp (Carassius auratus) to long-term ammonia exposure: Feeding, growth, and antioxidant defenses. J. Freshw. Ecol. 2011, 26, 563–570. [Google Scholar] [CrossRef]
- Maltez, L.C.; Stringhetta, G.R.; Enamorado, A.D.; Okamoto, M.H.; Romano, L.A.; Monserrat, J.M.; Sampaio, L.A.; Garcia, L. Ammonia exposure and subsequent recovery trigger oxidative stress responses in juveniles of Brazilian flounder Paralichthys orbignyanus. Fish Physiol. Biochem. 2017, 43, 1747–1759. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Abbreviation | Primer Sequence (5′-3′) | Amplicon Size (bp) | Accession Number |
|---|---|---|---|
| CPT-1 a | F: GCAACACGGCAAGATGTCC | 96 | XM_022742255.1 |
| R: GAACCCTGGTAGCTGTAGAGTAGA | |||
| FAS a | F: GGCTATCTGTCGCACTTTCTG | 173 | XM_022765992.1 |
| R: ATTCACGCACTCGCTTCG | |||
| PK a | F: GATTGAGAATGGCGGTATGC | 146 | XM_022756239.1 |
| R: GGATGAAGGAGGCGAAGA | |||
| CAT a | F: CAAGTTTTACACTGAGGAGGGC | 125 | XM_022756059.1 |
| R: TGTGGGTTTGGGGATTGC | |||
| Mn-SOD a | F: CCAGCCTCAGCCAAACTAT | 211 | XM_022737832.1 |
| R: GCGGTCACATCTCCCTTT | |||
| GR a | F: TCACGAGCAGGAAGAGTCAG | 106 | XM_022760278.1 |
| R: TGCAGCATCTCATCACAGC | |||
| Keap1 | F: CCTCCATAAACCCACCAAAG | 203 | XM_022766859.1 |
| R: CAGCGTAGAAAAGCCCACT | |||
| EF1-α b | F: ATCGTTGCCGCTGGTGTT | 134 | XM_022744048.1 |
| R: TCGGTGGAGTCCATCTTGTT | |||
| β-Actin b | F: TCTGGTGGGGCAATGATCTTGATCTT | 212 | XM_022757055.1 |
| R: CCTTCCTTCCTCGGTATGGAGTCC |
| Indexes | Groups | ||||
|---|---|---|---|---|---|
| D0 | D25 | D50 | D75 | D100 | |
| pH | 6.58 ± 0.10 a | 6.74 ± 0.05 a | 6.70 ± 0.14 a | 6.80 ± 0.11 a | 6.78 ± 0.11 a |
| DO (mg/L) | 9.89 ± 0.16 b | 10.58 ± 0.19 b | 7.68 ± 0.24 c | 5.82 ± 0.10 d | 14.75 ± 0.24 a |
| NH4+-N (mg/L) | 0.91 ± 0.04 c | 0.85 ± 0.03 c | 1.30 ± 0.08 b | 2.13 ± 0.12 a | 0.92 ± 0.04 c |
| NO2−-N (mg/L) | 0.01 ± 0.00 c | 0.01 ± 0.00 c | 0.02 ± 0.00 b | 0.04 ± 0.00 a | 0.01 ± 0.00 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Fu, Z.; Dai, S.; Yu, G.; Guo, Y.; Ma, Z. Effects of Eugenol on Water Quality and the Metabolism and Antioxidant Capacity of Juvenile Greater Amberjack (Seriola dumerili) under Simulated Transport Conditions. Animals 2022, 12, 2880. https://doi.org/10.3390/ani12202880
He Y, Fu Z, Dai S, Yu G, Guo Y, Ma Z. Effects of Eugenol on Water Quality and the Metabolism and Antioxidant Capacity of Juvenile Greater Amberjack (Seriola dumerili) under Simulated Transport Conditions. Animals. 2022; 12(20):2880. https://doi.org/10.3390/ani12202880
Chicago/Turabian StyleHe, Yuhang, Zhengyi Fu, Shiming Dai, Gang Yu, Yunfeng Guo, and Zhenhua Ma. 2022. "Effects of Eugenol on Water Quality and the Metabolism and Antioxidant Capacity of Juvenile Greater Amberjack (Seriola dumerili) under Simulated Transport Conditions" Animals 12, no. 20: 2880. https://doi.org/10.3390/ani12202880
APA StyleHe, Y., Fu, Z., Dai, S., Yu, G., Guo, Y., & Ma, Z. (2022). Effects of Eugenol on Water Quality and the Metabolism and Antioxidant Capacity of Juvenile Greater Amberjack (Seriola dumerili) under Simulated Transport Conditions. Animals, 12(20), 2880. https://doi.org/10.3390/ani12202880