
The Importance of Dietary Antioxidants on Oxidative Stress, Meat and Milk Production, and Their Preservative Aspects in Farm Animals: Antioxidant Action, Animal Health, and Product Quality—Invited Review


 , ,
, ,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
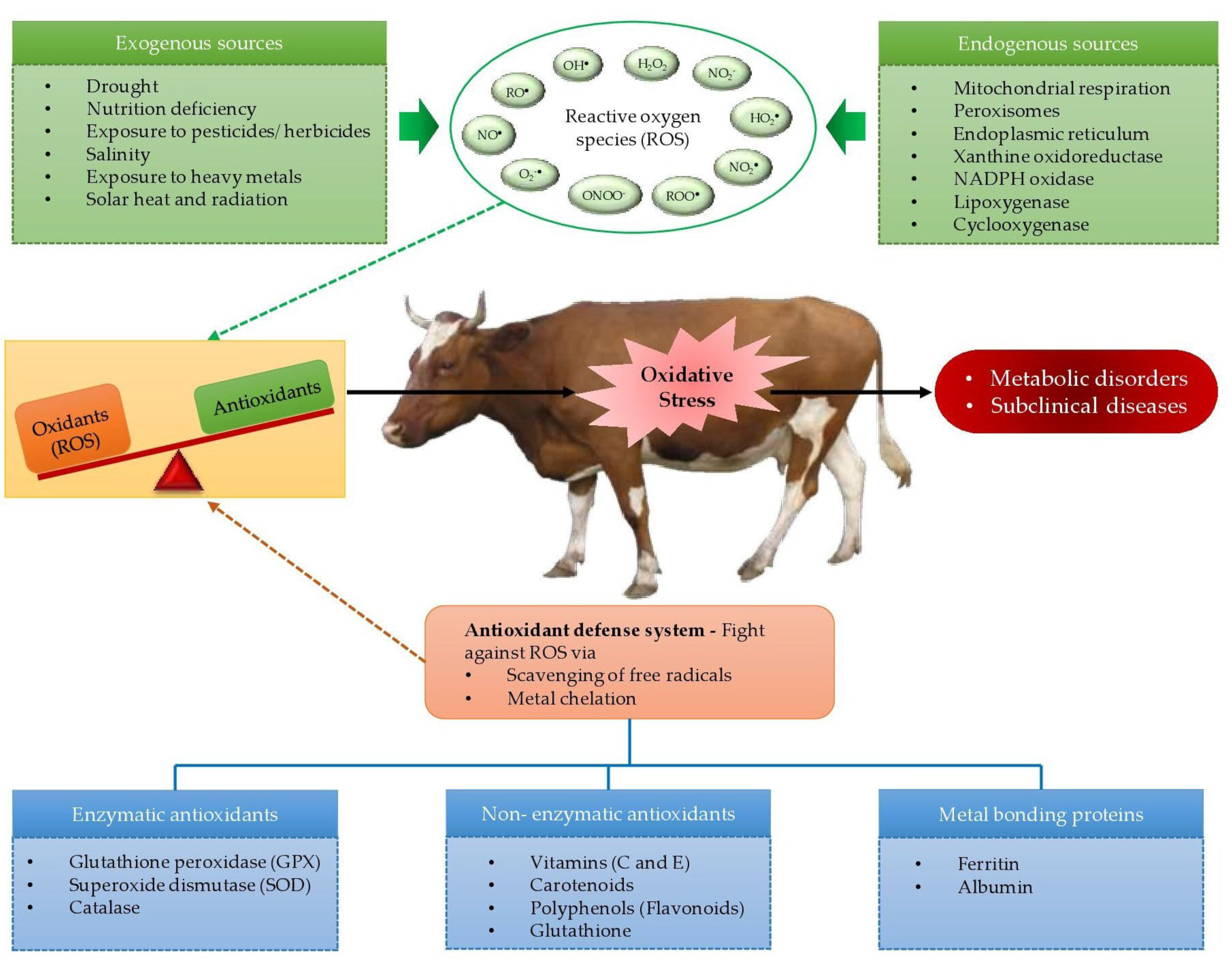
2. Development of Reactive Oxygen Species and Oxidative Stress in Living Organisms
2.1. Development of Reactive Oxygen Species
2.2. Development of Oxidative Stress
2.2.1. Oxidative Stress and Mitochondrial Damage
2.2.2. Oxidative Stress and Antioxidant Defence
3. Sources of Reactive Oxygen Species and Free Radicals in Biological Systems
3.1. Non-Regulated Production of Reactive Oxygen Species
3.1.1. Mitochondrial Respiration
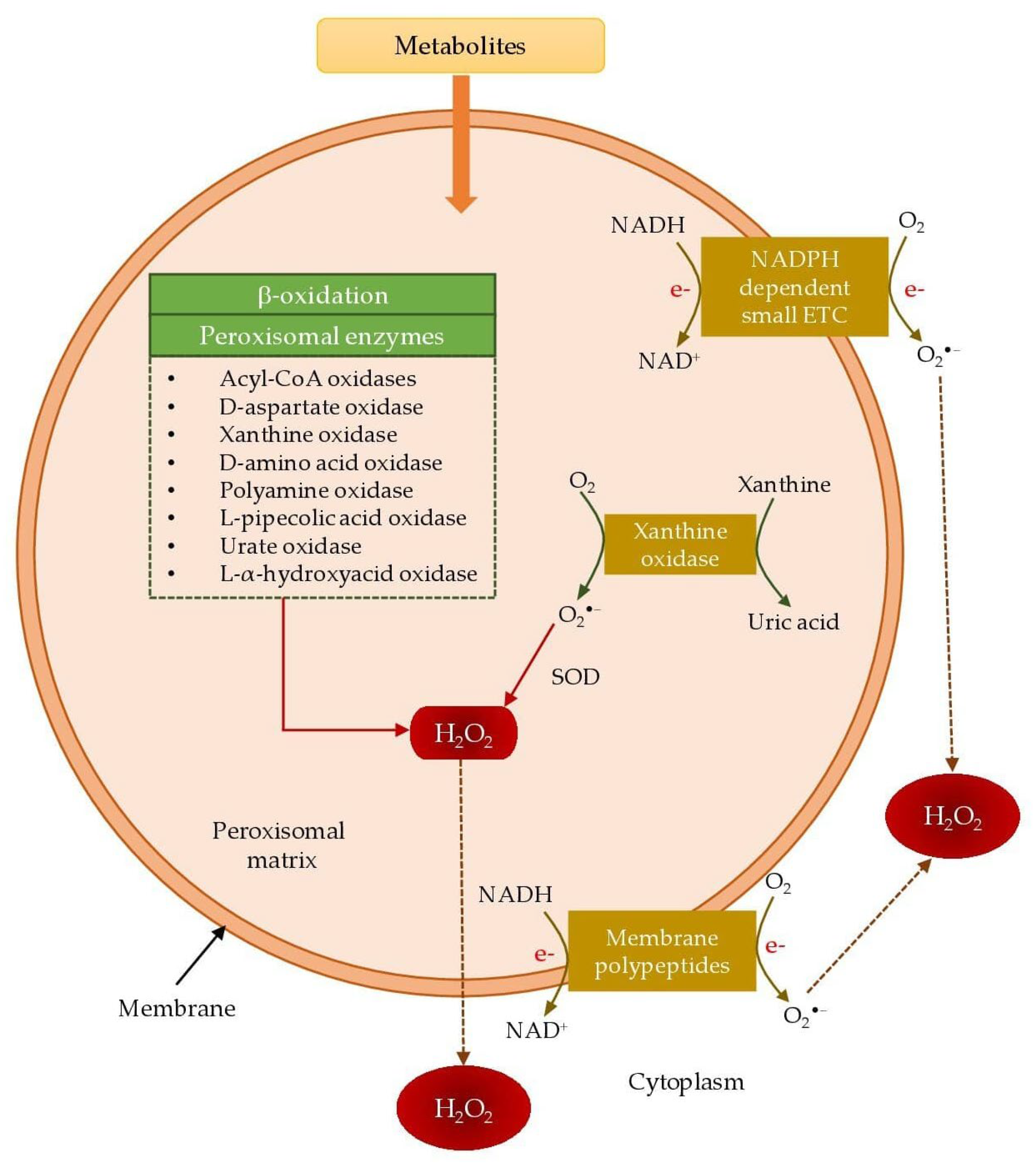
3.1.2. Peroxisomes
3.1.3. Endoplasmic Reticulum
3.1.4. Xanthine Oxidoreductase
3.1.5. Dopamine
3.1.6. Photosensitization Reactions
3.2. Regulated Production of Reactive Oxygen Species
3.2.1. Nitric Oxide Synthase
3.2.2. NADPH Oxidase
3.2.3. Arachidonate Cascade Enzymes
3.2.4. Cyclooxygenase (COX-1)
3.3. Other Cellular ROS Sources
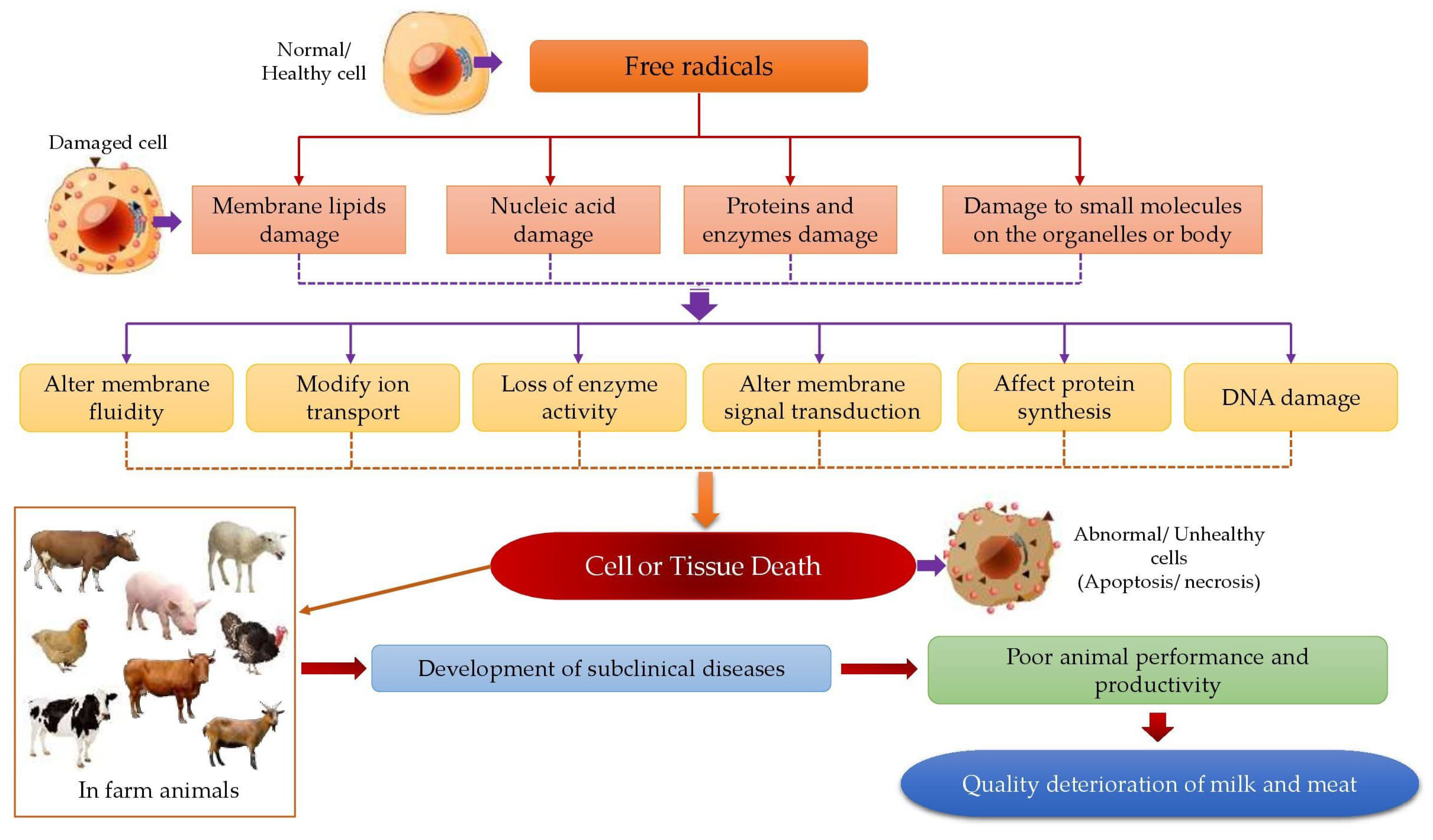
4. The Impact of Radical Formation on the Oxidation of Biological Macromolecules in Cellular System
4.1. The Oxidation of Lipids
Oxidation of Lipids Caused by ROOH
4.2. The Oxidation of Proteins
4.3. The Oxidation of Carbohydrates
4.4. The Oxidation of DNA
5. Actions of Dietary Antioxidants, Endogenous Antioxidants, and Metal Binding Proteins on Free Radical Formation in Biological Systems
5.1. Non-Enzyme Defence Systems
5.1.1. Antioxidant Actions of Vitamins
5.1.2. Antioxidant Actions of Minerals and Trace Elements
5.1.3. Antioxidant Actions of Proteins
5.1.4. Antioxidant Actions of Phytonutrients
5.2. Antioxidant Defence by Metal Binding Proteins
5.2.1. Actions of Iron in Biological Systems
5.2.2. Actions of Copper in Biological Systems
5.3. Enzymatic Defence Systems
5.3.1. Superoxide Dismutase and Superoxide Reductase
5.3.2. Glutathione Peroxidase
5.3.3. Catalase
5.3.4. Heme Oxygenase
5.4. Other Endogenous Antioxidants
5.4.1. Glutathione
5.4.2. Lipoic Acid
5.4.3. Uric Acid
5.4.4. Coenzyme Q10
6. Feeding Systems, Antioxidant Actions and Their Impact on Productivity and Functional Aspects of Meat and Milk in Farm Animals
6.1. The Impact of Dietary Antioxidants on Performance and Productivity of Farm Animals
6.2. The Impact of Dietary Antioxidants on Functional Aspects of Meat from Farm Animals
6.3. The Impact of Dietary Antioxidants on Performance and Milk Productivity of Farm Animals
6.4. The Impact of Dietary Antioxidants on Functional Aspects of Milk and Milk Products from Farm Animals
7. Future Perspectives
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Devine, P.J.; Perreault, S.D.; Luderer, U. Roles of reactive oxygen species and antioxidants in ovarian toxicity. Biol. Reprod. 2012, 86, 27. [Google Scholar] [CrossRef] [PubMed]
- Mutinati, M.; Pantaleo, M.; Roncetti, M.; Piccinno, M.; Rizzo, A.; Sciorsci, R. Oxidative stress in neonatology. A review. Reprod. Domest. Anim. 2014, 49, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, U.; Das, D.; Banerjee, R.K. Reactive oxygen species: Oxidative damage and pathogenesis. Curr. Sci. 1999, 77, 658–666. [Google Scholar]
- Beneš, L.; Ďuračková, Z.; Ferenčik, M. Chemistry, physiology and pathology of free radicals. Life Sci. 1999, 65, 1865–1874. [Google Scholar]
- Lauridsen, C. From oxidative stress to inflammation: Redox balance and immune system. Poult. Sci. 2019, 98, 4240–4246. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Ruiz, A.N.; Aguilar, M.T.H.; Franch, P.C.; Jaén, A.B.L.; Bellés, V.V.; Gallardo Alés, M. C-13. Actividad antioxidante de la leche humana: Relación con factores dietéticos. Pediatría Atención Primaria 2010, 12, e69. [Google Scholar]
- Sretenović, L.; Aleksić, S.; Petrović, M.P.; Miščević, B. Nutritional factors influencing improvement of milk and meat quality as well as productive and reproductive parameters of cattle. Biotechnol. Anim. Husb. 2007, 23, 217–226. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Hopkins, D.L.; Giri, K.; Jacobs, J.L.; Plozza, T.; Lewandowski, P.; Bekhit, A.E.D.A. The use of oxidative stress biomarkers in live animals (in vivo) to predict meat quality deterioration postmortem (in vitro) caused by changes in muscle biochemical components. J. Anim. Sci. 2017, 95, 3012–3024. [Google Scholar]
- Bekhit, A.E.D.A.; Hopkins, D.L.; Fahri, F.T.; Ponnampalam, E.N. Oxidative processes in muscle systems and fresh meat: Sources, markers, and remedies. Compr. Rev. Food Sci. Food Saf. 2013, 12, 565–597. [Google Scholar] [CrossRef]
- Castillo, C.; Pereira, V.; Abuelo, Á.; Hernández, J. Effect of supplementation with antioxidants on the quality of bovine milk and meat production. Sci. World J. 2013, 2013, 616098. [Google Scholar] [CrossRef] [PubMed]
- Salami, S.A.; Guinguina, A.; Agboola, J.O.; Omede, A.A.; Agbonlahor, E.M.; Tayyab, U. In vivo and postmortem effects of feed antioxidants in livestock: A review of the implications on authorization of antioxidant feed additives. Animal 2016, 10, 1375–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Ju, N.; Chang, J.; Ge, R.; Zhao, Y.; Zhang, G. Dietary α-lipoic acid supplementation improves postmortem color stability of the lamb muscles through changing muscle fiber types and antioxidative status. Meat Sci. 2022, 194, 108945. [Google Scholar] [CrossRef]
- Arnold, R.N.; Scheller, K.K.; Arp, S.C.; Williams, S.N.; Schaefer, D.M. Dietary α-tocopheryl acetate enhances beef quality in Holstein and beef breed steers. J. Food Sci. 1993, 58, 28–33. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Butler, K.L.; McDonagh, M.B.; Jacobs, J.L.; Hopkins, D.L. Relationship between muscle antioxidant status, forms of iron, polyunsaturated fatty acids and functionality (retail colour) of meat in lambs. Meat Sci. 2012, 90, 297–303. [Google Scholar] [CrossRef]
- Jensen, C.; Lauridsen, C.; Bertelsen, G. Dietary vitamin E: Quality and storage stability of pork and poultry. Trends Food Sci. Technol. 1998, 9, 62–72. [Google Scholar] [CrossRef]
- Karre, L.; Lopez, K.; Getty, K.J.K. Natural antioxidants in meat and poultry products. Meat Sci. 2013, 94, 220–227. [Google Scholar] [CrossRef]
- Nassu, R.T.; Uttaro, B.; Aalhus, J.L.; Zawadski, S.; Juárez, M.; Dugan, M.E.R. Type of packaging affects the colour stability of vitamin E enriched beef. Food Chem. 2012, 135, 1868–1872. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2015, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Krumova, K.; Cosa, G. Overview of reactive oxygen species. In Singlet Oxygen: Applications in Biosciences and Nanosciences; Nobell, S., Flors, C., Eds.; Royal Society of Chemistry: London, UK, 2016; Volume 1, pp. 1–21. [Google Scholar]
- Kafantaris, I.; Kotsampasi, B.; Christodoulou, V.; Makri, S.; Stagos, D.; Gerasopoulos, K.; Petrotos, K.; Goulas, P.; Kouretas, D. Effects of dietary grape pomace supplementation on performance, carcass traits and meat quality of lambs. In Vivo 2018, 32, 807–812. [Google Scholar] [CrossRef] [Green Version]
- Mutinati, M.; Piccinno, M.; Roncetti, M.; Campanile, D.; Rizzo, A.; Sciorsci, R.L. Oxidative stress during pregnancy in the sheep. Reprod. Domest. Anim. 2013, 48, 353–357. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masters, C.J. Cellular signalling: The role of the peroxisome. Cell. Signal. 1996, 8, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.; Hancock, J.T.; Bright, J.; Harrison, J.; Weir, I.; Hooley, R.; Neill, S.J. A role for ETR1 in hydrogen peroxide signaling in stomatal guard cells. Plant Physiol. 2005, 137, 831–834. [Google Scholar] [CrossRef] [PubMed]
- Covarrubias, L.; Hernández-García, D.; Schnabel, D.; Salas-Vidal, E.; Castro-Obregón, S. Function of reactive oxygen species during animal development: Passive or active? Dev. Biol. 2008, 320, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, I.S. Oxidative stress and vascular disease: Insights from isoprostane measurement. Clin. Chem. 2005, 51, 14–15. [Google Scholar] [CrossRef] [Green Version]
- Beaudeux, J.L.; Vasson, M.P. Sources cellulaires des espèces réactives de l’oxygène. In Radicaux Libres et Stress Oxydant; Delattre, J., Beaudeux, J.L., Bonnfort-Rousselot, D., Eds.; Lavoisier: Paris, France, 2005; pp. 45–86. [Google Scholar]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Vossen, E.; Ntawubizi, M.; Raes, K.; Smet, K.; Huyghebaert, G.; Arnouts, S.; De Smet, S. Effect of dietary antioxidant supplementation on the oxidative status of plasma in broilers. J. Anim. Physiol. Anim. Nutr. 2011, 95, 198–205. [Google Scholar] [CrossRef]
- Holman, B.W.B.; Baldi, G.; Chauhan, S.S.; Hopkins, D.L.; Seymour, G.R.; Dunshea, F.R.; Collins, D.; Ponnampalam, E.N. Comparison of grain-based diet supplemented with synthetic vitamin E and lucerne hay-based diet on blood oxidative stress biomarkers and lamb meat quality. Small Rumin. Res. 2019, 177, 146–152. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Celi, P.; Ponnampalam, E.N.; Leury, B.J.; Liu, F.; Dunshea, F.R. Antioxidant dynamics in the live animal and implications for ruminant health and product (meat/milk) quality: Role of vitamin E and selenium. Anim. Prod. Sci. 2014, 54, 1525–1536. [Google Scholar] [CrossRef]
- Halliwell, B. Lipid peroxidation, antioxidants and cardiovascular disease: How should we move forward? Cardiovasc. Res. 2000, 47, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Ponnampalam, E.N.; Vahid, V.; Giri, K.; Lewandowski, P.; Jacobs, J.L.; Dunshea, F.R. Muscle antioxidant enzymes activity and gene expression are altered by diet-induced increase in muscle essential fatty acid (α-linolenic acid) concentration in sheep used as a model. Nutrients 2019, 11, 723. [Google Scholar] [CrossRef] [Green Version]
- Yakes, F.M.; Van Houten, B. Mitochondrial DNA damage is more extensive and persists longer than nuclear DNA damage in human cells following oxidative stress. Proc. Natl. Acad. Sci. USA 1997, 94, 514–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreyev, A.Y.; Kushnareva, Y.E.; Starkov, A.A. Mitochondrial metabolism of reactive oxygen species. Biochemistry 2005, 70, 200–214. [Google Scholar] [CrossRef]
- Ježek, P.; Hlavatá, L. Mitochondria in homeostasis of reactive oxygen species in cell, tissues, and organism. Int. J. Biochem. Cell Biol. 2005, 37, 2478–2503. [Google Scholar] [CrossRef] [PubMed]
- Sickmann, A.; Reinders, J.; Wagner, Y.; Joppich, C.; Zahedi, R.; Meyer, H.E.; Schönfisch, B.; Perschil, I.; Chacinska, A.; Guiard, B. The proteome of Saccharomyces cerevisiae mitochondria. Proc. Natl. Acad. Sci. USA 2003, 100, 13207–13212. [Google Scholar] [CrossRef] [Green Version]
- Csiszar, A.; Labinskyy, N.; Perez, V.; Recchia, F.A.; Podlutsky, A.; Mukhopadhyay, P.; Losonczy, G.; Pacher, P.; Austad, S.N.; Bartke, A. Endothelial function and vascular oxidative stress in long-lived GH/IGF-deficient Ames dwarf mice. Am. J. Physiol. -Heart Circ. Physiol. 2008, 295, H1882–H1894. [Google Scholar] [CrossRef] [Green Version]
- Puche, J.E.; García-Fernández, M.; Muntane, J.; Rioja, J.; Gonzalez-Baron, S.; Cortazar, I.C. Low doses of insulin-like growth factor-I induce mitochondrial protection in aging rats. Endocrinology 2008, 149, 2620–2627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbarian, A.; Michiels, J.; Degroote, J.; Majdeddin, M.; Golian, A.; De Smet, S. Association between heat stress and oxidative stress in poultry; mitochondrial dysfunction and dietary interventions with phytochemicals. J. Anim. Sci. Biotechnol. 2016, 7, 37. [Google Scholar] [CrossRef] [Green Version]
- Montilla, S.I.R.; Johnson, T.P.; Pearce, S.C.; Gardan-Salmon, D.; Gabler, N.K.; Ross, J.W.; Rhoads, R.P.; Baumgard, L.H.; Lonergan, S.M.; Selsby, J.T. Heat stress causes oxidative stress but not inflammatory signaling in porcine skeletal muscle. Temperature 2014, 1, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Amadori, M.; Zanotti, C. Immunoprophylaxis in intensive farming systems: The way forward. Vet. Immunol. Immunopathol. 2016, 181, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Baldi, G.; Chauhan, S.S.; Linden, N.; Dunshea, F.R.; Hopkins, D.L.; Rossi, C.A.S.; Dell’Orto, V.; Ponnampalam, E.N. Comparison of a grain-based diet supplemented with synthetic vitamin E versus a lucerne (alfalfa) hay-based diet fed to lambs in terms of carcass traits, muscle vitamin E, fatty acid content, lipid oxidation, and retail colour of meat. Meat Sci. 2019, 148, 105–112. [Google Scholar] [CrossRef]
- Lauridsen, C.; Matte, J.J.; Lessard, M.; Celi, P.; Litta, G. Role of vitamins for gastro-intestinal functionality and health of pigs. Anim. Feed. Sci. Technol. 2021, 273, 114823. [Google Scholar] [CrossRef]
- Velasco, V.; Williams, P. Improving meat quality through natural antioxidants. Chil. J. Agric. Res. 2011, 71, 313. [Google Scholar] [CrossRef]
- Dunshea, F.R.; Gonzalez-Rivas, P.A.; Hung, A.T.; DiGiacomo, K.; Chauhan, S.S.; Leury, B.J.; Celi, P.P.; Ponnampalam, E.N.; Cottrell, J.J. Nutritional strategies to alleviate heat stress in sheep. In Sheep Production Adapting to Climate Change; Sejian, V., Bhatta, R., Gaughan, J., Malik, P.K., Naqvi, S.M.K., Lal, R., Eds.; Springer: Singapore, 2017; pp. 371–388. [Google Scholar]
- De Brito, G.F.; Ponnampalam, E.N.; Hopkins, D.L. The effect of extensive feeding systems on growth rate, carcass traits, and meat quality of finishing lambs. Compr. Rev. Food Sci. Food Saf. 2017, 16, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [Green Version]
- Radzikowska, U.; Rinaldi, A.O.; Sözener, Z.Ç.; Karaguzel, D.; Wojcik, M.; Cypryk, K.; Akdis, M.; Akdis, C.A.; Sokolowska, M. The influence of dietary fatty acids on immune responses. Nutrients 2019, 11, 2990. [Google Scholar] [CrossRef] [Green Version]
- Casas, A.I.; Dao, V.T.V.; Daiber, A.; Maghzal, G.J.; Di Lisa, F.; Kaludercic, N.; Leach, S.; Cuadrado, A.; Jaquet, V.; Seredenina, T. Reactive oxygen-related diseases: Therapeutic targets and emerging clinical indications. Antioxid. Redox Signal. 2015, 23, 1171–1185. [Google Scholar] [CrossRef] [Green Version]
- Casas, A.I.; Nogales, C.; Mucke, H.A.M.; Petraina, A.; Cuadrado, A.; Rojo, A.I.; Ghezzi, P.; Jaquet, V.; Augsburger, F.; Dufrasne, F. On the clinical pharmacology of reactive oxygen species. Pharmacol. Rev. 2020, 72, 801–828. [Google Scholar] [CrossRef]
- Brand, M.D. The sites and topology of mitochondrial superoxide production. Exp. Gerontol. 2010, 45, 466–472. [Google Scholar] [CrossRef] [Green Version]
- Castro, L.; Tórtora, V.; Mansilla, S.; Radi, R. Aconitases: Non-redox iron–sulfur proteins sensitive to reactive species. Acc. Chem. Res. 2019, 52, 2609–2619. [Google Scholar] [CrossRef] [PubMed]
- Veith, A.; Moorthy, B. Role of cytochrome P450s in the generation and metabolism of reactive oxygen species. Curr. Opin. Toxicol. 2018, 7, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Yasui, H.; Hayashi, S.; Sakurai, H. Possible involvement of singlet oxygen species as multiple oxidants in p450 catalytic reactions. Drug Metab. Pharmacokinet. 2005, 20, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puntarulo, S.; Cederbaum, A.I. Production of reactive oxygen species by microsomes enriched in specific human cytochrome P450 enzymes. Free Radic. Biol. Med. 1998, 24, 1324–1330. [Google Scholar] [CrossRef]
- Fransen, M.; Nordgren, M.; Wang, B.; Apanasets, O. Role of peroxisomes in ROS/RNS-metabolism: Implications for human disease. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2012, 1822, 1363–1373. [Google Scholar] [CrossRef] [Green Version]
- Nishino, T.; Okamoto, K.; Eger, B.T.; Pai, E.F.; Nishino, T. Mammalian xanthine oxidoreductase–mechanism of transition from xanthine dehydrogenase to xanthine oxidase. FEBS J. 2008, 275, 3278–3289. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S. Mammalian lipoxygenases: Molecular structures and functions. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1992, 1128, 117–131. [Google Scholar] [CrossRef]
- Bonizzi, G.; Piette, J.; Merville, M.P.; Bours, V. Cell type-specific role for reactive oxygen species in nuclear factor-kappaB activation by interleukin-1. Biochem. Pharmacol. 2000, 59, 7–11. [Google Scholar] [CrossRef]
- Los, M.; Schenk, H.; Hexel, K.; Baeuerle, P.A.; Dröge, W.; Schulze-Osthoff, K. IL-2 gene expression and NF-kappa B activation through CD28 requires reactive oxygen production by 5-lipoxygenase. EMBO J. 1995, 14, 3731–3740. [Google Scholar] [CrossRef] [Green Version]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2000, 279, L1005–L1028. [Google Scholar] [CrossRef] [Green Version]
- Dubois, R.N.; Abramson, S.B.; Crofford, L.; Gupta, R.A.; Simon, L.S.; Van De Putte, L.B.A.; Lipsky, P.E. Cyclooxygenase in biology and disease. FASEB J. 1998, 12, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.M.; Kim, J.Y.; Kim, J.H. Cytosolic phospholipase A 2, lipoxygenase metabolites, and reactive oxygen species. BMB Rep. 2008, 41, 555–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 83, 47–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Blake, D.R.; Stevens, C.R.; Kanuler, J.M.; Ward, P.G.; Symons, M.C.R.; Benboubetra, M.; Harrison, R. A reappraisal of xanthine dehydrogenase and oxidase in hypoxic reperfusion injury: The role of NADH as an electron donor. Free Radic. Res. 1998, 28, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.W.; Stamler, J.S. New insights into protein S-nitrosylation: Mitochondria as a model system. J. Biol. Chem. 2004, 279, 25891–25897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, T.M.; Stevens, C.R.; Benjamin, N.; Eisenthal, R.; Harrison, R.; Blake, D.R. Xanthine oxidoreductase catalyses the reduction of nitrates and nitrite to nitric oxide under hypoxic conditions. FEBS Lett. 1998, 427, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Biosa, A.; Arduini, I.; Soriano, M.E.; Bubacco, L.; Bisaglia, M. Analysis of the functional effects mediated by dopamine oxidation products at the mitochondrial level. J. Neurochem. 2017, 142, 106. [Google Scholar]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef]
- Weng, M.; Xie, X.; Liu, C.; Lim, K.L.; Zhang, C.W.; Li, L. The sources of reactive oxygen species and its possible role in the pathogenesis of Parkinson’s disease. Park. Dis. 2018, 2018, 9163040. [Google Scholar] [CrossRef] [Green Version]
- Hidiroglou, N.; Batra, T.R. Plasma and tissue concentrations of vitamin E following supplementation of two forms of vitamin E in sheep. Small Rumin. Res. 1996, 21, 83–87. [Google Scholar] [CrossRef]
- Skřivanová, E.; Marounek, M.; De Smet, S.; Raes, K. Influence of dietary selenium and vitamin E on quality of veal. Meat Sci. 2007, 76, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, R.; Ranjan, A.; Dhaliwal, G.S.; Patra, R.C. L-Ascorbic acid (vitamin C) supplementation to optimize health and reproduction in cattle. Vet. Q. 2012, 32, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Higaki, T.; Matsubara, Y.; Nawa, A. Nitric oxide and reactive oxygen species in the pathogenesis of preeclampsia. Int. J. Mol. Sci. 2015, 16, 4600–4614. [Google Scholar] [CrossRef] [PubMed]
- Ashoori, M.; Saedisomeolia, A. Riboflavin (vitamin B2) and oxidative stress: A review. Br. J. Nutr. 2014, 111, 1985–1991. [Google Scholar] [CrossRef] [Green Version]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- Van der Vliet, A.; Danyal, K.; Heppner, D.E. Dual oxidase: A novel therapeutic target in allergic disease. Br. J. Pharmacol. 2018, 175, 1401–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumimoto, H. Structure, regulation and evolution of Nox-family NADPH oxidases that produce reactive oxygen species. FEBS J. 2008, 275, 3249–3277. [Google Scholar] [CrossRef]
- Schramm, A.; Matusik, P.; Osmenda, G.; Guzik, T.J. Targeting NADPH oxidases in vascular pharmacology. Vasc. Pharmacol. 2012, 56, 216–231. [Google Scholar] [CrossRef] [Green Version]
- Chew, B.P. Antioxidant vitamins affect food animal immunity and health. J. Nutr. 1995, 125, 1804–1808. [Google Scholar]
- Gutteridge, J.M.C. Lipid peroxidation initiated by superoxide-dependent hydroxyl radicals using complexed iron and hydrogen peroxide. FEBS Lett. 1984, 172, 245–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutteridge, J.M. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin. Chem. 1995, 41, 1819–1828. [Google Scholar] [CrossRef]
- Bogdan, C.; Röllinghoff, M.; Diefenbach, A. Reactive oxygen and reactive nitrogen intermediates in innate and specific immunity. Curr. Opin. Immunol. 2000, 12, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, S.M.; Lee, R.T. Thioredoxin and thioredoxin target proteins: From molecular mechanisms to functional significance. Antioxid. Redox Signal. 2013, 18, 1165–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marnett, L.J. Lipid peroxidation—DNA damage by malondialdehyde. Mutat. Res./Fundam. Mol. Mech. Mutagen. 1999, 424, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Králová, M. The effect of lipid oxidation on the quality of meat and meat products. Maso Int. J. Food Sci. Technol. 2015, 2, 125–132. [Google Scholar]
- Poli, G.; Leonarduzzi, G.; Biasi, F.; Chiarpotto, E. Oxidative stress and cell signalling. Curr. Med. Chem. 2004, 11, 1163–1182. [Google Scholar] [CrossRef] [PubMed]
- Pappas, A.C.; Zoidis, E.; Surai, P.; Zervas, G. Selenoproteins and maternal nutrition. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 151, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Roche, M.; Rondeau, P.; Singh, N.R.; Tarnus, E.; Bourdon, E. The antioxidant properties of serum albumin. FEBS Lett. 2008, 582, 1783–1787. [Google Scholar] [CrossRef] [PubMed]
- White, K.N.; Conesa, C.; Sánchez, L.; Amini, M.; Farnaud, S.; Lorvoralak, C.; Evans, R.W. The transfer of iron between ceruloplasmin and transferrins. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Baltaci, A.K.; Yuce, K.; Mogulkoc, R. Zinc metabolism and metallothioneins. Biol. Trace Elem. Res. 2018, 183, 22–31. [Google Scholar] [CrossRef]
- Krężel, A.; Maret, W. The functions of metamorphic metallothioneins in zinc and copper metabolism. Int. J. Mol. Sci. 2017, 18, 1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaer, D.J.; Vinchi, F.; Ingoglia, G.; Tolosano, E.; Buehler, P.W. Haptoglobin, hemopexin, and related defense pathways—Basic science, clinical perspectives, and drug development. Front. Physiol. 2014, 5, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, M.; Hernandez-Mijares, A.; Garcia-Malpartida, K.; Banuls, C.; Bellod, L.; Victor, M.V. Mitochondria-targeted antioxidant peptides. Curr. Pharm. Des. 2010, 16, 3124–3131. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.S. Natural antioxidants: Sources, compounds, mechanisms of action, and potential applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant phytochemicals for the prevention and treatment of chronic diseases. Molecules 2015, 20, 21138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havsteen, B. Flavonoids, a class of natural products of high pharmacological potency. Biochem. Pharmacol. 1983, 32, 1141–1148. [Google Scholar] [CrossRef]
- Kim, H.P.; Son, K.H.; Chang, H.W.; Kang, S.S. Anti-inflammatory plant flavonoids and cellular action mechanisms. J. Pharmacol. Sci. 2004, 96, 229–245. [Google Scholar] [CrossRef]
- García-Lafuente, A.; Guillamón, E.; Villares, A.; Rostagno, M.A.; Martínez, J.A. Flavonoids as anti-inflammatory agents: Implications in cancer and cardiovascular disease. Inflamm. Res. 2009, 58, 537–552. [Google Scholar] [CrossRef]
- Mahfuz, S.; Shang, Q.; Piao, X. Phenolic compounds as natural feed additives in poultry and swine diets: A review. J. Anim. Sci. Biotechnol. 2021, 12, 48. [Google Scholar] [CrossRef]
- Brenes, A.; Viveros, A.; Chamorro, S.; Arija, I. Use of polyphenol-rich grape by-products in monogastric nutrition. A review. Anim. Feed. Sci. Technol. 2016, 211, 1–17. [Google Scholar] [CrossRef]
- Roila, R.; Valiani, A.; Miraglia, D.; Ranucci, D.; Forte, C.; Trabalza-Marinucci, M.; Servili, M.; Codini, M.; Branciari, R. Olive mill wastewater phenolic concentrate as natural antioxidant against lipid-protein oxidative deterioration in chicken meat during storage. Ital. J. Food Saf. 2018, 7, 7342. [Google Scholar] [CrossRef]
- Surai, P.F. Polyphenol compounds in the chicken/animal diet: From the past to the future. J. Anim. Physiol. Anim. Nutr. 2014, 98, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Antoniussen, C.S.; Rasmussen, H.H.; Holst, M.; Lauridsen, C. Reducing disease activity of inflammatory bowel disease by consumption of plant-based foods and nutrients. Front. Nutr. 2021, 8, 733433. [Google Scholar] [CrossRef]
- Santhiravel, S.; Bekhit, A.E.D.A.; Mendis, E.; Jacobs, J.L.; Dunshea, F.R.; Rajapakse, N.; Ponnampalam, E.N. The impact of plant phytochemicals on the gut microbiota of humans for a balanced life. Int. J. Mol. Sci. 2022, 23, 8124. [Google Scholar] [CrossRef] [PubMed]
- Okarter, N.; Liu, R.H. Health benefits of whole grain phytochemicals. Crit. Rev. Food Sci. Nutr. 2010, 50, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.C.; Bailey, R.L.; Blumberg, J.B.; Burton-Freeman, B.; Chen, C.O.; Crowe-White, K.M.; Drewnowski, A.; Hooshmand, S.; Johnson, E.; Lewis, R. Fruits, vegetables, and health: A comprehensive narrative, umbrella review of the science and recommendations for enhanced public policy to improve intake. Crit. Rev. Food Sci. Nutr. 2020, 60, 2174–2211. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Sang, S. Phytochemicals in whole grain wheat and their health-promoting effects. Mol. Nutr. Food Res. 2017, 61, 1600852. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Norng, S.; Burnett, V.F.; Dunshea, F.R.; Jacobs, J.L.; Hopkins, D.L. The synergism of biochemical components controlling lipid oxidation in lamb muscle. Lipids 2014, 49, 757–766. [Google Scholar] [CrossRef]
- Liu, Q.; Lanari, M.C.; Schaefer, D.M. A review of dietary vitamin E supplementation for improvement of beef quality. J. Anim. Sci. 1995, 73, 3131–3140. [Google Scholar] [CrossRef]
- O’Grady, M.N.; Monahan, F.J.; Fallon, R.J.; Allen, P. Effects of dietary supplementation with vitamin E and organic selenium on the oxidative stability of beef. J. Anim. Sci. 2001, 79, 2827–2834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellés, M.; del Mar Campo, M.; Roncalés, P.; Beltrán, J.A. Supranutritional doses of vitamin E to improve lamb meat quality. Meat Sci. 2019, 149, 14–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arshad, M.A.; Ebeid, H.M.; Hassan, F. Revisiting the effects of different dietary sources of selenium on the health and performance of dairy animals: A review. Biol. Trace Elem. Res. 2021, 199, 3319–3337. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Maya, M.A.; Domínguez-Vara, I.A.; Trujillo-Gutiérrez, D.; Morales-Almaráz, E.; Sánchez-Torres, J.E.; Bórquez-Gastelum, J.L.; Acosta-Dibarrat, J.; Grageola-Nuñez, F.; Rodríguez-Carpena, J.G. Growth performance parameters, carcass traits, and meat quality of lambs supplemented with zinc methionine and (or) zinc oxide in feedlot system. Can. J. Anim. Sci. 2019, 99, 585–595. [Google Scholar] [CrossRef]
- Hashem, N.M.; Abu-Tor, E.M.; Abo-Elezz, Z.R.; Latif, M.G.A. Relevance of antioxidant vitamin supplementation for improvement of milk production, milk quality and energy status of lactating ewes. Small Rumin. Res. 2019, 177, 153–159. [Google Scholar] [CrossRef]
- Maiorano, G.; Angwech, H.; Di Memmo, D.; Wilkanowska, A.; Mucci, R.; Abiuso, C.; Tavaniello, S. Effects of intramuscular vitamin E multiple injection on quality, oxidative stability and consumer acceptability of meat from Laticauda lambs fed under natural rearing conditions. Small Rumin. Res. 2016, 139, 52–59. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Muir, J.P.; Naumann, H.D.; Norris, A.B.; Ramírez-Restrepo, C.A.; Mertens-Talcott, S.U. Nutritional aspects of ecologically relevant phytochemicals in ruminant production. Front. Vet. Sci. 2021, 8, 628445. [Google Scholar] [CrossRef]
- Moñino, I.; Martínez, C.; Sotomayor, J.A.; Lafuente, A.; Jordán, M.J. Polyphenolic transmission to segureño lamb meat from ewes’ diet supplemented with the distillate from rosemary (Rosmarinus officinalis) leaves. J. Agric. Food Chem. 2008, 56, 3363–3367. [Google Scholar] [CrossRef]
- Simitzis, P.E.; Deligeorgis, S.G.; Bizelis, J.A.; Dardamani, A.; Theodosiou, I.; Fegeros, K. Effect of dietary oregano oil supplementation on lamb meat characteristics. Meat Sci. 2008, 79, 217–223. [Google Scholar] [CrossRef]
- Gobert, M.; Gruffat, D.; Habeanu, M.; Parafita, E.; Bauchart, D.; Durand, D. Plant extracts combined with vitamin E in PUFA-rich diets of cull cows protect processed beef against lipid oxidation. Meat Sci. 2010, 85, 676–683. [Google Scholar] [CrossRef]
- Delosière, M.; Durand, D.; Bourguet, C.; Terlouw, E.M.C. Lipid oxidation, pre-slaughter animal stress and meat packaging: Can dietary supplementation of vitamin E and plant extracts come to the rescue? Food Chem. 2020, 309, 125668. [Google Scholar] [CrossRef]
- Noziere, P.; Graulet, B.; Lucas, A.; Martin, B.; Grolier, P.; Doreau, M. Carotenoids for ruminants: From forages to dairy products. Anim. Feed. Sci. Technol. 2006, 131, 418–450. [Google Scholar] [CrossRef]
- Wang, X.D. Lycopene metabolism and its biological significance. Am. J. Clin. Nutr. 2012, 96, 1214–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Xu, C.; Qu, Y.; Hopkins, D.L.; Liu, C.; Wang, B.; Gao, Y.; Luo, H. Dietary lycopene powder improves meat oxidative stability in Hu lambs. J. Sci. Food Agric. 2019, 99, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Graf, E.; Eaton, J.W. Antioxidant functions of phytic acid. Free Radic. Biol. Med. 1990, 8, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, J.; Ward, D.M. The essential nature of iron usage and regulation. Curr. Biol. 2013, 23, 642–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaňhara, J.; Fajt, Z.; Blahová, J.; Tykačová, V.; Píšťková, K.; Hostovský, M.; Kazatelová, Z.; Vašek, J.; Svoboda, A. The effects of iron deficiency on the level of oxidative stress and antioxidant defenses in suckling piglets. Acta Vet. 2017, 86, 365–371. [Google Scholar] [CrossRef]
- Surai, P.F.; Fisinin, V.I. Selenium in sow nutrition. Anim. Feed. Sci. Technol. 2016, 211, 18–30. [Google Scholar] [CrossRef]
- Kim, B.E.; Nevitt, T.; Thiele, D.J. Mechanisms for copper acquisition, distribution and regulation. Nat. Chem. Biol. 2008, 4, 176–185. [Google Scholar] [CrossRef]
- Lutsenko, S. Copper trafficking to the secretory pathway. Metallomics 2016, 8, 840–852. [Google Scholar] [CrossRef] [Green Version]
- Meyer, L.A.; Durley, A.P.; Prohaska, J.R.; Harris, Z.L. Copper transport and metabolism are normal in aceruloplasminemic mice. J. Biol. Chem. 2001, 276, 36857–36861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linder, M.C. Ceruloplasmin and other copper binding components of blood plasma and their functions: An update. Metallomics 2016, 8, 887–905. [Google Scholar] [CrossRef] [PubMed]
- Nazıroğlu, M. New molecular mechanisms on the activation of TRPM2 channels by oxidative stress and ADP-ribose. Neurochem. Res. 2007, 32, 1990–2001. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.F.; Teixeira, M.; Valentine, J.S. Superoxide dismutases and superoxide reductases. Chem. Rev. 2014, 114, 3854–3918. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2013, 1830, 3289–3303. [Google Scholar] [CrossRef]
- Barañano, D.E.; Rao, M.; Ferris, C.D.; Snyder, S.H. Biliverdin reductase: A major physiologic cytoprotectant. Proc. Natl. Acad. Sci. USA 2002, 99, 16093–16098. [Google Scholar] [CrossRef] [Green Version]
- Pizzorno, J. Glutathione! Integr. Med. A Clin. J. 2014, 13, 8–12. [Google Scholar]
- Minich, D.M.; Brown, B.I. A review of dietary (phyto) nutrients for glutathione support. Nutrients 2019, 11, 2073. [Google Scholar] [CrossRef]
- Vongsawasdi, P.; Noomhorm, A. Bioactive compounds in meat and their functions. In Functional Foods and Dietary Supplements: Processing Effects and Health Benefits; Noomhorm, A., Ahmad, I., Anal, A.K., Eds.; John Wiley & Sons Ltd: West Sussex, UK, 2014; pp. 113–138. [Google Scholar]
- Sohaib, M.; Anjum, F.M.; Nasir, M.; Saeed, F.; Arshad, M.S.; Hussain, S. Alpha-lipoic acid: An inimitable feed supplement for poultry nutrition. J. Anim. Physiol. Anim. Nutr. 2018, 102, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.B.; Olson, K.C.; Meyer, D.L.; Brandt, M.M.; Rentfrow, G.K.; Stahl, C.A.; Berg, E.P. Effects of lipoic acid supplementation on finishing steer growth performance, carcass merit, beef tenderness, and beef retail display properties. Prof. Anim. Sci. 2005, 21, 480–485. [Google Scholar] [CrossRef]
- Bhagavan, H.N.; Chopra, R.K. Coenzyme Q10: Absorption, tissue uptake, metabolism and pharmacokinetics. Free Radic. Res. 2006, 40, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Pravst, I.; Žmitek, K.; Žmitek, J. Coenzyme Q10 contents in foods and fortification strategies. Crit. Rev. Food Sci. Nutr. 2010, 50, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Crane, F.L. Biochemical functions of coenzyme Q10. J. Am. Coll. Nutr. 2001, 20, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Bacou, E.; Walk, C.; Rider, S.; Litta, G.; Perez-Calvo, E. Dietary oxidative distress: A review of nutritional challenges as models for poultry, swine and fish. Antioxidants 2021, 10, 525. [Google Scholar] [CrossRef] [PubMed]
- Nieto, G.; Ros, G. Modification of fatty acid composition in meat through diet: Effect on lipid peroxidation and relationship to nutritional quality—A review. In Lipid Peroxidation; Catala, A., Ed.; IntechOpen: London, UK, 2012; Volume 12, pp. 239–258. [Google Scholar]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.R.; Warner, R.D.; Dunshea, F.R. A dietary sugarcane-derived polyphenol mix reduces the negative effects of cyclic heat exposure on growth performance, blood gas status, and meat quality in broiler chickens. Animals 2020, 10, 1158. [Google Scholar] [CrossRef]
- Iqbal, Y.; Ponnampalam, E.N.; Cottrell, J.J.; Suleria, H.A.R.; Dunshea, F.R. Extraction and characterization of polyphenols from non-conventional edible plants and their antioxidant activities. Food Res. Int. 2022, 157, 111205. [Google Scholar] [CrossRef]
- Adhikari, P.; Kiess, A.; Adhikari, R.; Jha, R. An approach to alternative strategies to control avian coccidiosis and necrotic enteritis. J. Appl. Poult. Res. 2020, 29, 515–534. [Google Scholar] [CrossRef]
- Williams, R.B. Intercurrent coccidiosis and necrotic enteritis of chickens: Rational, integrated disease management by maintenance of gut integrity. Avian Pathol. 2010, 34, 159–180. [Google Scholar] [CrossRef]
- Cera, K.R.; Mahan, D.C.; Reinhart, G.A. Apparent fat digestibilities and performance responses of postweaning swine fed diets supplemented with coconut oil, corn oil or tallow. J. Anim. Sci. 1989, 67, 2040–2047. [Google Scholar] [CrossRef]
- Esmaeilnejad, B.; Tavassoli, M.; Asri-Rezaei, S.; Dalir-Naghadeh, B. Evaluation of antioxidant status and oxidative stress in sheep naturally infected with Babesia ovis. Vet. Parasitol. 2012, 185, 124–130. [Google Scholar] [CrossRef]
- Doreau, M.; Ferlay, A. Digestion and utilisation of fatty acids by ruminants. Anim. Feed Sci. Technol. 1994, 45, 379–396. [Google Scholar] [CrossRef]
- Chilliard, Y.; Ferlay, A. Dietary lipids and forages interactions on cow and goat milk fatty acid composition and sensory properties. Reprod. Nutr. Dev. 2004, 44, 467–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, T.C.; Bridges, W.C., Jr. Protection of fatty acids against ruminal biohydrogenation in cattle. Eur. J. Lipid Sci. Technol. 2007, 109, 778–789. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Sinclair, A.J.; Holman, B.W.B. The sources, synthesis and biological actions of omega-3 and omega-6 fatty acids in red meat: An overview. Foods 2021, 10, 1358. [Google Scholar] [CrossRef]
- Yamin, D.; Beena, V.; Ramnath, V.; Zarina, A.; Harikumar, S.; Venkatachalapathy, R.T.; Gleeja, V.L. Impact of thermal stress on physiological, behavioural and biochemical parameters in native and crossbred goats. Small Rumin. Res. 2022, 216, 106794. [Google Scholar] [CrossRef]
- Burnett, V.F.; Seymour, G.R.; Norng, S.; Jacobs, J.L.; Ponnampalam, E.N. Lamb growth performance and carcass weight from rotationally grazed perennial pasture systems compared with annual pasture systems with supplements. Anim. Prod. Sci. 2012, 52, 248–254. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Linden, N.P.; Mitchell, M.L.; Hopkins, D.L.; Jacobs, J.L. Production systems to deliver premium grade lambs to the growing international and Australian markets. Small Rumin. Res. 2017, 157, 32–39. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Dunshea, F.R.; Warner, R.D. Use of lucerne hay in ruminant feeds to improve animal productivity, meat nutritional value and meat preservation under a more variable climate. Meat Sci. 2020, 170, 108235. [Google Scholar] [CrossRef]
- Zárate, R.; el Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Functional roles of fatty acids and their effects on human health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnampalam, E.N.; Hopkins, D.L.; Bruce, H.; Li, D.; Baldi, G.; Bekhit, A.E.D.A. Causes and contributing factors to “dark cutting” meat: Current trends and future directions: A review. Compr. Rev. Food Sci. Food Saf. 2017, 16, 400–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payam, V.; Ponnampalam, E.N.; Kraft, J.; Mapiye, C.; Bermingham, E.N.; Watkins, P.J.; Proctor, S.D.; Dugan, M.E.R. Bioactivity and health effects of ruminant meat lipids. Invited Review. Meat Sci. 2020, 165, 108114. [Google Scholar]
- Branciari, R.; Galarini, R.; Giusepponi, D.; Trabalza-Marinucci, M.; Forte, C.; Roila, R.; Miraglia, D.; Servili, M.; Acuti, G.; Valiani, A. Oxidative status and presence of bioactive compounds in meat from chickens fed polyphenols extracted from olive oil industry waste. Sustainability 2017, 9, 1566. [Google Scholar] [CrossRef] [Green Version]
- Chedea, V.S.; Palade, L.M.; Pelmus, R.S.; Dragomir, C.; Taranu, I. Red grape pomace rich in polyphenols diet increases the antioxidant status in key organs-kidneys, liver, and spleen of piglets. Animals 2019, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Kafantaris, I.; Stagos, D.; Kotsampasi, B.; Hatzis, A.; Kypriotakis, A.; Gerasopoulos, K.; Makri, S.; Goutzourelas, N.; Mitsagga, C.; Giavasis, I. Grape pomace improves performance, antioxidant status, fecal microbiota and meat quality of piglets. Animal 2018, 12, 246–255. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.W.; Zhou, D.W.; Li, K. Effects of chestnut tannins on performance and antioxidative status of transition dairy cows. J. Dairy Sci. 2013, 96, 5901–5907. [Google Scholar] [CrossRef] [Green Version]
- Safari, M.; Ghasemi, E.; Alikhani, M.; Ansari-Mahyari, S. Supplementation effects of pomegranate by-products on oxidative status, metabolic profile, and performance in transition dairy cows. J. Dairy Sci. 2018, 101, 11297–11309. [Google Scholar] [CrossRef] [Green Version]
- Belščak-Cvitanović, A.; Durgo, K.; Huđek, A.; Bačun-Družina, V.; Komes, D. Overview of polyphenols and their properties. In Polyphenols: Properties, Recovery, and Applications; Balanakis, C.M., Ed.; Woodhead Publishing: Cambridge, UK, 2018; pp. 3–44. [Google Scholar]
- Bars-Cortina, D.; de Las Hazas, M.C.L.; Benavent-Vallés, A.; Motilva, M.J. Impact of dietary supplementation with olive and thyme phenols on alpha-tocopherol concentration in the muscle and liver of adult Wistar rats. Food Funct. 2018, 9, 1433–1443. [Google Scholar] [CrossRef]
- Tufarelli, V.; Laudadio, V.; Casalino, E. An extra-virgin olive oil rich in polyphenolic compounds has antioxidant effects in meat-type broiler chickens. Environ. Sci. Pollut. Res. 2016, 23, 6197–6204. [Google Scholar] [CrossRef]
- Van Vliet, S.; Provenza, F.D.; Kronberg, S.L. Health-promoting phytonutrients are higher in grass-fed meat and milk. Front. Sustain. Food Syst. 2021, 4, 555426. [Google Scholar] [CrossRef]
- Calkins, C.R.; Hodgen, J.M. A fresh look at meat flavor. Meat Sci. 2007, 77, 63–80. [Google Scholar] [CrossRef] [PubMed]
- Resconi, V.C.; Campo, M.M.; Furnols, M.F.i.; Montossi, F.; Sañudo, C. Sensory evaluation of castrated lambs finished on different proportions of pasture and concentrate feeding systems. Meat Sci. 2009, 83, 31–37. [Google Scholar] [CrossRef]
- Webb, E.C.; Erasmus, L.J. The effect of production system and management practices on the quality of meat products from ruminant livestock. S. Afr. J. Anim. Sci. 2013, 43, 413–423. [Google Scholar] [CrossRef] [Green Version]
- Ponnampalam, E.N.; Holman, B.W.B.; Kerry, J.P. Impact of animal nutrition on muscle composition and meat quality. In Meat Quality: Genetic and Environmental Factors; Przybylski, W., Hopkins, D.L., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 101–145. [Google Scholar]
- Holman, B.W.B.; Hopkins, D.L. The use of conventional laboratory-based methods to predict consumer acceptance of beef and sheep meat: A review. Meat Sci. 2021, 181, 108586. [Google Scholar] [CrossRef] [PubMed]
- Suleman, R.; Wang, Z.; Aadil, R.M.; Hui, T.; Hopkins, D.L.; Zhang, D. Effect of cooking on the nutritive quality, sensory properties and safety of lamb meat: Current challenges and future prospects. Meat Sci. 2020, 167, 108172. [Google Scholar] [CrossRef]
- Takahashi, M.; Tsuboyama-Kasaoka, N.; Nakatani, T.; Ishii, M.; Tsutsumi, S.; Aburatani, H.; Ezaki, O. Fish oil feeding alters liver gene expressions to defend against PPARα activation and ROS production. Am. J. Physiol.-Gastrointest. Liver Physiol. 2002, 282, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Ohata, T.; Fukuda, K.; Takahashi, M.; Sugimura, T.; Wakabayashi, K. Suppression of nitric oxide production in lipopolysaccharide-stimulated macrophage cells by omega 3 polyunsaturated fatty acids. Jpn. J. Cancer Res. 1997, 88, 234–237. [Google Scholar] [CrossRef]
- Karami, M.; Ponnampalam, E.N.; Hopkins, D.L. The effect of palm oil or canola oil on feedlot performance, plasma and tissue fatty acid profile and meat quality in goats. Meat Sci. 2013, 94, 165–169. [Google Scholar] [CrossRef]
- Cucchi, D.; Camacho-Munoz, D.; Certo, M.; Niven, J.; Smith, J.; Nicolaou, A.; Mauro, C. Omega-3 polyunsaturated fatty acids impinge on CD4+ T cell motility and adipose tissue distribution via direct and lipid mediator-dependent effects. Cardiovasc. Res. 2020, 116, 1006–1020. [Google Scholar] [CrossRef]
- Scaioli, E.; Liverani, E.; Belluzzi, A. The imbalance between n-6/n-3 polyunsaturated fatty acids and inflammatory bowel disease: A comprehensive review and future therapeutic perspectives. Int. J. Mol. Sci. 2017, 18, 2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poorghasemi, M.; Seidavi, A.; Qotbi, A.A.A.; Chambers, J.R.; Laudadio, V.; Tufarelli, V. Effect of dietary fat source on humoral immunity response of broiler chickens. Eur. Poult. Sci. 2015, 79, 92. [Google Scholar]
- Haddi, A.; Guendouz, M.; Tabet, S.A.; Mehedi, N.; Kheroua, O.; Saidi, D. Polyunsaturated fatty acids affect intestinal anaphylactic response in BALB/c mice sensitized with β-lactoglobulin. Rev. Française D’allergologie 2018, 58, 437–443. [Google Scholar] [CrossRef]
- Shen, L.; Yang, Y.; Ou, T.; Key, C.C.C.; Tong, S.H.; Sequeira, R.C.; Nelson, J.M.; Nie, Y.; Wang, Z.; Boudyguina, E. Dietary PUFAs attenuate NLRP3 inflammasome activation via enhancing macrophage autophagy. J. Lipid Res. 2017, 58, 1808–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogero, M.M.; Calder, P.C. Obesity, inflammation, toll-like receptor 4 and fatty acids. Nutrients 2018, 10, 432. [Google Scholar] [CrossRef] [Green Version]
- Körner, A.; Schlegel, M.; Theurer, J.; Frohnmeyer, H.; Adolph, M.; Heijink, M.; Giera, M.; Rosenberger, P.; Mirakaj, V. Resolution of inflammation and sepsis survival are improved by dietary Ω-3 fatty acids. Cell Death Differ. 2018, 25, 421–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nugroho, P.; Wiryawan, K.G.; Astuti, D.A.; Manalu, W. Stimulation of follicle growth and development during estrus in Ettawa Grade does fed a flushing supplement of different polyunsaturated fatty acids. Vet. World 2021, 14, 11. [Google Scholar] [CrossRef]
- Turchini, G.M.; Nichols, P.D.; Barrow, C.; Sinclair, A.J. Jumping on the omega-3 bandwagon: Distinguishing the role of long-chain and short-chain omega-3 fatty acids. Crit. Rev. Food Sci. Nutr. 2012, 52, 795–803. [Google Scholar] [CrossRef] [PubMed]
- McDowell, L.; Williams, S.N.; Hidiroglou, N.; Njeru, C.A.; Hill, G.M.; Ochoa, L.; Wilkinson, N.S. Vitamin E supplementation for the ruminant. Anim. Feed. Sci. Technol. 1996, 60, 273–296. [Google Scholar] [CrossRef]
- Holman, B.W.B.; Bailes, K.L.; Kerr, M.J.; Hopkins, D.L. Point of purchase fatty acid profile, oxidative status and quality of vacuum-packaged grass fed Australian beef held chilled for up to 12 weeks. Meat Sci. 2019, 158, 107878. [Google Scholar] [CrossRef]
- Resconi, V.C.; Escudero, A.; Campo, M.M. The development of aromas in ruminant meat. Molecules 2013, 18, 6748–6781. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, S. Reducing lipid peroxidation for improving colour stability of beef and lamb: On-farm considerations. J. Sci. Food Agric. 2012, 92, 719–729. [Google Scholar] [CrossRef]
- Williams, G.M.; Iatropoulos, M.J.; Whysner, J. Safety assessment of butylated hydroxyanisole and butylated hydroxytoluene as antioxidant food additives. Food Chem. Toxicol. 1999, 37, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Decker, E.A.; Goddard, J.M. Controlling lipid oxidation of food by active packaging technologies. Food Funct. 2013, 4, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Andrée, S.; Jira, W.; Schwind, K.H.; Wagner, H.; Schwägele, F. Chemical safety of meat and meat products. Meat Sci. 2010, 86, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, G.R.; Saldanha, T.; Soares, R.A.M.; Torres, E.A.F.S. Effect of natural antioxidant combinations on lipid oxidation in cooked chicken meat during refrigerated storage. Food Chem. 2012, 135, 1383–1390. [Google Scholar] [CrossRef]
- Estévez, M. Critical overview of the use of plant antioxidants in the meat industry: Opportunities, innovative applications and future perspectives. Meat Sci. 2021, 181, 108610. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, T.L.; Kerry, J.P.; Kerry, J.F.; Lynch, P.B.; Buckley, D.J. Evaluation of the antioxidant potential of natural food/plant extracts as compared with synthetic antioxidants and vitamin E in raw and cooked pork patties. Meat Sci. 2001, 58, 45–52. [Google Scholar] [CrossRef]
- Haak, L.; Raes, K.; De Smet, S. Effect of plant phenolics, tocopherol and ascorbic acid on oxidative stability of pork patties. J. Sci. Food Agric. 2009, 89, 1360–1365. [Google Scholar] [CrossRef]
- Estévez, M. Oxidative damage to poultry: From farm to fork. Poult. Sci. 2015, 94, 1368–1378. [Google Scholar] [CrossRef]
- Nathan, C.; Cunningham-Bussel, A. Beyond oxidative stress: An immunologist’s guide to reactive oxygen species. Nat. Rev. Immunol. 2013, 13, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugrin, J.; Rosenblatt-Velin, N.; Parapanov, R.; Liaudet, L. The role of oxidative stress during inflammatory processes. Biol. Chem. 2014, 395, 203–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lykkesfeldt, J.; Svendsen, O. Oxidants and antioxidants in disease: Oxidative stress in farm animals. Vet. J. 2007, 173, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.; Kotur-Stevuljevic, J.; Berneis, K.; Spinas, G.; Rini, G.B.; Jelic-Ivanovic, Z.; Spasojevic-Kalimanovska, V.; Vekic, J. Atherogenic dyslipidemia and oxidative stress: A new look. Transl. Res. 2009, 153, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Gür, S.; Türk, G.; Demirci, E.; Yüce, A.; Sönmez, M.; Özer, Ş.; Aksu, E.H. Effect of pregnancy and foetal number on diameter of corpus luteum, maternal progesterone concentration and oxidant/antioxidant balance in ewes. Reprod. Domest. Anim. 2011, 46, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Toescu, V.; Nuttall, S.L.; Martin, U.; Nightingale, P.; Kendall, M.J.; Brydon, P.; Dunne, F. Changes in plasma lipids and markers of oxidative stress in normal pregnancy and pregnancies complicated by diabetes. Clin. Sci. 2004, 106, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Shoji, H.; Koletzko, B. Oxidative stress and antioxidant protection in the perinatal period. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 324–328. [Google Scholar] [CrossRef]
- Golightly, E.; Jabbour, H.N.; Norman, J.E. Endocrine immune interactions in human parturition. Mol. Cell. Endocrinol. 2011, 335, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Van Engelen, E.; De Groot, M.W.; Breeveld-Dwarkasing, V.N.A.; Everts, M.E.; Van Der Weyden, G.C.; Taverne, M.A.M.; Rutten, V.P.M.G. Cervical ripening and parturition in cows are driven by a cascade of pro-inflammatory cytokines. Reprod. Domest. Anim. 2009, 44, 834–841. [Google Scholar] [CrossRef]
- Lacetera, N. Impact of climate change on animal health and welfare. Anim. Front. 2019, 9, 26–31. [Google Scholar] [CrossRef]
- Ronchi, B.; Lacetera, N.; Bernabucci, U.; Nardone, A.; Verini Supplizi, A. Distinct and common effects of heat stress and restricted feeding on metabolic status of Holstein heifers. Zootec. E Nutr. Anim. 1999, 25, 71–80. [Google Scholar]
- Wheelock, J.B.; Rhoads, R.P.; VanBaale, M.J.; Sanders, S.R.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batistel, F.; Arroyo, J.M.; Bellingeri, A.; Wang, L.; Saremi, B.; Parys, C.; Trevisi, E.; Cardoso, F.C.; Loor, J.J. Ethyl-cellulose rumen-protected methionine enhances performance during the periparturient period and early lactation in Holstein dairy cows. J. Dairy Sci. 2017, 100, 7455–7467. [Google Scholar] [CrossRef] [Green Version]
- Osorio, J.S.; Trevisi, E.; Ji, P.; Drackley, J.K.; Luchini, D.; Bertoni, G.; Loor, J.J. Biomarkers of inflammation, metabolism, and oxidative stress in blood, liver, and milk reveal a better immunometabolic status in peripartal cows supplemented with Smartamine M or MetaSmart. J. Dairy Sci. 2014, 97, 7437–7450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adam-Vizi, V.; Chinopoulos, C. Bioenergetics and the formation of mitochondrial reactive oxygen species. Trends Pharmacol. Sci. 2006, 27, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Cowley, F.C.; Barber, D.G.; Houlihan, A.V.; Poppi, D.P. Immediate and residual effects of heat stress and restricted intake on milk protein and casein composition and energy metabolism. J. Dairy Sci. 2015, 98, 2356–2368. [Google Scholar] [CrossRef] [Green Version]
- Nardone, A.; Lacetera, N.; Bernabucci, U.; Ronchi, B. Composition of colostrum from dairy heifers exposed to high air temperatures during late pregnancy and the early postpartum period. J. Dairy Sci. 1997, 80, 838–844. [Google Scholar] [CrossRef]
- Ríus, A.G. Invited Review: Adaptations of protein and amino acid metabolism to heat stress in dairy cows and other livestock species. Appl. Anim. Sci. 2019, 35, 39–48. [Google Scholar] [CrossRef]
- Nan, X.; Bu, D.; Li, X.; Wang, J.; Wei, H.; Hu, H.; Zhou, L.; Loor, J.J. Ratio of lysine to methionine alters expression of genes involved in milk protein transcription and translation and mTOR phosphorylation in bovine mammary cells. Physiol. Genom. 2014, 46, 268–275. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Ledesma, B.; Amigo, L. La leche como fuente de antioxidantes naturales. Aliment. Nutr. Salud 2004, 11, 61–65. [Google Scholar]
- Heard, J.W.; Stockdale, C.R.; Walker, G.P.; Leddin, C.M.; Dunshea, F.R.; McIntosh, G.H.; Shields, P.M.; McKenna, A.; Young, G.P.; Doyle, P.T. Increasing selenium concentration in milk: Effects of amount of selenium from yeast and cereal grain supplements. J. Dairy Sci. 2007, 90, 4117–4127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, G.P.; Dunshea, F.R.; Heard, J.W.; Stockdale, C.R.; Doyle, P.T. Output of selenium in milk, urine, and feces in proportional to selenium intake in dairy cows fed a total mixed ration supplemented with selenium yeast. J. Dairy Sci. 2010, 93, 4644–4650. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lindmark-Månsson, H.; Gorton, L.; Åkesson, B. Antioxidant capacity of bovine milk as assayed by spectrophotometric and amperometric methods. Int. Dairy J. 2003, 13, 927–935. [Google Scholar] [CrossRef]
- Clausen, M.R.; Connolly, C.; Skibsted, L.H.; Stagsted, J. Oxidative stability of bovine milk determined by individual variability in herd irrespective of selenium status. Int. Dairy J. 2010, 20, 507–513. [Google Scholar] [CrossRef]
- Weiss, W.P. Antioxidant nutrients and milk quality. In Proceedings of the Roche Symposium, Pacific Northwest Nutrition Conference, Vancouver, BC, Canada, 13–15 January 2010; pp. 1–10. [Google Scholar]
- Khan, I.T.; Nadeem, M.; Imran, M.; Ullah, R.; Ajmal, M.; Jaspal, M.H. Antioxidant properties of Milk and dairy products: A comprehensive review of the current knowledge. Lipids Health Dis. 2019, 18, 41. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.E.; Kim, J.W.; Cheon, S.; Nam, M.S.; Kim, K.K. Alpha-Casein and beta-lactoglobulin from cow milk exhibit antioxidant activity: A plausible link to antiaging effects. J. Food Sci. 2019, 84, 3083–3090. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, S.; Prakash, V. Bioactive peptides from bovine milk α-casein: Isolation, characterization and multifunctional properties. Int. J. Pept. Res. Ther. 2010, 16, 7–15. [Google Scholar] [CrossRef]
- Miao, J.; Liao, W.; Pan, Z.; Wang, Q.; Duan, S.; Xiao, S.; Yang, Z.; Cao, Y. Isolation and identification of iron-chelating peptides from casein hydrolysates. Food Funct. 2019, 10, 2372–2381. [Google Scholar] [CrossRef]
- Smialowska, A.; Matia-Merino, L.; Carr, A.J. Oxidative stability of iron fortified goat and cow milk and their peptide isolates. Food Chem. 2017, 237, 1021–1024. [Google Scholar] [CrossRef]
- Boor, K.J. Fluid dairy product quality and safety: Looking to the future. J. Dairy Sci. 2001, 84, 1–11. [Google Scholar] [CrossRef]
- Guha, S.; Sharma, H.; Deshwal, G.K.; Rao, P.S. A comprehensive review on bioactive peptides derived from milk and milk products of minor dairy species. Food Prod. Process. Nutr. 2021, 3, 2. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Corresponding Substrates | Reactive Oxygen Species |
|---|---|---|
| Acyl-CoA oxidases | Long chain fatty acids, methyl branched chain fatty acids, bile acid intermediates | H2O2 |
| D-aspartate oxidase | D-aspartate, N-methyl-D-aspartate | H2O2 |
| Xanthine oxidase | Xanthine | H2O2, O2•− |
| D-amino acid oxidase | D-proline | H2O2 |
| Polyamine oxidase | N-Acetyl spermine, spermidine | H2O2 |
| L-pipecolic acid oxidase | L-pipecolic acid | H2O2 |
| Urate oxidase | Uric acid | H2O2 |
| L-α-hydroxyacid oxidase | Glycolate, lactate | H2O2 |
| Cattle Production | Production Systems | Sheep Production |
|---|---|---|
 | Grazing systems offering high to moderate antioxidant function (cattle and sheep grazing tall fescue pasture) (left) and perennial ryegrass (right) in spring. |  |
 | Grazing systems offering marginal antioxidants and point of ROS development and free radical formation. Photos taken in autumn season. |  |
 | Poor antioxidant function and oxidative stress due to malnutrition caused by drought and lack of green grass availability. Photos taken in summer season. |  |
| Incidence of high ketone bodies in the blood, increased somatic cells in milk, and dark cutting meat in cattle. | Initiation of cellular dysfunction and metabolic disorders, leading to reduced performance and product quality. | Lower muscle: fat ratio in the carcass and reduced eating quality of meat in sheep. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponnampalam, E.N.; Kiani, A.; Santhiravel, S.; Holman, B.W.B.; Lauridsen, C.; Dunshea, F.R. The Importance of Dietary Antioxidants on Oxidative Stress, Meat and Milk Production, and Their Preservative Aspects in Farm Animals: Antioxidant Action, Animal Health, and Product Quality—Invited Review. Animals 2022, 12, 3279. https://doi.org/10.3390/ani12233279
Ponnampalam EN, Kiani A, Santhiravel S, Holman BWB, Lauridsen C, Dunshea FR. The Importance of Dietary Antioxidants on Oxidative Stress, Meat and Milk Production, and Their Preservative Aspects in Farm Animals: Antioxidant Action, Animal Health, and Product Quality—Invited Review. Animals. 2022; 12(23):3279. https://doi.org/10.3390/ani12233279
Chicago/Turabian StylePonnampalam, Eric N., Ali Kiani, Sarusha Santhiravel, Benjamin W. B. Holman, Charlotte Lauridsen, and Frank R. Dunshea. 2022. "The Importance of Dietary Antioxidants on Oxidative Stress, Meat and Milk Production, and Their Preservative Aspects in Farm Animals: Antioxidant Action, Animal Health, and Product Quality—Invited Review" Animals 12, no. 23: 3279. https://doi.org/10.3390/ani12233279
APA StylePonnampalam, E. N., Kiani, A., Santhiravel, S., Holman, B. W. B., Lauridsen, C., & Dunshea, F. R. (2022). The Importance of Dietary Antioxidants on Oxidative Stress, Meat and Milk Production, and Their Preservative Aspects in Farm Animals: Antioxidant Action, Animal Health, and Product Quality—Invited Review. Animals, 12(23), 3279. https://doi.org/10.3390/ani12233279