

Microsatellite Analysis Revealed Potential DNA Markers for Gestation Length and Sub-Population Diversity in Kari Sheep
 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Selection and Experimental Design
2.2. Isolation of Genomic DNA from Blood
2.3. Polymerase Chain Reaction
2.4. Data Collection
2.5. Statistical Analysis
3. Results
3.1. Gestation Length Variation among Kari Subtypes
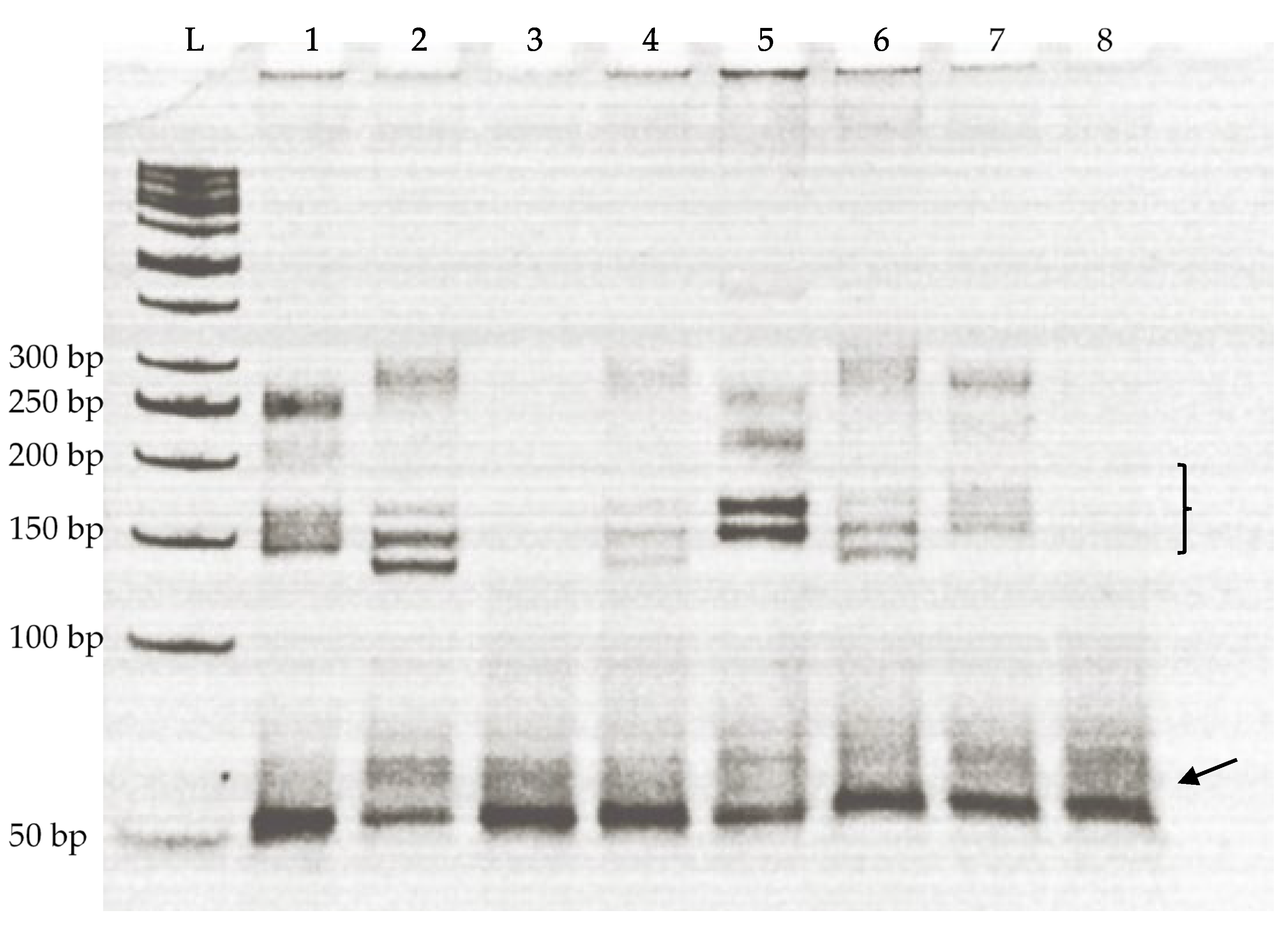
3.2. Association of Microsatellite Polymorphism and Gestation Length
3.3. Genetic Diversity among the Subtypes
3.3.1. Intra-Subtype Genetic Variation
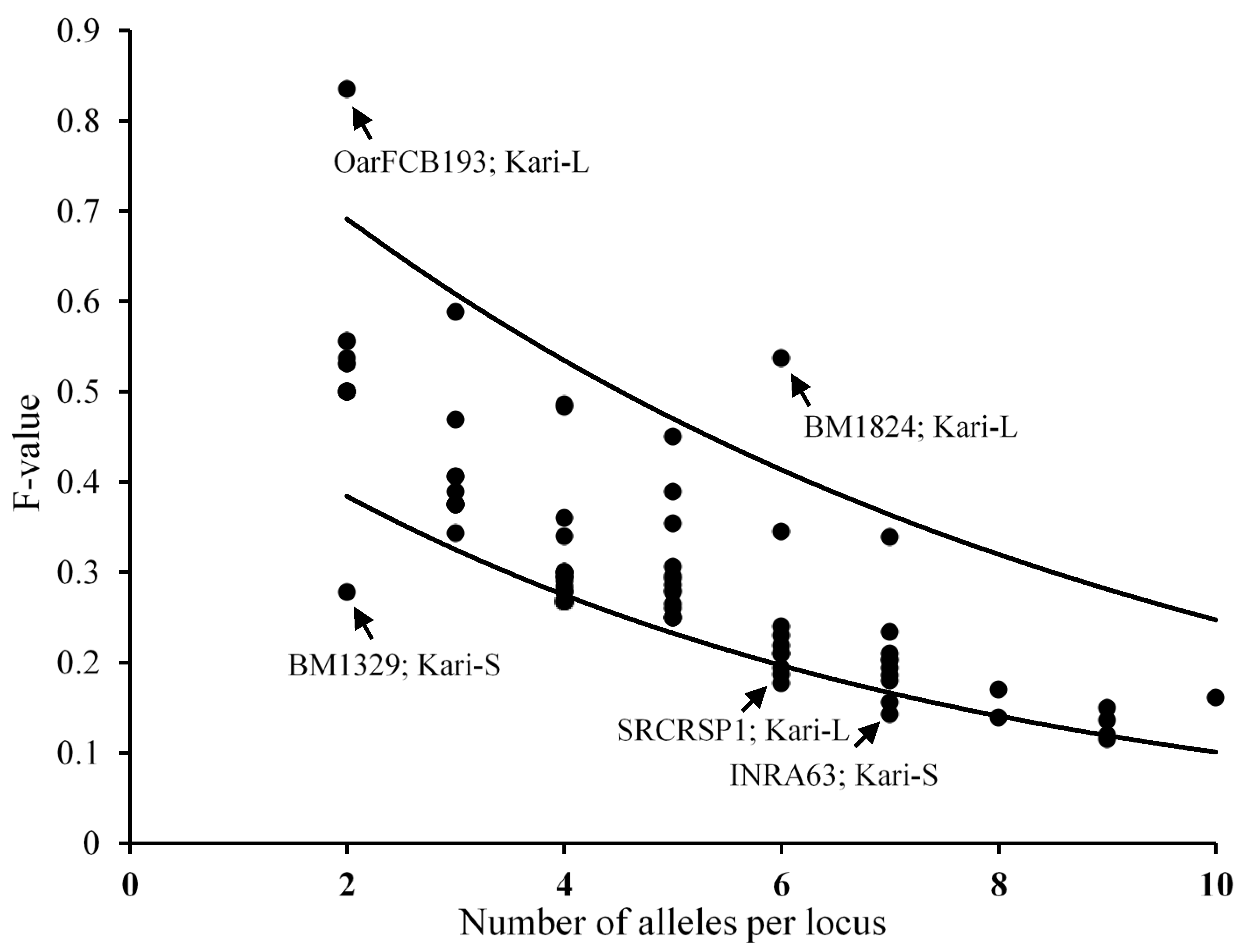
3.3.2. Allelic Polymorphisms
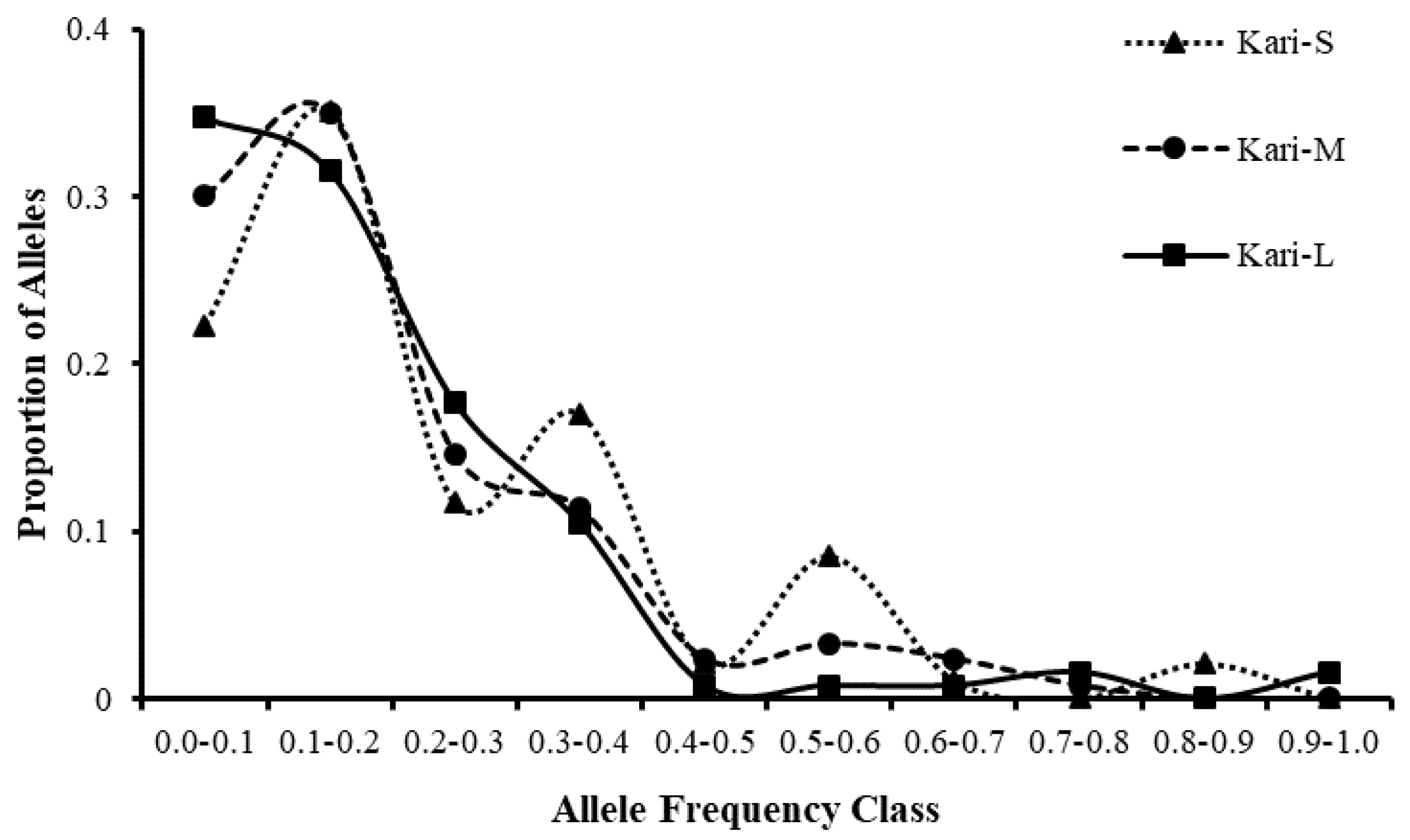
3.3.3. Ewens-Watterson Test for Microsatellites’ Neutrality
3.4. Genetic Assignment of Individuals to Their Respective Subtype
3.5. Population Sub-Structure
3.6. Mutation Drift Equilibrium in Kari Subtypes
4. Discussion
4.1. Genetics of Gestation Length Variation in Sheep
4.2. Genetic Diversity among Subtypes
4.3. Assigning Individuals to Subtypes
4.4. Inter-Subtype Breeding and Differentiation
4.5. Population Bottleneck and Mutation Drift
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Ahmad, S. Performance and Phylogenetic Position of Kari Sheep in Pakistan; University of Agriculture Faisalabad: Faisalabad, Pakistan, 2008. [Google Scholar]
- IUCN. NWFP Chitral—An Integrated Development Vision (Chitral Coservation Strategy); IUCN The World Conservation Union Pakistan and NWFP: Karachi, Pakistan, 2004; p. 103. [Google Scholar]
- Ahmad, S.; Riaz, M.; Siddiqui, M.M.; Habib, G. The Kari sheep—A genetic heritage. J. Anim. Plant. Sci. 2002, 12, 14–16. [Google Scholar]
- AKRSP. The Wheel of Fortune, a Study of the Patti Production System in Chitral; Agha Khan Rural Support Program, Agha Khan Foundation: London, UK, 1996; p. 70. [Google Scholar]
- Wallace, J.M.; Milne, J.S.; Aitken, R.P. Early pregnancy weight gain and fat accrual predict pregnancy outcome in growing adolescent sheep. Reproduction 2021, 161, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Gatford, K.L.; Kennaway, D.J.; Liu, H.; Kleemann, D.O.; Kuchel, T.R.; Varcoe, T.J. Simulated shift work disrupts maternal circadian rhythms and metabolism, and increases gestation length in sheep. J. Physiol. 2019, 597, 1889–1904. [Google Scholar] [CrossRef] [Green Version]
- Cleal, J.K.; Poore, K.R.; Newman, J.P.; E Noakes, D.; A Hanson, M.; Green, L.R. The Effect of Maternal Undernutrition in Early Gestation on Gestation Length and Fetal and Postnatal Growth in Sheep. Pediatr. Res. 2007, 62, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, S.; Froy, H.; Underwood, S.L.; Dorrens, J.; Seeker, L.A.; Watt, K.; Wilbourn, R.V.; Pilkington, J.G.; Harrington, L.; Pemberton, J.M.; et al. The association between female reproductive performance and leukocyte telomere length in wild Soay sheep. Mol. Ecol. 2021, 31, 6184–6196. [Google Scholar] [CrossRef] [PubMed]
- Künzle, H.; Nautrup, C.P.; Schwarzenberger, F. High inter-individual variation in the gestation length of the hedgehog tenrec, Echinops telfairi (Afrotheria). Anim. Reprod. Sci. 2007, 97, 364–374. [Google Scholar] [CrossRef]
- Rowell, J.E.; Shipka, M.P. Variation in gestation length among captive reindeer (Rangifer tarandus tarandus). Theriogenology 2009, 72, 190–197. [Google Scholar] [CrossRef]
- Dutta, N.; Singh, R.K.; Pathak, A.; Mohindra, V.; Mandal, S.; Kaur, G.; Lal, K.K. Mitochondrial DNA markers reveal genetic connectivity among populations of Osteoglossiform fish Chitala chitala. Mol. Biol. Rep. 2020, 47, 8579–8592. [Google Scholar] [CrossRef]
- Chan, A.H.E.; Chaisiri, K.; Saralamba, S.; Morand, S.; Thaenkham, U. Assessing the suitability of mitochondrial and nuclear DNA genetic markers for molecular systematics and species identification of helminths. Parasites Vectors 2021, 14, 233. [Google Scholar] [CrossRef]
- Kobayashi, M.; Yusa, Y.; Sekino, M. Microsatellite DNA markers applicable to paternity inference in the androdioecious gooseneck barnacle Octolasmis warwickii (Lepadiformes: Poecilasmatidae). Mol. Biol. Rep. 2020, 47, 4885–4890. [Google Scholar] [CrossRef]
- Sasazaki, S. Development of DNA markers for improvement of meat quality in a Japanese Black cattle population in Hyogo Prefecture. Anim. Sci. J. Nihon Chikusan Gakkaiho 2021, 92, e13663. [Google Scholar] [CrossRef]
- Chen, C.-C.; Huang, C.-W.; Lin, C.-Y.; Ho, C.-H.; Pham, H.N.; Hsu, T.-H.; Lin, T.-T.; Chen, R.-H.; Yang, S.-D.; Chang, C.-I.; et al. Development of Disease-Resistance-Associated Microsatellite DNA Markers for Selective Breeding of Tilapia (Oreochromis spp.) Farmed in Taiwan. Genes 2021, 13, 99. [Google Scholar] [CrossRef] [PubMed]
- Geldermann, H.; Mir, M.R.; Kuss, A.W.; Bartenschlager, H. OLA-DRB1 microsatellite variants are associated with ovine growth and reproduction traits. Genet. Sel. Evol. 2006, 38, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S. Gestation length of kari sheep. Nat. Précéd. 2008, 1620, 1. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T. Popgene 1.31; Molecular Biology and Technology Center, University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Raymond, M.; Rousset, F. GENEPOP (version 4.3): Population genetic software for exact test and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Piry, S.; Alapetite, A.; Cornuet, J.-M.; Paetkau, D.; Baudouin, L.; Estoup, A. Geneclass2: A Software for Genetic Assignment and First-Generation Migrant Detection. J. Hered. 2004, 95, 536–539. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT, 2.9.3.2; Department of Ecology & Evolution, Biology Building: Lausanne, Switzerland, 2001. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.-M. Computer note. Bottleneck: A computer program for detecting recent reductions in the effective size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Anderson, G.; Bradford, G.; Cupps, P. Length of gestation in ewes carrying lambs of two different breeds. Theriogenology 1981, 16, 119–129. [Google Scholar] [CrossRef]
- Li, L.; Brown, D. Estimation of genetic parameters for lambing ease, birthweight and gestation length in Australian sheep. Anim. Prod. Sci. 2016, 56, 934–940. [Google Scholar] [CrossRef]
- Iyiola-Tunji, A.; Akpa, G.; Nwagu, B.; Adeyinka, I.; Osuhor, C.; Lawal, T.; Ojo, O. Relationship between gestation length and birth weight in Nigerian Sheep and their crosses. Anim. Prod. 2010, 12, 135–138. [Google Scholar]
- Cloete, S.; Snyman, M.; Herselman, M. Productive performance of Dorper sheep. Small Rumin. Res. 2000, 36, 119–135. [Google Scholar] [CrossRef]
- Bessette, N.W.; Rurak, D.W. Chronic fetal and maternal instrumentation in pregnant sheep: Effect on gestation length and birthweight. Reprod. Fertil. Dev. 2010, 22, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.L.C.; Santini, L.; Diniz, A.L.; Munhoz, C.D.F. Microsatellite markers: What they mean and why they are so useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef] [Green Version]
- Lei, Y.; Zhou, Y.; Price, M.; Song, Z. Genome-wide characterization of microsatellite DNA in fishes: Survey and analysis of their abundance and frequency in genome-specific regions. BMC Genom. 2021, 22, 421. [Google Scholar] [CrossRef]
- Vinces, M.D.; Legendre, M.; Caldara, M.; Hagihara, M.; Verstrepen, K.J. Unstable tandem repeats in promoters confer transcriptional evolvability. Science 2009, 324, 1213–1216. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, F.C.; Crawford, A.M. Ovine dinucleotide repeat polymorphism at the MAF214 locus. Anim. Genet. 1992, 23, 394. [Google Scholar] [CrossRef]
- Castillo-Rodríguez, R.G.; Segura-León, O.L.; Hernández-Rodríguez, M.; Serna-Lagunes, R.; Salinas-Ruiz, J.; Salazar-Ortiz, J. Genetic Diversity of Creole Sheep Managed by Indigenous Communities of the Central Region of Veracruz, Mexico. Animals 2022, 12, 456. [Google Scholar] [CrossRef]
- FAO. Genetic characterization of livestock populations and its use in conservation decision making. In The Role of Biotechnology in Exploring and Protecting Agricultural Genetic Resources; Ruane, J., Sonnino, A., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2005. [Google Scholar]
- Musavi, S.A.A.; Ahmad, S.; Ibrahim, M. Molecular characterization of Hazaragie sheep native to Central Afghanistan. Ind. J. Anim. Sci. 2011, 81, 711–717. [Google Scholar]
- Ibrahim, M.; Ahmad, S.; Durrani, I.S.; Iqbal, A.; Munir, I.; Swati, Z.A. Genetic Polymorphism and Bottleneck Analysis of Balkhi, Hashtnagri, and Michni Sheep Populations Using Microsatellite Markers. Anim. Biotechnol. 2017, 29, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Al-Atiyat, R.M.; Aljumaah, R.S.; Alshaikh, M.A.; Abudabos, A.M. Microsatellite-Based Genetic Structure and Diversity of Local Arabian Sheep Breeds. Front. Genet. 2018, 9, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Sharma, R.; Pandey, A.K.; Gour, D.S.; Malik, G.; Ahlawat, S.P.S.; Jain, A. Genetic diversity and bottleneck analysis of Indian bellary sheep by microsatellite markers. Russ. J. Genet. 2007, 43, 996–1005. [Google Scholar] [CrossRef]
- Ibrahim, M.; Ahmad, S.; Swati, Z.A.; Khan, M.S. Genetic diversity in Balkhi, Hashtnagri and Michni sheep populations using SSR markers. Afr. J. Biotech. 2010, 9, 7617–7628. [Google Scholar]
- Elliott, J.M.; Manly, B.J.F. The Statistics of Natural Selection on Animal Populations; Routledge, Chapman & Hall, Incorporated: London, UK, 1985; Volume 56, p. 373. [Google Scholar] [CrossRef]
- Nielsen, R. Statistical tests of selective neutrality in the age of genomics. Heredity 2001, 86, 641–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornuet, J.M.; Piry, S.; Luikart, G.; Estoup, A.; Solignac, M. New methods employing multilocous genotypes to select or exclude populations as origins of individuals. Genetics 1999, 153, 1989–2000. [Google Scholar] [CrossRef]
- Goldstein, D.B.; Ruiz Linares, A.; Cavalli-Sforza, L.L.; Feldman, M.W. An evaluation of genetic distances for use with microsatellite loci. Genetics 1995, 139, 463–471. [Google Scholar] [CrossRef]
- Hartl, D.L.; Clark, A.G. Principles of Population Genetics, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 1997. [Google Scholar]
- Shriver, M.D.; Jin, L.; Chakraborty, R.; Boerwinkle, E. VNTR allele frequency distributions under the step-wise mutation model: A computer simulation approach. Genetics 1993, 134, 983–993. [Google Scholar] [CrossRef]
- Alves, R.R.N.; Policarpo, I.D.S. Animals and Human Health: Where Do They Meet? Ethnozoology 2018, 233, 59. [Google Scholar]
- Mi, E.; Mi, E.; Jeggo, M. Where to Now for One Health and Ecohealth? Ecohealth 2016, 13, 12–17. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subtype | Sample Size | Gestation Length | Litter Size | Birth Weight |
|---|---|---|---|---|
| Kari-S | 24 | 100.7 ± 1.8 a | 1.54 ± 0.12 | 1.92 ± 0.13 |
| Kari-M | 26 | 123.1 ± 1.0 b | 1.57 ± 0.12 | 2.23 ± 0.12 |
| Kari-L | 28 | 143.8 ± 1.5 c | 1.53 ± 0.10 | 2.26 ± 0.11 |
| Sig. | *** | ns | ns |
| Subtype | Np | NT | Na | Ne | AR | Ho | He | FIS | PIC | HWE (p Value) |
|---|---|---|---|---|---|---|---|---|---|---|
| Kari-S | 21 | 92 | 4.318 | 3.451 | 2.790 | 0.812 | 0.659 | −0.136 | 0.578 | 0.275 |
| Kari-M | 22 | 121 | 5.454 | 3.798 | 2.822 | 0.761 | 0.722 | 0.006 | 0.628 | 0.098 |
| Kari-L | 21 | 119 | 5.454 | 4.171 | 2.889 | 0.757 | 0.691 | 0.040 | 0.622 | 0.152 |
| Subtype | Frequency Criteria | Bayesian Criteria | Distance Criteria | |||||
|---|---|---|---|---|---|---|---|---|
| R&M | B&L | DS | DM | DA | DC | DAS | ||
| Kari-S | 66.67 | 66.67 | 50.00 | 83.33 | 83.33 | 66.67 | 66.67 | 83.33 |
| Kari-M | 63.64 | 54.55 | 72.73 | 72.73 | 72.73 | 72.73 | 72.73 | 54.55 |
| Kari-L | 83.33 | 83.33 | 75.00 | 75.00 | 66.67 | 83.33 | 83.33 | 58.33 |
| Kari Subtypes | Kari Subtypes | ||
|---|---|---|---|
| Kari-M | Kari-L | ||
| Population differentiation (FST) | Kari-S | 0.147 ± 0.021 * | 0.149 ± 0.022 ** |
| Kari-M | 0.105 ± 0.022 * | ||
| Total inbreeding (FIT) | Kari-S | 0.030 ± 0.047 * | 0.015 ± 0.042 |
| Kari-M | 0.046 ± 0.041 ** | ||
| Gene flow (NM) | Kari-S | 3.779 ± 0.753 | 4.234 ± 0.721 |
| Kari-M | 9.315 ± 2.609 | ||
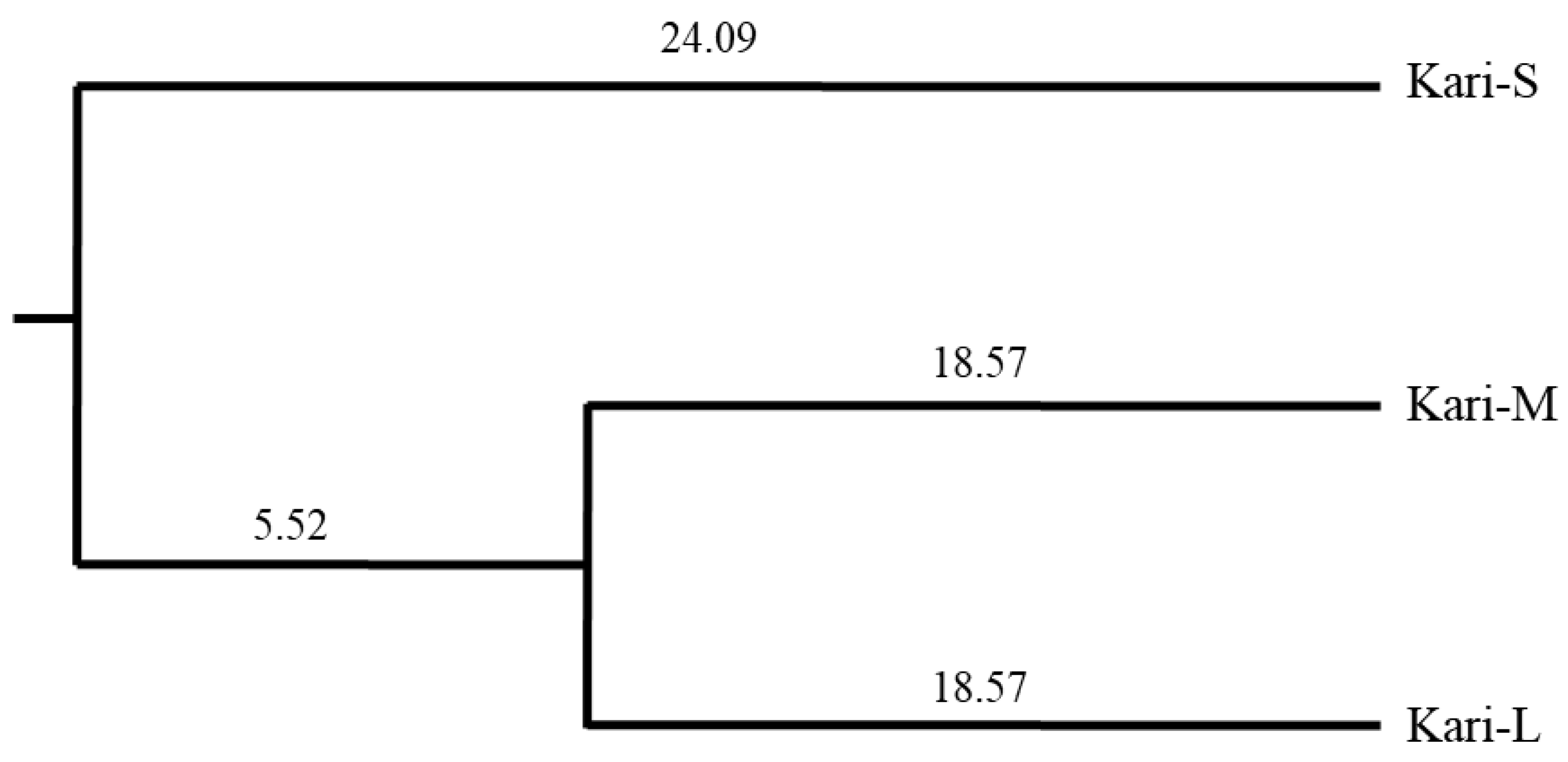
| Genetic distance | Kari-S | 0.285 | 0.280 |
| Kari-M | 0.205 | ||
| Subtype | Mutation Model | Sign Test | Standardized Differences Test | WS Rank Test | |||
|---|---|---|---|---|---|---|---|
| Hee | He | p | T2 | p | p | ||
| Kari-S | IAM | 12.46 | 19 | 0.003 | 5.07 | 0.000 | 0.000 |
| SMM | 12.74 | 17 | 0.048 | 3.06 | 0.001 | 0.008 | |
| TPM | 12.58 | 19 | 0.003 | 4.16 | 0.000 | 0.000 | |
| Kari-M | IAM | 13.47 | 19 | 0.013 | 3.91 | 0.000 | 0.000 |
| SMM | 13.59 | 16 | 0.210 * | 0.82 | 0.205 * | 0.172 * | |
| TPM | 13.48 | 17 | 0.098 * | 2.63 | 0.004 | 0.009 | |
| Kari-L | IAM | 13.11 | 20 | 0.002 | 4.19 | 0.000 | 0.000 |
| SMM | 13.24 | 18 | 0.032 | 1.70 | 0.044 | 0.007 | |
| TPM | 13.33 | 18 | 0.035 | 2.99 | 0.001 | 0.000 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, M.; Ahmad, S.; Ud Din, I.; Ahmad, W.; Ahmad, I.; Khan, S.H.; Ul Haq, I.; Zeb, J.; Sparagano, O.A. Microsatellite Analysis Revealed Potential DNA Markers for Gestation Length and Sub-Population Diversity in Kari Sheep. Animals 2022, 12, 3292. https://doi.org/10.3390/ani12233292
Ibrahim M, Ahmad S, Ud Din I, Ahmad W, Ahmad I, Khan SH, Ul Haq I, Zeb J, Sparagano OA. Microsatellite Analysis Revealed Potential DNA Markers for Gestation Length and Sub-Population Diversity in Kari Sheep. Animals. 2022; 12(23):3292. https://doi.org/10.3390/ani12233292
Chicago/Turabian StyleIbrahim, Muhammad, Sohail Ahmad, Israr Ud Din, Waqas Ahmad, Ijaz Ahmad, Sher Hayat Khan, Ihtesham Ul Haq, Jehan Zeb, and Olivier Andre Sparagano. 2022. "Microsatellite Analysis Revealed Potential DNA Markers for Gestation Length and Sub-Population Diversity in Kari Sheep" Animals 12, no. 23: 3292. https://doi.org/10.3390/ani12233292
APA StyleIbrahim, M., Ahmad, S., Ud Din, I., Ahmad, W., Ahmad, I., Khan, S. H., Ul Haq, I., Zeb, J., & Sparagano, O. A. (2022). Microsatellite Analysis Revealed Potential DNA Markers for Gestation Length and Sub-Population Diversity in Kari Sheep. Animals, 12(23), 3292. https://doi.org/10.3390/ani12233292