
Sustainable Valorization of Tomato Pomace (Lycopersicon esculentum) in Animal Nutrition: A Review

 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. The Production and Nutritional Value of TP
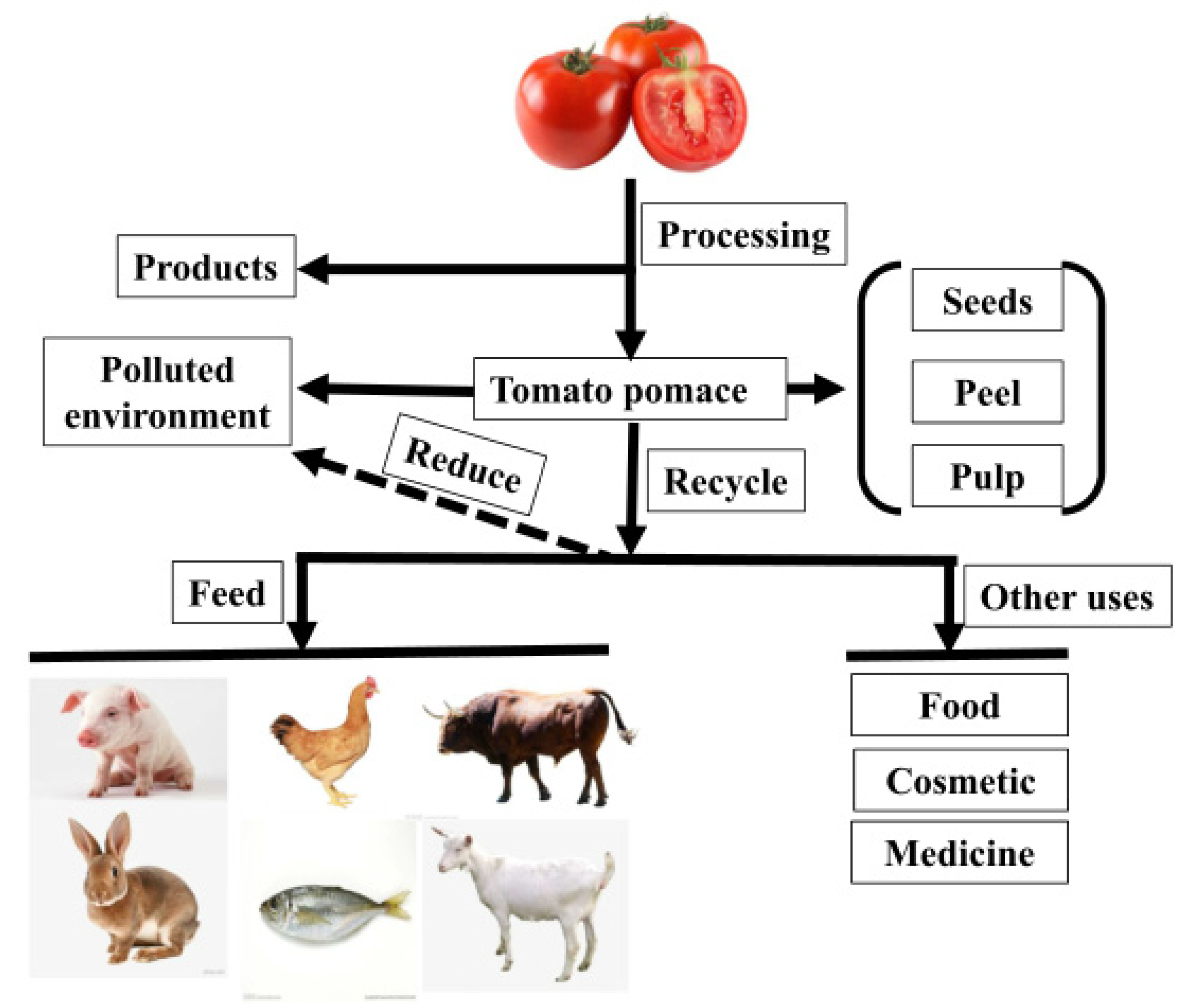
2.1. Production of TP
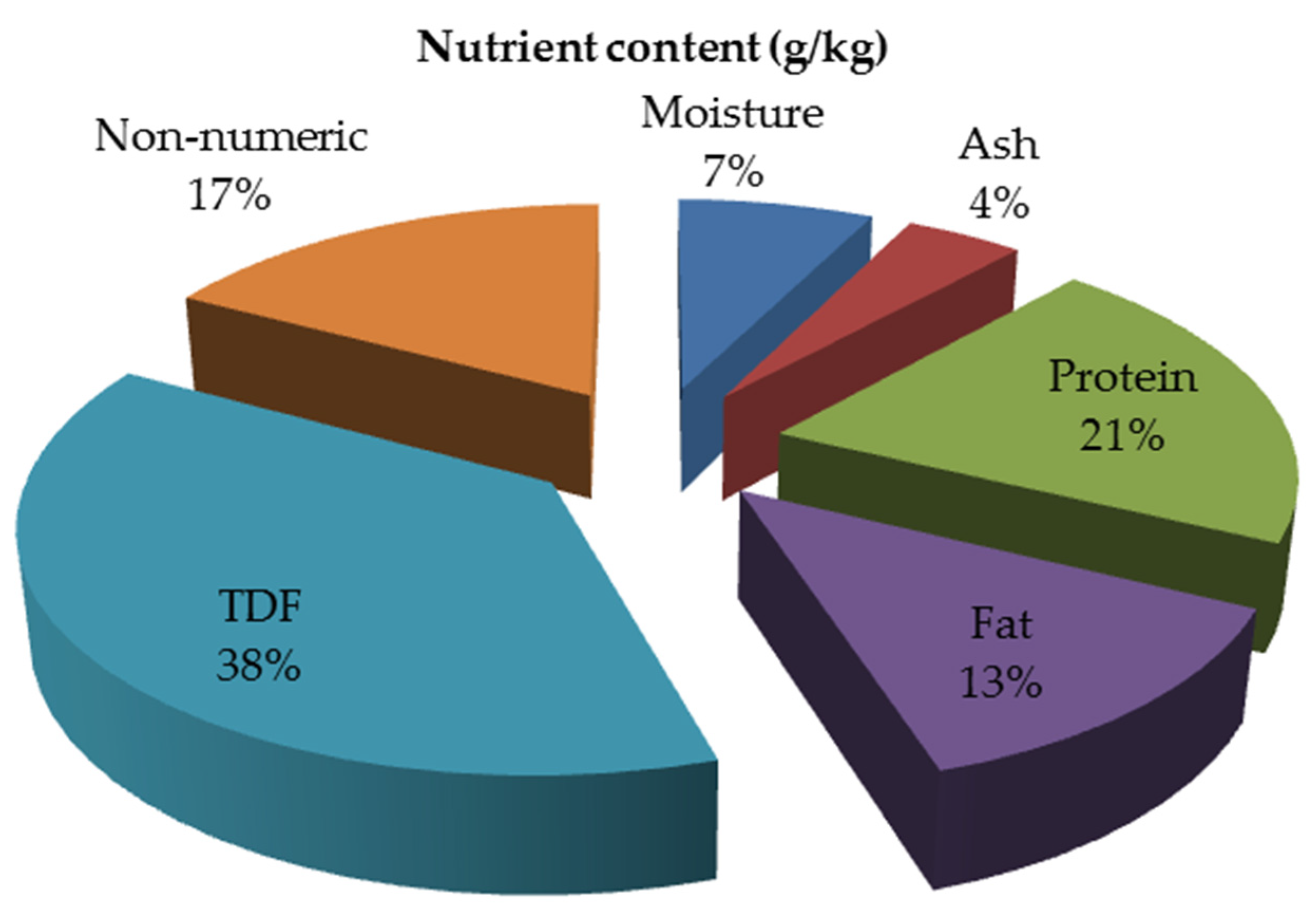
2.2. Regular Nutritional Content of TP
2.3. Antioxidant Potency of TP
2.4. Mineral Composition
2.5. Fatty Acid Profile
2.6. Amino Acid Profile
3. Antioxidant Mechanisms of Bioactive Substances
3.1. Total Phenol Content (TPC)
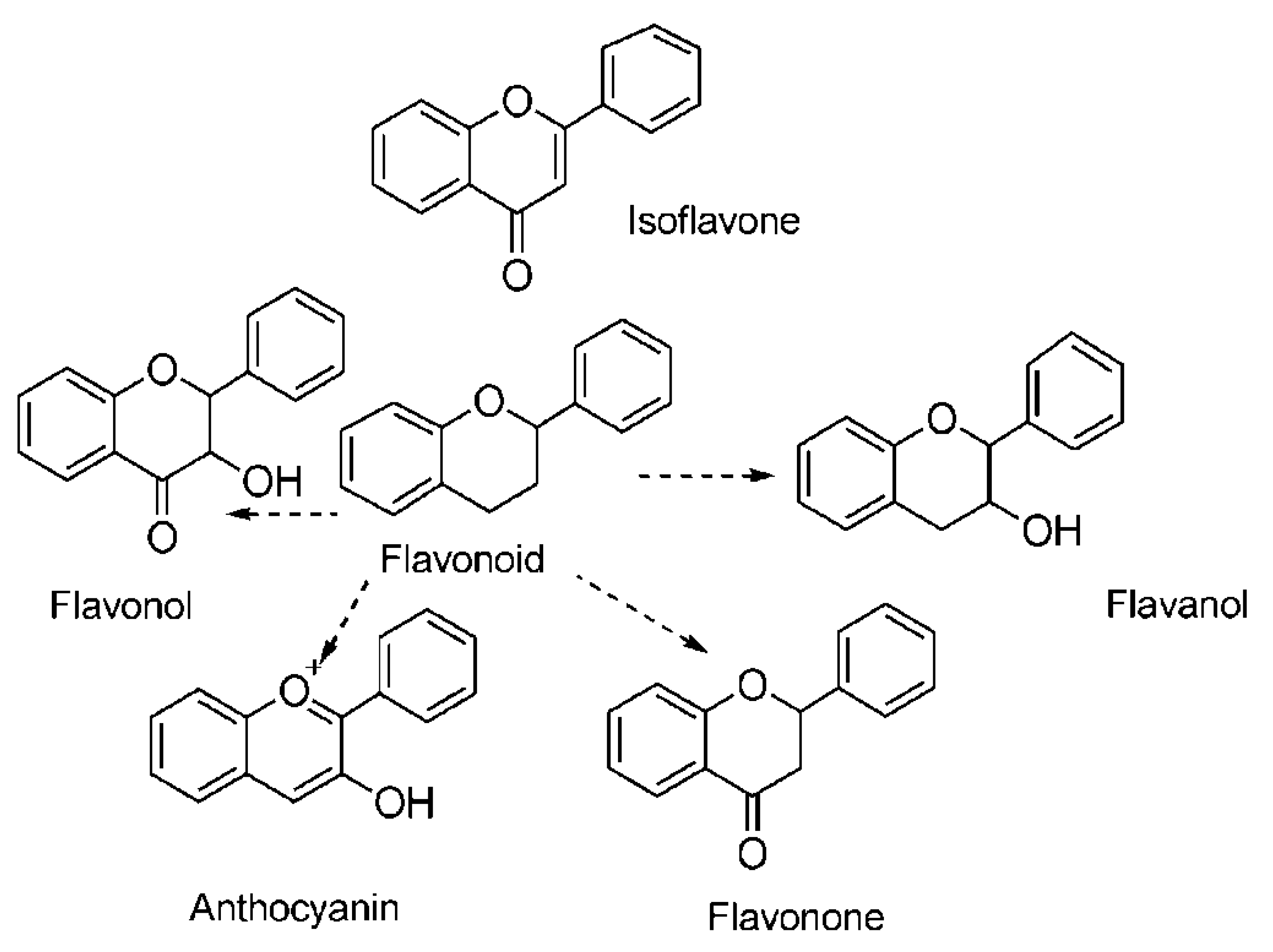
3.2. Total Flavonoid Content (TFC)
3.3. Carotenoids
3.4. DPPH Radical Scavenging Activity
4. Nutrition of TP on Poultry
5. Nutrition of TP on Swine
6. Nutrition of TP on Ruminants
7. Nutrition of TP on Rabbits
8. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Statistics, F.G.F. What Is the Global Feed Industry. International Feed Industry Federation Factsheet; International Feed Industry Federation (IFIF): Wiehl, Germany, 2019; Available online: https://ifif.org/wp-content/uploads/2019/06/IFIF-Fact-Sheet-October-11th-2019.pdf (accessed on 11 October 2019).
- Van, D.M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar]
- McLeod, A. World Livestock 2011-Livestock in Food Security; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2011. [Google Scholar]
- Lu, S.; Taethaisong, N.; Meethip, W.; Surakhunthod, J.; Sinpru, B.; Sroichak, T.; Archa, P.; Thongpea, S.; Paengkoum, S.; Purba, R.A.P.; et al. Nutritional Composition of Black Soldier Fly Larvae (Hermetia illucens L.) and Its Potential Uses as Alternative Protein Sources in Animal Diets: A Review. Insects 2022, 13, 831. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Islam, M.; Shehata, A.A.; Basiouni, S.; Hafez, H.M.; Azhar, E.I.; Khafaga, A.F.; Bovera, F.; Attia, Y.A. Influence of COVID-19 on the sustainability of livestock performance and welfare on a global scale. Trop. Anim. Health Prod. 2022, 54, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, X.; Liu, Z.; Huang, J.; Liu, Z. Application of fermented liquid feed in pig production. Chin. J. Anim. Nutr. 2018, 30, 4312–4319. [Google Scholar]
- Sogari, G.; Amato, M.; Biasato, I.; Chiesa, S.; Gasco, L. The potential role of insects as feed: A multi-perspective review. Animals 2019, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Coelho, M.; Pereira, R.; Rodrigues, A.S.; Teixeira, J.A.; Pintado, M.E. Extraction of tomato by-products’ bioactive compounds using ohmic technology. Food Bioprod. Process. 2019, 117, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Wang, J.; Gao, R.; Ye, F.; Zhao, G. Sustainable valorisation of tomato pomace: A comprehensive review. Trends Food Sci. Technol. 2019, 86, 172–187. [Google Scholar] [CrossRef]
- Li, J.; Yang, Z.; Zhang, Y.; Gao, B.; Niu, Y.; Yu, L.L. The structural and functional characteristics of soluble dietary fibers modified from tomato pomace with increased content of lycopene. Food Chem. 2022, 382, 132333. [Google Scholar] [CrossRef]
- Mildner-Szkudlarz, S.; Bajerska, J.; Górnaś, P.; Segliņa, D.; Pilarska, A.; Jesionowski, T. Physical and bioactive properties of muffins enriched with raspberry and cranberry pomace powder: A promising application of fruit by-products rich in biocompounds. Plant Foods Hum. Nutr. 2016, 71, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Pinela, J.; Montoya, C.; Carvalho, A.M.; Martins, V.; Rocha, F.; Barata, A.M.; Barros, L.; Ferreira, I.C. Phenolic composition and antioxidant properties of ex-situ conserved tomato (Solanum lycopersicum L.) germplasm. Food Res. Int. 2019, 125, 108545. [Google Scholar] [CrossRef] [Green Version]
- Szabo, K.; Cătoi, A.F.; Vodnar, D.C. Bioactive compounds extracted from tomato processing by-products as a source of valuable nutrients. Plant Foods Hum. Nutr. 2018, 73, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Belović, M.; Torbica, A.; Lijaković, I.P.; Tomić, J.; Lončarević, I.; Petrović, J. Tomato pomace powder as a raw material for ketchup production. Food Biosci. 2018, 26, 193–199. [Google Scholar] [CrossRef]
- Correia, C.; Alfaia, C.; Madeira, M.; Lopes, P.; Matos, T.; Cunha, L.; Prates, J.; Freire, J. Dietary inclusion of tomato pomace improves meat oxidative stability of young pigs. J. Anim. Physiol. Anim. Nutr. 2017, 101, 1215–1226. [Google Scholar] [CrossRef] [PubMed]
- Caluya, R.; Sair, R.; Balneg, B. Fresh tomato pomace (FTP) as good feed for growing and fattening pigs. Highlights’ 99 2000, 100, 143. [Google Scholar]
- Mizael, W.C.; Costa, R.G.; Rodrigo, B.C.G.; Ramos, C.F.F.; Ribeiro, N.L.; Lima, A.; Domínguez, R.; Lorenzo, J.M. Effect of the use of tomato pomace on feeding and performance of lactating goats. Animals 2020, 10, 1574. [Google Scholar] [CrossRef]
- Mohammed, L.S.; Sallam, E.A.; Edris, S.N.; Khalifa, O.A.; Soliman, M.M.; Shehata, S.F. Growth performance, economic efficiency, meat quality, and gene expression in two broiler breeds fed different levels of tomato pomace. Vet. Res. Commun. 2021, 45, 381–397. [Google Scholar] [CrossRef]
- Al-Wandawi, H.; Abdul-Rahman, M.; Al-Shaikhly, K. Tomato processing wastes as essential raw materials source. J. Agric. Food Chem. 1985, 33, 804–807. [Google Scholar] [CrossRef]
- Poojary, M.M.; Passamonti, P. Extraction of lycopene from tomato processing waste: Kinetics and modelling. Food Chem. 2015, 173, 943–950. [Google Scholar] [CrossRef]
- Ghuttora, N. Increase the Usage of Biopolymers and Biodegradable Polymers for Sustainable Environment. 2016. Available online: https://urn.fi/URN:NBN:fi:amk-201701131346 (accessed on 1 October 2022).
- Strati, I.; Oreopoulou, V. Recovery of carotenoids from tomato processing by-products—A review. Food Res. Int. 2014, 65, 311–321. [Google Scholar] [CrossRef]
- Kalogeropoulos, N.; Chiou, A.; Pyriochou, V.; Peristeraki, A.; Karathanos, V.T. Bioactive phytochemicals in industrial tomatoes and their processing byproducts. LWT Food Sci. Technol. 2012, 49, 213–216. [Google Scholar] [CrossRef]
- Nagarajan, J.; Pui, K.H.; Krishnamurthy, N.P.; Ramakrishnan, N.R.; Aldawoud, T.; Galanakis, C.M.; Wei, O.C. Extraction of carotenoids from tomato pomace via water-induced hydrocolloidal complexation. Biomolecules 2020, 10, 1019. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, E.; Trostchansky, A.; Reguengo, L.M.; Junior, M.R.; Palomo, I. Antiplatelet effects of bioactive compounds present in tomato pomace. Curr. Drug Targets 2021, 22, 1716–1724. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.U.; Khan, A.; Muhammad, M.D.; Naz, S. Tomato pomace waste as safe feed additive for poultry health and production—A review. Ann. Anim. Sci. 2022, 4, 2300–8733. [Google Scholar] [CrossRef]
- Purba, R.A.P.; Paengkoum, P.; Paengkoum, S. The links between supplementary tannin levels and conjugated linoleic acid (CLA) formation in ruminants: A systematic review and meta-analysis. PLoS ONE 2020, 15, e0216187. [Google Scholar] [CrossRef] [Green Version]
- Torbica, A.; Belović, M.; Mastilović, J.; Kevrešan, Ž.; Pestorić, M.; Škrobot, D.; Hadnađev, T.D. Nutritional, rheological, and sensory evaluation of tomato ketchup with increased content of natural fibres made from fresh tomato pomace. Food Bioprod. Process. 2016, 98, 299–309. [Google Scholar] [CrossRef]
- Denek, N.; Can, A. Feeding value of wet tomato pomace ensiled with wheat straw and wheat grain for Awassi sheep. Small Rumin. Res. 2006, 65, 260–265. [Google Scholar] [CrossRef]
- Shao, D.; Atungulu, G.G.; Pan, Z.; Yue, T.; Zhang, A.; Fan, Z. Characteristics of isolation and functionality of protein from tomato pomace produced with different industrial processing methods. Food Bioprocess Technol. 2014, 7, 532–541. [Google Scholar] [CrossRef]
- Azabou, S.; Louati, I.; Ben, T.F.; Nasri, M.; Mechichi, T. Towards sustainable management of tomato pomace through the recovery of valuable compounds and sequential production of low-cost biosorbent. Environ. Sci. Pollut. Res. 2020, 27, 39402–39412. [Google Scholar] [CrossRef]
- Valenti, B.; Luciano, G.; Pauselli, M.; Mattioli, S.; Biondi, L.; Priolo, A.; Natalello, A.; Morbidini, L.; Lanza, M. Dried tomato pomace supplementation to reduce lamb concentrate intake: Effects on growth performance and meat quality. Meat Sci. 2018, 145, 63–70. [Google Scholar] [CrossRef]
- Shao, D.; Bartley, G.E.; Yokoyama, W.; Pan, Z.; Zhang, H.; Zhang, A. Plasma and hepatic cholesterol-lowering effects of tomato pomace, tomato seed oil and defatted tomato seed in hamsters fed with high-fat diets. Food Chem. 2013, 139, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Rehal, J.K.; Aggarwal, P.; Dhaliwal, I.; Sharma, M.; Kaushik, P. A Tomato Pomace Enriched Gluten-Free Ready-to-Cook Snack’s Nutritional Profile, Quality, and Shelf Life Evaluation. Horticulturae 2022, 8, 403. [Google Scholar] [CrossRef]
- Ayhan, V.; Aktan, S. Using possibilities of dried tomato pomace in broiler chicken diets. Hayvansal ÜRetim 2004, 45, 19–22. [Google Scholar]
- Razzaghi, A.; Naserian, A.A.; Valizadeh, R.; Ebrahimi, S.H.; Khorrami, B.; Malekkhahi, M.; Khiaosa-Ard, R. Pomegranate seed pulp, pistachio hulls, and tomato pomace as replacement of wheat bran increased milk conjugated linoleic acid concentrations without adverse effects on ruminal fermentation and performance of Saanen dairy goats. Anim. Feed. Sci. Technol. 2015, 210, 46–55. [Google Scholar] [CrossRef]
- Herrera, P.G.; Sánchez-Mata, M.; Cámara, M. Nutritional characterization of tomato fiber as a useful ingredient for food industry. Innov. Food Sci. Emerg. Technol. 2010, 11, 707–711. [Google Scholar] [CrossRef]
- Nour, V.; Panaite, T.D.; Ropota, M.; Turcu, R.; Trandafir, I.; Corbu, A.R. Nutritional and bioactive compounds in dried tomato processing waste. CyTA J. Food 2018, 16, 222–229. [Google Scholar] [CrossRef]
- Pinela, J.; Barros, L.; Carvalho, A.M.; Ferreira, I.C. Nutritional composition and antioxidant activity of four tomato (Lycopersicon esculentum L.) farmer’varieties in Northeastern Portugal homegardens. Food Chem. Toxicol. 2012, 50, 829–834. [Google Scholar] [CrossRef]
- Elbadrawy, E.; Sello, A. Evaluation of nutritional value and antioxidant activity of tomato peel extracts. Arab. J. Chem. 2016, 9, S1010–S1018. [Google Scholar] [CrossRef] [Green Version]
- Namir, M.; Suleiman, A.R.; Hassanien, M.F.R. Characterization and functionality of alcohol insoluble solids from tomato pomace as fat substitute in low fat cake. J. Food Meas. Charact. 2015, 9, 557–563. [Google Scholar] [CrossRef]
- Navarro-González, I.; García-Valverde, V.; García-Alonso, J.; Periago, M.J. Chemical profile, functional and antioxidant properties of tomato peel fiber. Food Res. Int. 2011, 44, 1528–1535. [Google Scholar] [CrossRef]
- Westphal, A.; Bauerfeind, J.; Rohrer, C.; Böhm, V. Analytical characterisation of the seeds of two tomato varieties as a basis for recycling of waste materials in the food industry. Eur. Food Res. Technol. 2014, 239, 613–620. [Google Scholar] [CrossRef]
- Isik, F.; Yapar, A. Effect of tomato seed supplementation on chemical and nutritional properties of tarhana. J. Food Meas. Charact. 2017, 11, 667–674. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, R.; Sharopov, F.; Namiesnik, J.; Roointan, A.; Kamle, M.; Kumar, P.; Martins, N.; Sharifi-Rad, J. Beneficial effects and potential risks of tomato consumption for human health: An overview. Nutrition 2019, 62, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Namir, M.; Siliha, H.; Ramadan, M.F. Fiber pectin from tomato pomace: Characteristics, functional properties and application in low-fat beef burger. J. Food Meas. Charact. 2015, 9, 305–312. [Google Scholar] [CrossRef]
- Biondi, L.; Luciano, G.; Cutello, D.; Natalello, A.; Mattioli, S.; Priolo, A.; Lanza, M.; Morbidini, L.; Gallo, A.; Valenti, B. Meat quality from pigs fed tomato processing waste. Meat Sci. 2020, 159, 107940. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient Requirements of Poultry: 1994; National Academies Press: Washington, DC, USA, 1994. [Google Scholar]
- Applegate, T.J.; Angel, R. Nutrient requirements of poultry publication: History and need for an update. J. Appl. Poult. Res. 2014, 23, 567–575. [Google Scholar] [CrossRef]
- Halliwell, B. Vitamin C and genomic stability. Mutat. Res. 2001, 475, 29–35. [Google Scholar] [CrossRef]
- Mayne, S.T. Antioxidant nutrients and chronic disease: Use of biomarkers of exposure and oxidative stress status in epidemiologic research. J. Nutr. 2003, 133, 933S–940S. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Çakmakçı, S.; Topdaş, E.F.; Kalın, P.; Han, H.; Şekerci, P.; Köse, L.; Gülçin, İ. Antioxidant capacity and functionality of oleaster (E laeagnus angustifolia L.) flour and crust in a new kind of fruity ice cream. Int. J. Food Sci. Technol. 2015, 50, 472–481. [Google Scholar] [CrossRef]
- Purba, R.A.P.; Paengkoum, S.; Yuangklang, C.; Paengkoum, P. Flavonoids and their aromatic derivatives in Piper betle powder promote in vitro methane mitigation in a variety of diets. Ciência Agrotecnologia 2020, 44, e012420. [Google Scholar] [CrossRef]
- Bursal, E.; Köksal, E.; Gülçin, İ.; Bilsel, G.; Gören, A.C. Antioxidant activity and polyphenol content of cherry stem (Cerasus avium L.) determined by LC–MS/MS. Food Res. Int. 2013, 51, 66–74. [Google Scholar] [CrossRef]
- Sharma, A.; Bhardwaj, G.; Gaba, J.; Cannoo, D.S. Natural Antioxidants: Assays and Extraction Methods/Solvents Used for Their Isolation. In Antioxidants in Fruits: Properties and Health Benefits; Springer: Berlin/Heidelberg, Germany, 2020; pp. 1–33. [Google Scholar]
- Anwar, H.; Hussain, G.; Mustafa, I. Antioxidants from natural sources. Antioxid. Foods Its Appl. 2018, 10, 3–28. [Google Scholar]
- Lattanzio, V.; Kroon, P.A.; Quideau, S.; Treutter, D. Plant phenolics—Secondary metabolites with diverse functions. Recent Adv. Polyphen. Res. 2009, 1, 1–35. [Google Scholar]
- Jain, C.; Khatana, S.; Vijayvergia, R. Bioactivity of secondary metabolites of various plants: A review. Int. J. Pharm. Sci. Res. 2019, 10, 494–504. [Google Scholar]
- Purba, R.A.P.; Paengkoum, P. Bioanalytical HPLC method of Piper betle L. for quantifying phenolic compound, water-soluble vitamin, and essential oil in five different solvent extracts. J. Appl. Pharm. Sci. 2019, 9, 033–039. [Google Scholar]
- Fernández-Martínez, A.; Charlet, L. Selenium environmental cycling and bioavailability: A structural chemist point of view. Rev. Environ. Sci. Bio/Technol. 2009, 8, 81–110. [Google Scholar] [CrossRef]
- Godswill, A.G.; Somtochukwu, I.V.; Ikechukwu, A.O.; Kate, E.C. Health benefits of micronutrients (vitamins and minerals) and their associated deficiency diseases: A systematic review. Int. J. Food Sci. 2020, 3, 1–32. [Google Scholar] [CrossRef]
- Diplock, A.; Charuleux, J.L.; Crozier-Willi, G.; Kok, F.; Rice-Evans, C.; Roberfroid, M.; Stahl, W.; Vina-Ribes, J. Functional food science and defence against reactive oxidative species. Br. J. Nutr. 1998, 80, S77–S112. [Google Scholar] [CrossRef] [Green Version]
- Waterman, P.G.; Mole, S. Analysis of Phenolic Plant Metabolites; Blackwell Scientific: Washington, DC, USA, 1994. [Google Scholar]
- Eberhardt, M.V.; Lee, C.Y.; Liu, R.H. Antioxidant activity of fresh apples. Nature 2000, 405, 903–904. [Google Scholar] [CrossRef]
- Tian, X.; Lu, Q.; Paengkoum, P.; Paengkoum, S. Effect of purple corn pigment on change of anthocyanin composition and unsaturated fatty acids during milk storage. J. Dairy Sci. 2020, 103, 7808–7812. [Google Scholar] [CrossRef] [PubMed]
- Issaoui, M.; Delgado, A.M.; Caruso, G.; Micali, M.; Barbera, M.; Atrous, H.; Ouslati, A.; Chammem, N. Phenols, flavors, and the mediterranean diet. J. AOAC Int. 2020, 103, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Petlum, A.; Paengkoum, P.; Liang, J.; Vasupen, K.; Paengkoum, S. Molecular weight of condensed tannins of some tropical feed-leaves and their effect on in vitro gas and methane production. Anim. Prod. Sci. 2019, 59, 2154–2160. [Google Scholar] [CrossRef]
- Ban, C.; Paengkoum, S.; Yang, S.; Tian, X.; Thongpea, S.; Purba, R.A.P.; Paengkoum, P. Feeding meat goats mangosteen (Garcinia mangostana L.) peel rich in condensed tannins, flavonoids, and cinnamic acid improves growth performance and plasma antioxidant activity under tropical conditions. J. Appl. Anim. Res. 2022, 50, 307–315. [Google Scholar] [CrossRef]
- Priftis, A.; Soursou, V.; Makiou, A.S.; Tekos, F.; Veskoukis, A.S.; Tsantarliotou, M.P.; Taitzoglou, I.A.; Kouretas, D. A lightly roasted coffee extract improves blood and tissue redox status in rats through enhancement of GSH biosynthesis. Food Chem. Toxicol. 2019, 125, 305–312. [Google Scholar] [CrossRef]
- Purba, R.; Yuangklang, C.; Paengkoum, S.; Paengkoum, P. Milk fatty acid composition, rumen microbial population and animal performance in response to diets rich in linoleic acid supplemented with Piper betle leaves in Saanen goats. Anim. Prod. Sci. 2020, 62, 1391–1401. [Google Scholar] [CrossRef]
- Laudadio, V.; Ceci, E.; Lastella, N.; Tufarelli, V. Dietary high-polyphenols extra-virgin olive oil is effective in reducing cholesterol content in eggs. Lipids Health Dis. 2015, 14, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Shen, T.; Lou, H. Dietary polyphenols and their biological significance. Int. J. Mol. Sci. 2007, 8, 950–988. [Google Scholar] [CrossRef]
- Kikusato, M. Phytobiotics to improve health and production of broiler chickens: Functions beyond the antioxidant activity. Anim. Biosci. 2021, 34, 345. [Google Scholar] [CrossRef]
- Purba, R.A.P.; Paengkoum, S.; Yuangklang, C.; Paengkoum, P.; Salem, A.Z.M.; Juan Boo, L. Mammary gene expressions and oxidative indicators in ruminal fluid, blood, milk, and mammary tissue of dairy goats fed a total mixed ration containing piper meal (Piper betle L.). Ital. J. Anim. Sci. 2022, 21, 129–141. [Google Scholar] [CrossRef]
- Patias, N.S.; Gindri Sinhorin, V.D.; de Moura, F.R.; da Cunha, A.P.S.; da Silva, L.R.R.; da Costa, T.B.; da Costa, R.J.; Cavalheiro, L.; de Campos, B.R.; Sinhorin, A.P. Identification of Flavonoids by LC-MS/MS in Leaves Extract From Protium heptaphyllum (Aubl.) March and Antioxidant Activity in Mice. Nat. Prod. J. 2021, 11, 715–727. [Google Scholar] [CrossRef]
- Vecchia, C.A.; Locateli, G.; Serpa, P.Z.; Bianchin, G.D.; Ernetti, J.; Miorando, D.; Zanatta, M.E.D.; Kaio, S.N.R.; Wildner, S.M.; Gutiérrez, M.V. Sonchus oleraceus L. Promotes Gastroprotection in Rodents via Antioxidant, Anti-Inflammatory, and Antisecretory Activities. Evid. Based Complement. Altern. Med. 2022, 2022, 7413231. [Google Scholar] [CrossRef] [PubMed]
- Durand, D.; Damon, M.; Gobert, M. Oxidative stress in farm animals: General aspects. Cahiers Nutrition Diététique 2013, 48, 218–224. [Google Scholar] [CrossRef]
- Landete, J. Dietary intake of natural antioxidants: Vitamins and polyphenols. Crit. Rev. Food Sci. Nutr. 2013, 53, 706–721. [Google Scholar] [CrossRef] [PubMed]
- Purba, R.A.P.; Suong, N.T.M.; Paengkoum, S.; Schonewille, J.T.; Paengkoum, P. Dietary inclusion of anthocyanin-rich black cane silage treated with ferrous sulfate heptahydrate reduces oxidative stress and promotes tender meat production in goats. Front. Vet. Sci. 2022, 9, 969321. [Google Scholar] [CrossRef]
- Lipiński, K.; Mazur, M.; Antoszkiewicz, Z.; Purwin, C. Polyphenols in monogastric nutrition—A review. Ann. Anim. Sci. 2017, 17, 41–58. [Google Scholar] [CrossRef] [Green Version]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Ahmed, N.A.; Radwan, N.M.; Aboul Ezz, H.S.; Salama, N.A. The antioxidant effect of Green Tea Mega EGCG against electromagnetic radiation-induced oxidative stress in the hippocampus and striatum of rats. Electromagn. Biol. Med. 2017, 36, 63–73. [Google Scholar] [CrossRef]
- Negishi, H.; Xu, J.W.; Ikeda, K.; Njelekela, M.; Nara, Y.; Yamori, Y. Black and green tea polyphenols attenuate blood pressure increases in stroke-prone spontaneously hypertensive rats. J. Nutr. 2004, 134, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef]
- Yiannakopoulou, E.C. Targeting oxidative stress response by green tea polyphenols: Clinical implications. Free. Radic. Res. 2013, 47, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Do, Q.D.; Angkawijaya, A.E.; Tran-Nguyen, P.L.; Huynh, L.H.; Soetaredjo, F.E.; Ismadji, S.; Ju, Y.H. Effect of extraction solvent on total phenol content, total flavonoid content, and antioxidant activity of Limnophila aromatica. J. Food Drug Anal. 2014, 22, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Ghafar, F.; Nazrin, T.; Salleh, M.; Hadi, N.N.; Ahmad, N.; Hamzah, A.A.; Yusof, Z.A.M.; Azman, I.N. Total phenolic content and total flavonoid content in moringa oleifera seed. Galeri Waris. Sains 2017, 1, 23–25. [Google Scholar] [CrossRef]
- Matić, P.; Sabljić, M.; Jakobek, L. Validation of spectrophotometric methods for the determination of total polyphenol and total flavonoid content. J. AOAC Int. 2017, 100, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef] [PubMed]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [Green Version]
- Ballard, C.R.; Junior, M.R.M. Health benefits of flavonoids. In Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2019; pp. 185–201. [Google Scholar]
- Paengkoum, P. Effects of neem (Azadirachta indica) and leucaena (Leucaena leucocephala) fodders on digestibility, rumen fermentation and nitrogen balance of goats fed corn silage. J. Anim. Vet. Adv. 2010, 9, 883–886. [Google Scholar] [CrossRef]
- Suong, N.T.M.; Paengkoum, S.; Schonewille, J.T.; Purba, R.A.P.; Paengkoum, P. Growth performance, blood biochemical indices, rumen bacterial community, and carcass characteristics in goats fed anthocyanin-rich black cane silage. Front. Vet. Sci. 2022, 9, 880838. [Google Scholar] [CrossRef]
- Suong, N.T.M.; Paengkoum, S.; Purba, R.A.P.; Paengkoum, P. Optimizing Anthocyanin-Rich Black Cane (Saccharum sinensis Robx.) Silage for Ruminants Using Molasses and Iron Sulphate: A Sustainable Alternative. Fermentation 2022, 8, 248. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef] [PubMed]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Shi, Z.; Yang, X.; Cai, Z.; Wang, L.; Wu, X.; Ye, A.; Jiang, J. Hypolipidemic effect of pure total flavonoids from peel of Citrus (PTFC) on hamsters of hyperlipidemia and its potential mechanism. Exp. Gerontol. 2020, 130, 110786. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; Van, N.E.; Van, H.D.E.; Boelens, P.G.; Van, N.K.; Van, L.P.A. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Important flavonoids and their role as a therapeutic agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef]
- Paengkoum, S.; Petlum, A.; Purba, R.A.P.; Paengkoum, P. Protein-binding affinity of various condensed tannin molecular weights from tropical leaf peel. J. Appl. Pharm. Sci. 2021, 11, 114–120. [Google Scholar]
- Langi, P.; Kiokias, S.; Varzakas, T.; Proestos, C. Carotenoids: From plants to food and feed industries. Microb. Carotenoids 2018, 1852, 57–71. [Google Scholar]
- Lyu, X.; Lyu, Y.; Yu, H.; Chen, W.; Ye, L.; Yang, R. Biotechnological advances for improving natural pigment production: A state-of-the-art review. Bioresour. Bioprocess. 2022, 9, 1–38. [Google Scholar] [CrossRef]
- Aberoumand, A. A review article on edible pigments properties and sources as natural biocolorants in foodstuff and food industry. World J. Dairy Food Sci. 2011, 6, 71–78. [Google Scholar]
- Kirti, K.; Amita, S.; Priti, S.; Jyoti, S. Colorful world of microbes: Carotenoids and their applications. Adv. Biol. 2014, 2014, 837891. [Google Scholar] [CrossRef]
- Rodrigo-Baños, M.; Garbayo, I.; Vílchez, C.; Bonete, M.J.; Martínez-Espinosa, R.M. Carotenoids from Haloarchaea and their potential in biotechnology. Mar. Drugs 2015, 13, 5508–5532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imran, M.; Ghorat, F.; Ul-Haq, I.; Ur-Rehman, H.; Aslam, F.; Heydari, M.; Shariati, M.A.; Okuskhanova, E.; Yessimbekov, Z.; Thiruvengadam, M. Lycopene as a natural antioxidant used to prevent human health disorders. Antioxidants 2020, 9, 706. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, N.P.; Holzapfel, B.M.; Champ, S.; Feldthusen, J.; Clements, J.; Hutmacher, D.W. The potential role of lycopene for the prevention and therapy of prostate cancer: From molecular mechanisms to clinical evidence. Int. J. Mol. Sci. 2013, 14, 14620–14646. [Google Scholar] [CrossRef] [Green Version]
- Calvo, M.; García, M.L.; Selgas, M.D. Dry fermented sausages enriched with lycopene from tomato peel. Meat Sci. 2008, 80, 167–172. [Google Scholar] [CrossRef]
- Capanoglu, E.; Beekwilder, J.; Boyacioglu, D.; De Vos, R.C.; Hall, R.D. The effect of industrial food processing on potentially health-beneficial tomato antioxidants. Crit. Rev. Food Sci. Nutr. 2010, 50, 919–930. [Google Scholar] [CrossRef]
- Bose, K.; Agrawal, B. Effect of lycopene from cooked tomatoes on serum antioxidant enzymes, lipid peroxidation rate and lipid profile in coronary heart disease. Singap. Med. J. 2007, 48, 415. [Google Scholar]
- Przybylska, S. Lycopene—A bioactive carotenoid offering multiple health benefits: A review. Int. J. Food Sci. Technol. 2020, 55, 11–32. [Google Scholar] [CrossRef]
- Edge, R.; Truscott, T.G. Singlet oxygen and free radical reactions of retinoids and carotenoids—A review. Antioxidants 2018, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.H.; Park, M.K.; Kim, S.K.; Cho, Y.H. Antioxidant capacity and anti-inflammatory activity of lycopene in watermelon. Int. J. Food Sci. Technol. 2014, 49, 2083–2091. [Google Scholar] [CrossRef]
- Puah, B.P.; Jalil, J.; Attiq, A.; Kamisah, Y. New insights into molecular mechanism behind anti-cancer activities of lycopene. Molecules 2021, 26, 3888. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and chlorophylls as antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Prasad, K.N.; Kong, K.W.; Jiang, Y.; Ismail, A. Carotenoids and their isomers: Color pigments in fruits and vegetables. Molecules 2011, 16, 1710–1738. [Google Scholar] [CrossRef] [PubMed]
- Zia-Ul-Haq, M. Historical and introductory aspects of carotenoids. In Carotenoids: Structure and Function in the Human Body; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–42. [Google Scholar]
- Olmedilla-Alonso, B.; Benítez-González, A.M.; Estévez-Santiago, R.; Mapelli-Brahm, P.; Stinco, C.M.; Meléndez-Martínez, A.J. Assessment of Food Sources and the Intake of the Colourless Carotenoids Phytoene and Phytofluene in Spain. Nutrients 2021, 13, 4436. [Google Scholar] [CrossRef] [PubMed]
- Simkin, A.J. Carotenoids and apocarotenoids in planta: Their role in plant development, contribution to the flavour and aroma of fruits and flowers, and their nutraceutical benefits. Plants 2021, 10, 2321. [Google Scholar] [CrossRef]
- Boonlao, N.; Ruktanonchai, U.R.; Anal, A.K. Enhancing bioaccessibility and bioavailability of carotenoids using emulsion-based delivery systems. Colloids Surf. B Biointerfaces 2022, 209, 112211. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Mandić, A.I.; Bantis, F.; Böhm, V.; Borge, G.I.A.; Brnčić, M.; Bysted, A.; Cano, M.P.; Dias, M.G.; Elgersma, A. A comprehensive review on carotenoids in foods and feeds: Status quo, applications, patents, and research needs. Crit. Rev. Food Sci. Nutr. 2022, 62, 1999–2049. [Google Scholar] [CrossRef]
- Ribeiro, D.; Freitas, M.; Silva, A.M.; Carvalho, F.; Fernandes, E. Antioxidant and pro-oxidant activities of carotenoids and their oxidation products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef]
- Shi, M.; Gu, J.; Wu, H.; Rauf, A.; Emran, T.B.; Khan, Z.; Mitra, S.; Aljohani, A.S.; Alhumaydhi, F.A.; Al-Awthan, Y.S. Phytochemicals, Nutrition, Metabolism, Bioavailability, and Health Benefits in Lettuce—A Comprehensive Review. Antioxidants 2022, 11, 1158. [Google Scholar] [CrossRef]
- Nićiforović, N.; Mihailović, V.; Mašković, P.; Solujić, S.; Stojković, A.; Muratspahić, D.P. Antioxidant activity of selected plant species; potential new sources of natural antioxidants. Food Chem. Toxicol. 2010, 48, 3125–3130. [Google Scholar] [CrossRef]
- Kedare, S.B.; Singh, R. Genesis and development of DPPH method of antioxidant assay. J. Food Sci. Technol. 2011, 48, 412–422. [Google Scholar] [CrossRef] [Green Version]
- Habu, J.B.; Ibeh, B.O. In vitro antioxidant capacity and free radical scavenging evaluation of active metabolite constituents of Newbouldia laevis ethanolic leaf extract. Biol. Res. 2015, 48, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Niu, F.; Hu, D.; Gu, F.; Du, Y.; Zhang, B.; Ma, S.; Pan, W. Preparation of ultra-long stable ovalbumin/sodium carboxymethylcellulose nanoparticle and loading properties of curcumin. Carbohydr. Polym. 2021, 271, 118451. [Google Scholar] [CrossRef] [PubMed]
- Hioe, J.; Šakić, D.; Vrček, V.; Zipse, H. The stability of nitrogen-centered radicals. Org. Biomol. Chem. 2015, 13, 157–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gülçin, İ.; Elmastaş, M.; Aboul-Enein, H.Y. Antioxidant activity of clove oil–A powerful antioxidant source. Arab. J. Chem. 2012, 5, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Islam, M.; Biswas, M.; Khurshid, A.A. In vitro antioxidant and free radical scavenging activity of different parts of Tabebuia pallida growing in Bangladesh. BMC Res. Notes 2015, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbasi-Parizad, P.; De Nisi, P.; Adani, F.; Pepé Sciarria, T.; Squillace, P.; Scarafoni, A.; Iametti, S.; Scaglia, B. Antioxidant and anti-inflammatory activities of the crude extracts of raw and fermented tomato pomace and their correlations with aglycate-polyphenols. Antioxidants 2020, 9, 179. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Tomar, M.; Bhuyan, D.J.; Punia, S.; Grasso, S.; Sa, A.G.A.; Carciofi, B.A.M.; Arrutia, F.; Changan, S.; Singh, S. Tomato (Solanum lycopersicum L.) seed: A review on bioactives and biomedical activities. Biomed. Pharmacother. 2021, 142, 112018. [Google Scholar] [CrossRef]
- Reda, F.M.; Madkour, M.; Abd El-Azeem, N.; Aboelazab, O.; Ahmed, S.Y.; Alagawany, M. Tomato pomace as a nontraditional feedstuff: Productive and reproductive performance, digestive enzymes, blood metabolites, and the deposition of carotenoids into egg yolk in quail breeders. Poult. Sci. 2022, 101, 101730. [Google Scholar] [CrossRef]
- Hosseini-Vashan, S.; Golian, A.; Yaghobfar, A. Growth, immune, antioxidant, and bone responses of heat stress-exposed broilers fed diets supplemented with tomato pomace. Int. J. Biometeorol. 2016, 60, 1183–1192. [Google Scholar] [CrossRef]
- Nemli, E.; Ozakdogan, S.; Tomas, M.; McClements, D.J.; Capanoglu, E. Increasing the bioaccessibility of antioxidants in tomato pomace using excipient emulsions. Food Biophys. 2021, 16, 355–364. [Google Scholar] [CrossRef]
- Niu, Z.; Liu, F.; Yan, Q.; Li, L. Effects of different levels of selenium on growth performance and immunocompetence of broilers under heat stress. Arch. Anim. Nutr. 2009, 63, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Habibian, M.; Ghazi, S.; Moeini, M.M.; Abdolmohammadi, A. Effects of dietary selenium and vitamin E on immune response and biological blood parameters of broilers reared under thermoneutral or heat stress conditions. Int. J. Biometeorol. 2014, 58, 741–752. [Google Scholar] [CrossRef]
- Rezaeipour, V.; Boldaji, F.; Dastar, B.; Yaghoubfar, A.; Gheysari, A. Determination of apparent nutrients digestibility and metabolizable energy of diets with different inclusion of Dried Tomato Pomace in broiler chickens. J. Agric. Sci. Nat. Resour. 2009, 16, 90–102. [Google Scholar]
- Sogi, D.; Sidhu, J.; Arora, M.; Garg, S.; Bawa, A. Effect of tomato seed meal supplementation on the dough and bread characteristics of wheat (PBW 343) flour. Int. J. Food Prop. 2002, 5, 563–571. [Google Scholar] [CrossRef]
- Yolao, C.; Yammuen-art, S. Use of tomato pomace as antioxidant on growth performance of broilers under stress condition. Vet. Integr. Sci. 2016, 14, 63–71. [Google Scholar]
- Omar, M.A.; Abdel-Hamid, T.M.; Esam, S.; Omar, A.E. Growth and economic performance of using Dried Tomato Pomace for Mallard Ducks. Slov. Vet. Res. 2019, 56, 699–706. [Google Scholar] [CrossRef] [Green Version]
- Yalcin, S.; Siegel, P. Exposure to cold or heat during incubation on developmental stability of broiler embryos. Poult. Sci. 2003, 82, 1388–1392. [Google Scholar] [CrossRef]
- Caldwell, P.J.; Cornwell, G.W. Incubation behavior and temperatures of the mallard duck. Auk 1975, 92, 706–731. [Google Scholar] [CrossRef] [Green Version]
- An, B.K.; Kim, D.H.; Joo, W.D.; Kang, C.W.; Lee, K.W. Effects of lycopene and tomato paste on oxidative stability and fatty acid composition of fresh belly meat in finishing pigs. Ital. J. Anim. Sci. 2019, 18, 630–635. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.; Fan, Y.; Zhu, M.; Yang, Y.; Ma, Y. Energy content, nutrient digestibility coefficient, growth performance and serum parameters of pigs fed diets containing tomato pomace. J. Appl. Anim. Res. 2018, 46, 1483–1489. [Google Scholar] [CrossRef] [Green Version]
- Fachinello, M.R.; Gasparino, E.; Partyka, A.V.S.; de Souza, K.A.; Castilha, L.D.; Huepa, L.M.D.; Ferreira, L.F.M.; Pozza, P.C. Dietary lycopene alters the expression of antioxidant enzymes and modulates the blood lipid profile of pigs. Anim. Prod. Sci. 2020, 60, 806–814. [Google Scholar] [CrossRef]
- Meng, Q.; Zhang, Y.; Li, J.; Shi, B.; Ma, Q.; Shan, A. Lycopene Affects Intestinal Barrier Function and the Gut Microbiota in Weaned Piglets Via Antioxidant Signaling Regulation. J. Nutr. 2022, 152, 2396–2408. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Okawara, S.; Bhuiyan, M.; Fukui, Y. Effect of lycopene on cytoplasmic maturation of porcine oocytes in vitro. Reprod. Domest. Anim. 2010, 45, 838–845. [Google Scholar]
- Wen, W.; Chen, X.; Huang, Z.; Chen, D.; Yu, B.; He, J.; Luo, Y.; Yan, H.; Chen, H.; Zheng, P. Dietary lycopene supplementation improves meat quality, antioxidant capacity and skeletal muscle fiber type transformation in finishing pigs. Anim. Nutr. 2022, 8, 256–264. [Google Scholar] [CrossRef]
- Chatzifotis, S.; Pavlidis, M.; Jimeno, C.D.; Vardanis, G.; Sterioti, A.; Divanach, P. The effect of different carotenoid sources on skin coloration of cultured red porgy (Pagrus pagrus). Aquac. Res. 2005, 36, 1517–1525. [Google Scholar] [CrossRef]
- Abdullahzadeh, F. The effect of tomato pomace on carcass traits, blood metabolites and fleece characteristic of growing Markhoz goat. J. Am. Sci. 2012, 8, 848–852. [Google Scholar]
- Omer, H.; Abdel-Magid, S.S. Incorporation of dried tomato pomace in growing sheep rations. Glob. Vet. 2015, 14, 1–16. [Google Scholar]
- Abbeddou, S.; Rischkowsky, B.; Hilali, M.E.-D.; Haylani, M.; Hess, H.D.; Kreuzer, M. Supplementing diets of Awassi ewes with olive cake and tomato pomace: On-farm recovery of effects on yield, composition and fatty acid profile of the milk. Trop. Anim. Health Prod. 2015, 47, 145–152. [Google Scholar] [CrossRef]
- Romero-Huelva, M.; Ramos-Morales, E.; Molina-Alcaide, E. Nutrient utilization, ruminal fermentation, microbial abundances, and milk yield and composition in dairy goats fed diets including tomato and cucumber waste fruits. J. Dairy Sci. 2012, 95, 6015–6026. [Google Scholar] [CrossRef] [Green Version]
- Tuoxunjiang, H.; Yimamu, A.; Li, X.; Maimaiti, R.; Wang, Y. Effect of ensiled tomato pomace on performance and antioxidant status in the peripartum dairy cow. J. Anim. Feed Sci. 2020, 29, 105–114. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, J.; Zhang, Y.; Wang, X.; Wang, R.; Li, J. Effects of tomato pomace fermentation feed on growth performance, milk composition and blood cell parameters for Xinjiang Brown cows. Xinjiang Agric. Sci. 2012, 49, 1546–1551. [Google Scholar]
- Tahmasbi, R.; Moghadam, H.; Naserian, A.; Saremi, B. Chemical composition of mixed corn plant and tomato pomace silage and its effect on Holstein dairy cattle performance. In New Dimensions and Challenges for Sustainable Livestock Farming: Proceedings of the 11th Animal Science Congress, Kuala Lumpur, Malaysia, 5–9 September 2004; Malaysian Society of Animal Production: Darul Ehsan, Malaysia; p. 2004.
- Yuangklang, C.; Vasupen, K.; Wongsuthavas, S.; Panyakaew, P.; Alhaidary, A.; Mohamed, H.; Beynen, A. Growth performance in beef cattle fed rations containing dried tomato pomace. J. Anim. Vet. Adv. 2010, 9, 2261–2264. [Google Scholar] [CrossRef]
- Yuangklang, C.; Vasupen, K.; Wongsuthavas, S.; Bureenok, S.; Panyakaew, P.; Alhaidary, A.; Mohamed, H.; Beynen, A. Effect of replacement of soybean meal by dried tomato pomace on rumen fermentation and nitrogen metabolism in beef cattle. Am. J. Agric. Biol. Sci. 2010, 5, 256–260. [Google Scholar] [CrossRef] [Green Version]
- Yuangklang, C.; Vasupen, K.; Srenanul, P.; Wongsutthavas, S.; Mitchaothai, J. Effect of utilization of dried tomato pomace as roughage source on feed intake, rumen fermentation and blood metabolites in beef cattle. In Proceedings of the 44th Kasetsart University Annual Conference, Kasetsart, Thailand, 30 January–2 February 2006; pp. 158–166. [Google Scholar]
- Marcos, C.N.; de Evan, T.; Molina-Alcaide, E.; Carro, M. Nutritive value of tomato pomace for ruminants and its influence on in vitro methane production. Animals 2019, 9, 343. [Google Scholar] [CrossRef] [Green Version]
- Peiretti, P.; Gai, F.; Rotolo, L.; Brugiapaglia, A.; Gasco, L. Effects of tomato pomace supplementation on carcass characteristics and meat quality of fattening rabbits. Meat Sci. 2013, 95, 345–351. [Google Scholar] [CrossRef]
- Culler, M.D.; Inchingolo, R.; McClements, D.J.; Decker, E.A. Impact of polyunsaturated fatty acid dilution and antioxidant addition on lipid oxidation kinetics in oil/water emulsions. J. Agric. Food Chem. 2021, 69, 750–755. [Google Scholar] [CrossRef]
- Hassan, F.A.; Elkassas, N.; Salim, I.; El-Medany, S.; Aboelenin, S.M.; Shukry, M.; Taha, A.E.; Peris, S.; Soliman, M.; Mahrose, K. Impacts of dietary supplementations of orange peel and tomato pomace extracts as natural sources for ascorbic acid on growth performance, carcass characteristics, plasma biochemicals and antioxidant status of growing rabbits. Animals 2021, 11, 1688. [Google Scholar] [CrossRef]
- Hassan, F.; Abd-ElMola, L.; Mobarez, S.; Othman, D.; Zedan, A.; Mekawy, A.; Mansour, A.M.; Mahrose, K. Influence of tomato processing by-product extract as dietary supplementation on growth performance, carcass characteristics and antioxidant status of growing rabbits under high ambient temperature. Anim. Biotechnol. 2022, 1–10. [Google Scholar] [CrossRef]
- Mennani, A.; Arbouche, Y.; Arbouche, R.; Arbouche, F.; Ouzzir, L. Dehydrated tomato pulp in rabbit feed: Effects of incorporation rate on growth performance, carcass yield, meat quality and economic efficiency. J. Anim. Feed. Sci. 2021, 30, 271–278. [Google Scholar] [CrossRef]
- Elwan, H.A.; Elnesr, S.S.; Mohany, M.; Al-Rejaie, S.S. The effects of dietary tomato powder (Solanum lycopersicum L.) supplementation on the haematological, immunological, serum biochemical and antioxidant parameters of growing rabbits. J. Anim. Physiol. Anim. Nutr. 2019, 103, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Khadr, N.; Abdel-Fattah, F. Tomato Waste As An Unusual Feedstuff For Rabbits 2. Effect Of Tomato Waste On Productive Performance And Efficiency Of Rabbits. Zag. Vet. J. 2008, 36, 49–57. [Google Scholar]
- King, A.; Zeidler, G. Tomato pomace may be a good source of vitamin E in broiler diets. Calif. Agric. 2004, 58, 59–62. [Google Scholar] [CrossRef] [Green Version]
- Rengaraj, D.; Hong, Y.H. Effects of dietary vitamin E on fertility functions in poultry species. Int. J. Mol. Sci. 2015, 16, 9910–9921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Ratel, I. Impact of lycopene or folic acid treatment on semen quality, blood constituents and fertility of rabbit bucks. Egypt. J. Nutr. Feed. 2017, 20, 213–223. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Moisture Ash | 43.4 | 79.0 70.1 | 88.4 27.5 | 37.0 | 59.6 42.7 | 81.0 41.4 | 59.0 45.0 |
| Protein Fat TDF | 298.5 244.7 413.5 | 201.4 88.3 641.1 | 186.7 144.5 123.2 | 219.0 159.0 115.0 | 149.5 85.2 663.0 | 173.2 86.5 305.4 | 217.0 93.0 |
| References | [31] | [32] | [33] | [34] | [35] | [36] | [37] |
| TPC (mg GAE/g) | 199.4 | 179.0 | 94.5 | 122.9 | 213.4 |
| TFC (mg QE/g) | 102.1 | 68.77 | 378.7 | 41.5 | 30.6 |
| Lycopene content (g/ kg) | 36.7 | 41.4 | 50.1 | 50.2 | |
| DPPH radical scavenging activity (%) | 52.5 | 52.4 | 29.9 | 75.0 | |
| β-carotene bleaching inhibition activity (%) | 80.6 | 149.8 | 95.6 | 211.0 | |
| References | [32] | [35] | [38] | [39] | [40] |
| Calcium (Ca) | 160.0 | 76.4 | 371.5 | 141.6 | 131.8 | 140.5 |
| Phosphorus (P) | 219.7 | |||||
| Magnesium (Mg) | 149.0 | 126.7 | 3.1 | 251.1 | 211.0 | 157.3 |
| Sodium (Na) | 73.6 | 129.1 | 191.7 | 47.2 | 66.5 | 78.3 |
| Potassium (K) | 1097 | 1011.5 | 808.5 | 667.9 | 303.0 | 1125.0 |
| Iron (Fe) | 1.5 | 9.3 | 11.0 | 5.6 | 1.7 | |
| Zinc (Zn) | 3.12 | 3.46 | 1.8 | 6.3 | 0.5 | |
| References | [41] | [35] | [32] | [42] | [39] | [43] |
| Source | Amarelo | Caracal, Romania | Waltinger | Red Currant | Batateiro | Comprido |
|---|---|---|---|---|---|---|
| C12:0 | 1.5 | 1.1 | 0.4 | |||
| C14:0 | 9.3 | 4.1 | 0.9 | 0.6 | 6.2 | 3.2 |
| C15:0 | 1.5 | 0.9 | 0.1 | 0.2 | 1.1 | 0.8 |
| C16:0 | 205.3 | 163.2 | 133.9 | 133.9 | 193.1 | 159.6 |
| C17:0 | 3.3 | 1.9 | 0.9 | 2.4 | 2.5 | 1.8 |
| C18:0 | 63.4 | 54.3 | 43.5 | 46.7 | 54.7 | 63.6 |
| C20:0 | 12.6 | 4.8 | 5.8 | 8 | 6.1 | |
| C22:0 | 8.2 | 1.5 | 1.8 | 5.5 | 3.1 | |
| C23:0 | 15.2 | 0.2 | 0.2 | 7.8 | 1.6 | |
| C24:0 | 10.1 | 2.9 | 1.7 | 2 | 7.3 | 4.5 |
| C25:0 | 0.3 | 0.3 | ||||
| C26:0 | 0.8 | 0.9 | ||||
| SFA | 322.2 | 227.2 | 190 | 197.7 | 289.1 | 245.7 |
| C16:1 | 2.5 | 6.4 | 2.3 | 1.5 | ||
| C18:1n9 | 106 | 185 | 181.3 | 198.6 | ||
| C20:1 | 1.2 | 0.8 | 1 | |||
| C22:1n9 | 0.3 | 0.2 | 0.4 | |||
| MUFA | 110 | 197.5 | 192.4 | 207.9 | 130.8 | 176.6 |
| C18:2n6 | 398 | 519.1 | 463.3 | 520.5 | ||
| C20:4n6 | 0.6 | 0.4 | 0.1 | |||
| C22:2n6 | 3.9 | 0.1 | 0.4 | |||
| C22:3n6 | 5.5 | |||||
| n-6 PUFA | 398.6 | 530.7 | 463.7 | 520.6 | ||
| C18:3n3 | 155.3 | 35.5 | 114.1 | 55.5 | ||
| C20:3n3 | 2 | 0.9 | 0.8 | |||
| C20:5n3 | 0.3 | 2.6 | 0.5 | 0.4 | ||
| C22:3n3 | 1.3 | |||||
| n-3 PUFA | 157.6 | 42.2 | 115.5 | 56.7 | ||
| MUFA/SFA, % | 34.1 | 86.9 | 101.1 | 105.2 | 45.2 | 71.9 |
| n-3 PUFA/n-6 PUFA, % | 39.5 | 8 | 24.9 | 10.9 | ||
| References | [40] | [39] | [44] | [44] |
| Source | Caracal, Romania | Turkey (Seed) | No Message | Cairo, Egypt | Sicily, Italy | Best Factory (Peel) | Pig Requirements | Poultry Requirements |
|---|---|---|---|---|---|---|---|---|
| Indispensable amino acids | ||||||||
| Arginine | 14.6 | 10.6 | 1.8 | 10.4 | 10.9 | 43.4 | 2.4 | 10 |
| Histidine | 2.6 | 0.5 | 4.6 | 5.1 | 36.4 | 2.1 | 2.7 | |
| Isoleucine | 6.9 | 2.9 | 0.8 | 9.6 | 6.3 | 38.6 | 3.7 | 6.2 |
| Leucine | 10.7 | 6.4 | 1.5 | 14.6 | 11.9 | 50.7 | 6.7 | 9.3 |
| Lysine | 8.8 | 5.9 | 1.7 | 10.4 | 7.9 | 44 | 6.6 | 8.5 |
| Methionine | 2.7 | 3.1 | 1.2 | 4 | 10.2 | 1.8 | 3.2 | |
| Phenylalanine | 6.1 | 9 | 9.8 | 7.2 | 50.2 | 4 | 5.6 | |
| Threonine | 5.5 | 4.3 | 8.1 | 6 | 23.4 | 4.3 | 6.8 | |
| Tryptophan | 6 | 34.2 | 1.2 | 1.6 | ||||
| Valine | 5.4 | 3.6 | 1.2 | 12.3 | 7 | 45.8 | 4.5 | 7 |
| Dispensable amino acids | ||||||||
| Alanine | 7.1 | 4.7 | 1 | 10.7 | 7.9 | 50.2 | ||
| Aspartic acid | 15.7 | 10.3 | 2.4 | 32.9 | 13.2 | 7 | ||
| Cysteine | 2.3 | 3.1 | 0.5 | 2.7 | 4.1 | 3.9 | ||
| Glycine | 6.3 | 12.7 | 6.7 | 42.9 | ||||
| Glutamic acid | 72.1 | 4.8 | 5.4 | 61.2 | 29.7 | 145.6 | ||
| Proline | 4.3 | 0.9 | 11.1 | 27.8 | ||||
| Serine | 1.7 | 4.5 | 1 | 3.7 | 7.5 | 30.8 | ||
| Tyrosine | 6.9 | 2.5 | 7.1 | 2.3 | 34.2 | |||
| Total amino acids | 172.4 | 80.1 | 131 | 156 | 719.3 | |||
| References | [39] | [45] | [46] | [47] | [48] | [41] | [49] | [50] |
| References | Species/ Breed | Age | Type | Level, % | Performance |
|---|---|---|---|---|---|
| [19] | Male IR and Cobb | 1–42 days | DTP | 4, 6 | Increased feed cost, total variable cost, and total cost by 4% and 6%, and feeding TP consumed more feed. |
| Lower pH. No negative effect of adding 6% on growth performance parameters, WHC or drip loss, mRNA expression of GHR or IGF-1. | |||||
| [137] | Japanese Quail | 8 weeks | DTP | 3, 6, 9, 12 | Improves immune performance, antioxidant properties, and digestive enzymes. Lower cholesterol, LDL. |
| Increased HDL, egg weight, and hatchability, the largest of which was 6%, had a positive effect on lycopene deposition. | |||||
| [138] | Male Arian | 1–42 days | DTP | 3, 5 | Increased body weight and production index from 5%. Reduced feed conversion ratio in 5%. Reduced serum triglyceride and HDL cholesterol concentrations on Day 28 from 5%. Increases GPx and SOD activities and decreases MDA from 5%. No effect on growth performance. Improved serum enzyme activity, GPx, and lipid peroxidation during heat stress. |
| [142] | Ross 308 | 21–42 days | DTP | 5, 10, 15, 20 | Decreased body weight in 15% and 20%. Increased feed intake. Decreased nutrient apparent digestibility and crude fat apparent metabolizable energy and apparent digestibility. |
| [144] | Cobb-500 | 4–6 weeks | DTP | 10, 15, 20 | Increased weight gain and ADG. No effect on FCR. Lower heterophil/lymphocyte (H/L) ratio. Increase in catalase level from 20%. |
| [145] | Wild duck | 1–72 days | DTP | 10, 15, 20 | Increased live weight and feed intake and the most economical from 20%. Decreased total cholesterol, triglycerides, and HDL from 20%. No effect on LDL and total protein. |
| References | Breed | Age | Type | Level, % | Performance |
|---|---|---|---|---|---|
| [48] | Nero Siciliano | 7 months | DTP | 15 | No effect on growth performance, flesh color, and muscle antioxidant capacity. Decreased intramuscular fat, SFA, and MUFA content. Increased PUFA, PUFA n-3 and PUFA n-6 concentrations, and the n-6:n-3 ratio. |
| [148] | Landrace × Yorkshire × Duroc | 18 weeks | Lycopene (Ly) Ketchup (Kc) | Ly 20 Kc 3.4 Ly 10 + Kc 1.7 | No effect on production traits, plasma lipids, including total lipids, total cholesterol, high-density and low-density cholesterol, and triglycerides. |
| Decreased MDA concentration in fresh pork belly. | |||||
| [149] | Barrows | BW: 50.3 ± 1.1 kg | DTP | 50 or 100 g/kg | No effect on growth performance and digestibility of nutrients. Increased GSH-Px and glucose, total protein, and globulin. |
| [150] | Piétrain × Landrace × Large White | BW: 75.04 ± 1.6 kg | Lycopene | 12.5, 25.0, 37.5 or 50.0 mg/kg | Decreased SOD in the liver, total cholesterol, LDL, HDL, and LDL:HDL. Decreased catalase gene expression, plasma urea, and triglyceride concentrations. |
| [151] | Duroc × Landrace × Yorkshire | 21–49 days | Lycopene | 50 mg/kg | Increased serum CAT activity, TC concentration, and jejunal SOD activity. Decreased serum and jejunal H2O2 concentrations. Increased mRNA and protein expression of NRF2 and CD36 and decreased KEAP1 expression in the jejunum. Increased villus height, villus/crypt ratio, and abundance of beneficial flora; decreased abundance of pathogenic bacteria. |
| [152] | Sow | Culture medium | Lycopene | 10 IU/mL | Delayed disruption of communication between oocytes and cumulus cells. Increased glutathione levels and fertilization rates in mature oocytes. |
| [153] | Duroc × Landrace × Yorkshire | BW: 63.89 ± 1.15 kg | Lycopene | 100 or 200 mg/kg | Increased muscle redness a* value, intramuscular fat, crude protein content, and antioxidant capacity. MyHC protein levels and percentage of slow-twitch fibers at 200 mg/kg. Decreased muscle lightness L* and yellow b* values, fast myosin levels, and percentage of fast-twitch fibers at 200 mg/kg. |
| References | Breed | Age | Type | Level, % | Performance |
|---|---|---|---|---|---|
| Goat | |||||
| [33] | Comisana | 45 days | DTP | Feel free to provide | Increased L*, b*, C*, and H*. No effect on growth performance and lipid oxidation. |
| Decreased TBARS. | |||||
| [155] | Markhoz | BW: 18.6 ± 0.7 kg | DTP | 10, 20, 30 | No effect on body weight, hot carcass, slaughter rate, carcass length, blood sugar, total protein, urea, or cholesterol. Increased crude fat and crude protein content in muscle at 30%. |
| [156] | Ossimi | BW: 19.25 ± 0.18 kg | DTP | 5, 10, 15 | No effect on the digestion of DM and CP and total blood lipids. Increased digestibility of OM, CF, EE, and NFE at 10% or 15%. Increased FW, TBWG, ADG, and TVFA. Decreased rumen pH and ammonia nitrogen concentration. |
| [157] | Awassi | 3–6 years | DTP | 30 | Decreased milk production and milk protein content. Increased milk fat content, n-6:n-3 ratio. No effect on conjugated linoleic acid ratio. |
| [18] | Saanen | BW: 46.2 ± 7.50 kg | DTP | 20, 40, 60 | Reduced weight at 60%. Increased milk production and fat mass at 20% and 40%. No effect on feed efficiency and feed conversion ratio, blood glucose, cholesterol, urea, albumin, T3, and T4. |
| [158] | Murciano-Granadina | BW: 39.4 ± 5.39 kg | DTP | 35 | No effect on nutrient apparent digestibility, the urinary excretion of total purine derivatives, milk production and composition, or total bacterial and methanogen abundance. |
| Decreased N in urine, microbial N flux in rumen, NH3-N and CH4. | |||||
| Cattle | |||||
| [159] | Holstein cow | -- | ETP | 10 | No effect on milk yield and composition. Increased vitamin concentration in milk, DM intake, and digestibility. Increased concentrations of total cholesterol, high-density lipoprotein cholesterol, serum aspartate aminotransferase, antioxidants, IgA, IgG, and IgM. |
| [160] | Xinjiang brown cow | -- | FTP | 14 | Increased DMI and 4% fat-corrected milk. No effect on average milk yield, feed conversion ratio, milk fat, protein, or total solids. Reduced feed costs and increased benefits. |
| [161] | Holstein cow | BW: 594.2 ± 37.8 kg | ETP | 7.5, 15 | No effect on dry matter and nutrient intake. No effect on digestibility of dry matter, organic matter, NDF, or ADF. No effect on fecal and rumen pH, or rumen ammonia. No effect on daily milk production, or the percentages of milk protein and fatty acids. Reduced total blood protein. |
| References | Breed | Age | Type | Level, % | Performance |
|---|---|---|---|---|---|
| [166] | Hycole × Grimaud | 38 days | TP | 3, 6 | No effect on muscle pH, carcass characteristics, muscle nutrient composition, and antioxidant status of meat. |
| Increased muscle polyunsaturated fatty acids, and the yellow (b*) and chromatic values at 6%. | |||||
| [168] | V-Line Male Rabbit | 5 weeks | TPE | 200 g | Increased SOD activity, economic benefits, growth performance, antioxidant status, regulation of AA levels in plasma and meat, and carcass weight. Reduced plasma total cholesterol and LDL. Reduced fat of the kidney, belly, and back. |
| [169] | NZW | 6 weeks | TPE | 100, 200, 250 mg/kg | Heaviest body weight, lowest feed intake, and best feed conversion ratio at 250 mg/kg. Reduced mortality. Increased catalase and glutathione peroxidase. Decreased plasma total protein, globulin, catalase, and glutathione peroxidase. Improved net income and economic benefits. |
| [170] | Bai Rabbit | 33 days | DTP | 30, 40, 60 | Increased liver weight at 60% and waist weight at 30%. Increased economic benefits. Decreased perirenal fat mass. |
| [171] | NZW | 45 days | DTP | 1, 2 | Increased final body weight and feed efficiency values. No effect on PCV, Hb, MCV, MCH, MCHC, lymphocytes, monocytes, neutrophils, or eosinophils. Increased phagocytic activity of leukocytes, IgG, IgM, and IgA. Increased serum and liver TAC, SOD, GST, and CAT. |
| [172] | Mature rabbit | 6–8 months | DTP | 14, 22, 30 | No effect on average daily feed intake, litter size, and mortality rate. Increased weaning weight. No effect on semen color and consistency, pH, sperm motility and viability, total protein, albumin, and globulin in semen. Increased ejaculation volume (at 30%) and sperm cell concentration. |
| [175] | NWZ male | 5 months | Lycopene | 500 mg/kg | No effect on FBW and water intake. Increased hemoglobin concentration, hematocrit value, red blood cell, platelet counts, serum total protein, albumin, globulin, glucose, and HDL concentrations. Decreased MAD, white blood cell count, serum urea concentration, creatinine concentration, total lipids, triglycerides, total cholesterol, and LDL concentrations. Increased total antioxidant and testosterone concentrations. Improved sperm quantity, quality, total sperm output, initial semen fructose concentration, and conception rate. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, S.; Chen, S.; Li, H.; Paengkoum, S.; Taethaisong, N.; Meethip, W.; Surakhunthod, J.; Sinpru, B.; Sroichak, T.; Archa, P.; et al. Sustainable Valorization of Tomato Pomace (Lycopersicon esculentum) in Animal Nutrition: A Review. Animals 2022, 12, 3294. https://doi.org/10.3390/ani12233294
Lu S, Chen S, Li H, Paengkoum S, Taethaisong N, Meethip W, Surakhunthod J, Sinpru B, Sroichak T, Archa P, et al. Sustainable Valorization of Tomato Pomace (Lycopersicon esculentum) in Animal Nutrition: A Review. Animals. 2022; 12(23):3294. https://doi.org/10.3390/ani12233294
Chicago/Turabian StyleLu, Shengyong, Shengchang Chen, Haixia Li, Siwaporn Paengkoum, Nittaya Taethaisong, Weerada Meethip, Jariya Surakhunthod, Boontum Sinpru, Thakun Sroichak, Pawinee Archa, and et al. 2022. "Sustainable Valorization of Tomato Pomace (Lycopersicon esculentum) in Animal Nutrition: A Review" Animals 12, no. 23: 3294. https://doi.org/10.3390/ani12233294
APA StyleLu, S., Chen, S., Li, H., Paengkoum, S., Taethaisong, N., Meethip, W., Surakhunthod, J., Sinpru, B., Sroichak, T., Archa, P., Thongpea, S., & Paengkoum, P. (2022). Sustainable Valorization of Tomato Pomace (Lycopersicon esculentum) in Animal Nutrition: A Review. Animals, 12(23), 3294. https://doi.org/10.3390/ani12233294