
Sensor-Based Detection of Predator Influence on Livestock: A Case Study Exploring the Impacts of Wild Dogs (Canis familiaris) on Rangeland Sheep



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
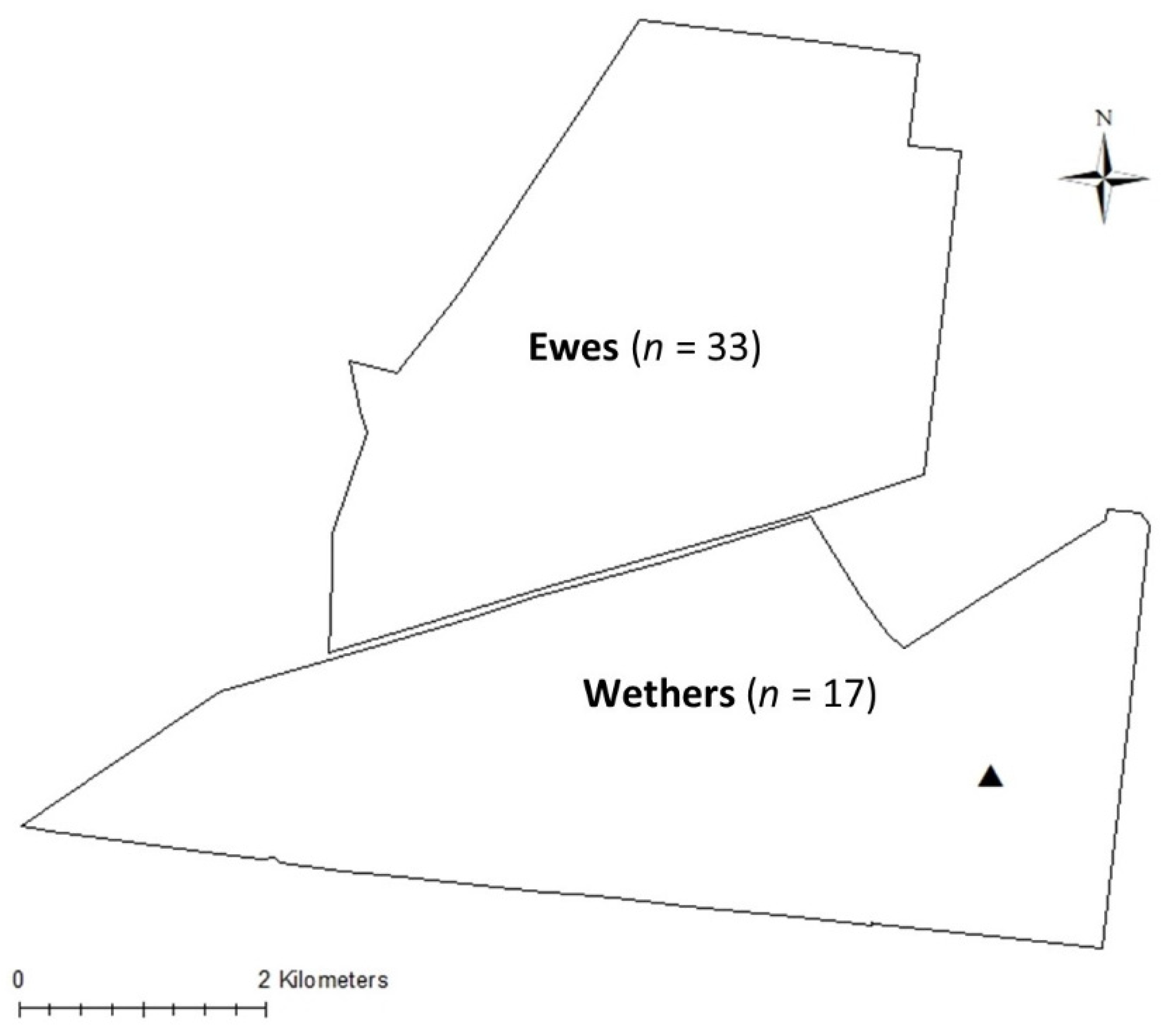
2.1. Location and Animals
2.2. Weather
2.3. GPS Tracking Collars and Data Analysis
2.4. Statistical Analysis
3. Results
3.1. Daily Distance Travelled and Activity
3.2. Diurnal Activity
3.3. Drivers of Daily Distance Travelled
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holder, F.W.; Howe, J.H.; Kirkpatrick, A.A.; Playford, T.; Moule, J.; Osman, J.J.; White, J.W. Report of the Vermin-Proof Fencing Commission, Together with the Proceedings, Evidence and Appendices; Government Printer: Adelaide, Australia, 1893.
- Allen, B.L.; West, P. Re: Dingoes are a major causal factor for the decline and distribution of sheep in Australia. Aust. Vet. J. 2015, 93, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Allen, B.L.; West, P. Influence of dingoes on sheep distribution in Australia. Aust. Vet. J. 2013, 91, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Fleming, P.J.S.; Allen, L.R.; Lapidge, S.J.; Robley, A.; Saunders, G.R.; Thomson, P.C. A strategic approach to mitigating the impacts of wild canids: Proposed activities of the Invasive Animals Cooperative Research Centre. Aust. J. Exp. Agric. 2006, 46, 753–762. [Google Scholar] [CrossRef]
- Jackson, S.M.; Fleming, P.J.; Eldridge, M.D.; Archer, M.; Ingleby, S.; Johnson, R.N.; Helgen, K.M. Taxonomy of the Dingo: It’s an ancient dog. Aust. J. Zool. 2021, 41, 347–357. [Google Scholar] [CrossRef]
- Primary Industries and Regions SA. The Dog Fence in South Australia. Available online: https://pir.sa.gov.au/biosecurity/introduced-pest-feral-animals/find_a_pest_animal/wild_dogs_and_dingoes/dog_fence (accessed on 10 August 2021).
- Gong, W.; Sinden, J.; Braysher, M.; Jones, R. The Economic Impacts of Vertebrate Pests in Australia; Invasive Animals CRC: Canberra, Australia, 2009.
- McLeod, R. Cost of Pest Animals in NSW and Australia, 2013–2014. In Report Prepared for the NSW Natural Resources Commission; eSYS Development Pty Ltd.: Sydney, Australia, 2016. [Google Scholar]
- Clark, P.; Clark, E.; Allen, B.L. Sheep, dingoes and kangaroos: New challenges and a change of direction 20 years on. In Advances in Conservation through Sustainable Use of Wildlife; Baxter, G., Finch, N., Murray, P., Eds.; University of Queensland: Brisbane, Australia, 2018; pp. 173–178. [Google Scholar]
- Petherick, J.C.; Edge, M. Measuring the welfare of livestock in extensive production systems: Can we, should we? In Proceedings of the 6th International Animal Welfare Conference, Brisbane, Australia, 31 August 2008; pp. 1–9. [Google Scholar]
- Temple, D.; Manteca, X. Animal welfare in extensive production systems is still an area of concern. Front. Sustain. Food Syst. 2020, 4, 154. [Google Scholar] [CrossRef]
- Fitzgerald, G.; Wilkinson, R. Assessing the Social Impact of Invasive Animals in Australia: A Report Produced for the Invasive Animals Coopoerative Research Centre; Invasive Animals Cooperative Research Centre: Canberra, Australia, 2009.
- Ecker, S.; Please, P.M.; Maybery, D.J. Constantly chasing dogs: Assessing landholder stress from wild dog attacks on livestock using quantitative and qualitative methods. Australas. J. Environ. Manag. 2017, 24, 16–30. [Google Scholar] [CrossRef]
- Allen, L. Best Practice Baiting: Dispersal and Seasonal Movement of Wild Dogs (Canis lupus familiaris); The State of Queensland, Department of Employment, Economic Development and Innovation: Brisbane, Australia, 2009; pp. 61–62.
- Van Eeden, L.M.; Crowther, M.S.; Dickman, C.R.; Macdonald, D.W.; Ripple, W.J.; Ritchie, E.G.; Newsome, T.M. Managing conflict between large carnivores and livestock. Conserv. Biol. 2018, 32, 26–34. [Google Scholar] [CrossRef]
- Allen, L.; Sparkes, E. The effect of dingo control on sheep and beef cattle in Queensland. J. Appl. Ecol. 2001, 38, 76–87. [Google Scholar] [CrossRef]
- Smith, B.P.; Appleby, R.G. Promoting human-dingo co-existence in Australia: Moving towards more innovative methods of protecting livestock rather than killing dingoes (Canis dingo). Wildl. Res. 2018, 45, 1–15. [Google Scholar] [CrossRef]
- Morris, J.E.; Cronin, G.M.; Bush, R.D. Improving sheep production and welfare in extensive systems through precision sheep management. Anim. Prod. Sci. 2012, 52, 665–670. [Google Scholar] [CrossRef]
- Fogarty, E.S.; Swain, D.L.; Cronin, G.; Trotter, M. Autonomous on-animal sensors in sheep research: A systematic review. Comput. Electron. Agric. 2018, 150, 245–256. [Google Scholar] [CrossRef]
- FrontierSI. SBAS Test-Bed Demonstrator Trial: Economic Benefits Report; 30/11/2021; FrontierSI: Docklands, Australia, 2019. [Google Scholar]
- Trotter, M.; Cosby, A.; Manning, J.; Thomson, M.; Trotter, T.; Graz, P.; Fogarty, E.; Lobb, A.; Smart, A. Demonstrating the Value of Animal Location and Behaviour in the Red Meat Value Chain; Meat and Livestock Australia Limited: North Sydney, Australia, 2018.
- Manning, J.K.; Fogarty, E.S.; Trotter, M.G.; Schneider, D.A.; Thomson, P.C.; Bush, R.D.; Cronin, G.M. A pilot study into the use of global navigation satellite system technology to quantify the behavioural responses of sheep during simulated dog predation events. Anim. Prod. Sci. 2014, 54, 1676–1681. [Google Scholar] [CrossRef]
- Queensland Government. SILO, Australian Climate Data from 1889 to Yesterday. Available online: https://www.longpaddock.qld.gov.au/silo (accessed on 23 July 2021).
- Marai, I.F.M.; Ayyat, M.S.; Abd El-Monem, U.M. Growth performance and reproductive traits at first parity of New Zealand White female rabbits as affected by heat stress and its alleviation, under Egyptian conditions. Trop. Anim. 2011, 33, 451–462. [Google Scholar] [CrossRef]
- Fogarty, E.S.; Swain, D.L.; Cronin, G.M.; Moraes, L.E.; Bailey, D.W.; Trotter, M.G. Potential for autonomous detection of lambing using global navigation satellite system technology. Anim. Prod. Sci. 2020, 60, 1217–1226. [Google Scholar] [CrossRef]
- Putfarken, D.; Dengler, J.; Lehmann, S.; Hardtle, W. Site use of grazing cattle and sheep in a large-scale pasture landscape: A GPS/GIS assessment. Appl. Anim. Behav. Sci. 2008, 111, 54–67. [Google Scholar] [CrossRef] [Green Version]
- Squires, V.R.; Wilson, A.D.; Daws, G.T. Comparisons of the walking activity of some Australian sheep. Proc. Aust. Soc. Anim. Prod. 1972, 9, 376–380. [Google Scholar]
- Heglund, N.C.; Taylor, R.C. Speed, stride frequency and energy cost per stride: How do they change with body size and gait? J. Exp. Biol. 1988, 138, 301–318. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Frid, A.; Dill, L. Human-caused Disturbance Stimuli as a Form of Predation Risk. Conserv. Ecol. 2001, 6, 11–26. [Google Scholar] [CrossRef]
- Dwyer, C.M. How has the risk of predation shaped the behavioural responses of sheep to fear and distress? Anim. Welf. 2004, 13, 269–281. [Google Scholar]
- Vandenheede, M.; Bouissou, M.F. Sex differences in fear reactions in sheep. Appl. Anim. Behav. Sci. 1993, 37, 39–55. [Google Scholar] [CrossRef]
- Hansen, I.I.; Christiansen, F.; Hansen, H.S.; Braastad, B.; Bakken, M. Variation in behavioural responses of ewes towards predator-related stimuli. Appl. Anim. Behav. Sci. 2001, 70, 227–237. [Google Scholar] [CrossRef]
- Pulliam, H.R. On the advantages of flocking. J. Theor. Biol. 1973, 38, 419–422. [Google Scholar] [CrossRef]
- Rook, A.; Penning, P. Synchronisation of eating, ruminating and idling activity by grazing sheep. Appl. Anim. Behav. Sci. 1991, 32, 157–166. [Google Scholar] [CrossRef]
- Hutson, G.D. Behavioural Principles of Sheep Handling. In Livestock Handling and Transport, 3rd ed.; Grandin, T., Ed.; Centre for Agriculture and Biosciences International (CABI): Wallingford, UK, 2007; pp. 155–174. [Google Scholar]
- Thomas, D.T.; Wilmot, M.G.; Alchin, M.; Masters, D.G. Preliminary indications that Merino sheep graze different areas on cooler days in the Southern Rangelands of Western Australia. Aust. J. Exp. Agric. 2008, 48, 889–892. [Google Scholar] [CrossRef]
- Monty, D., Jr; Kelley, L.; Rice, W. Acclimatization of St.-Croix, Karakul and Rambouillet sheep to intense and dry summer heat. Small Rumin. Res. 1991, 4, 379–392. [Google Scholar] [CrossRef]
- Leite, P.G.; Marques, J.I.; Furtado, D.A.; Neto, J.P.L.; de Souza, B.B.; do Nascimento, J.W.B. Ethology, physiological, and ingestive responses of sheep subjected to different temperatures and salinity levels of water. Int. J. Biometeorol. 2019, 63, 1091–1098. [Google Scholar] [CrossRef]
- Squires, V.R. Walking, watering and grazing behaviour of merino sheep on two semi-arid rangelands in south-west New South Wales. Aust. Rangel. J. 1976, 1, 13–23. [Google Scholar] [CrossRef]
- Fardell, L.L.; Pavey, C.R.; Dickman, C.R. Fear and stressing in predator-prey ecology: Considering the twin stressors of predators and people on mammals. PeerJ 2020, 8, e9104. [Google Scholar] [CrossRef]
- Schmitz, O.J.; Krivan, V.; Ovadia, O. Trophic cascades: The primacy of train-mediated indirect interactions. Ecol. Lett. 2004, 7, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Sejian, V.; Maurya, V.P.; Naqvi, S.M. Effect of walking stress on growth, physiological adaptability and endocrine responses in Malpura ewes in a semi-arid tropical environment. Int. J. Biometeorol. 2012, 56, 243–252. [Google Scholar] [CrossRef]
- Epperly, H.K.; Clinchy, M.; Zanette, L.Y.; McCeery, R.A. Fear of large carnivores is tied to ungulate habitat use: Evidence from a bifactorial experiment. Sci. Rep. 2021, 11, 12979. [Google Scholar] [CrossRef]
- Tambling, C.J.; Minnie, L.; Meyer, J.; Freeman, E.W.; Santymire, R.M.; Adendorff, J.; Kerley, G.I.H. Temporal shifts in activity of prey following large predator reintroductions. Behav. Ecol. Sociobiol. 2015, 69, 1153–1161. [Google Scholar] [CrossRef]
- Illius, A.W.; Fitzgibbon, C. Costs of vigilance in foraging ungulates. Anim. Behav. 1994, 47, 481–484. [Google Scholar] [CrossRef] [Green Version]
- Fortin, D.; Boyce, M.S.; Merrill, E.H.; Fryxell, J.M. Foraging costs of vigilance in large mammalian herbivores. Oikos 2004, 107, 172–180. [Google Scholar] [CrossRef]
- Barwick, J.; Lamb, D.; Dobos, R.; Schneider, D.; Welch, M.; Trotter, M. Predicting Lameness in Sheep Activity Using Tri-Axial Acceleration Signals. Animals 2018, 8, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogarty, E.S.; Swain, D.L.; Cronin, G.M.; Moraes, L.E.; Trotter, M. Can accelerometer ear tags identify behavioural changes in sheep associated with parturition? Anim. Reprod. Sci. 2020, 216, 106345. [Google Scholar] [CrossRef] [PubMed]
- Dobos, R.C.; Dickson, S.; Bailey, D.W.; Trotter, M.G. The use of GNSS technology to identify lambing behaviour in pregnant grazing Merino ewes. Anim. Prod. Sci. 2014, 54, 1722–1727. [Google Scholar] [CrossRef]
- Fogarty, E.S.; Manning, J.K.; Trotter, M.G.; Schneider, D.A.; Thomson, P.C.; Bush, R.D.; Cronin, G.M. GNSS technology and its application for improved reproductive management in extensive sheep systems. Anim. Prod. Sci. 2015, 55, 1272–1280. [Google Scholar] [CrossRef]
- Falzon, G.; Schneider, D.; Trotter, M.; Lamb, D. A relationship between faecal egg counts and the distance travelled by sheep. Small Rumin. Res. 2013, 111, 171–174. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Variable | Dog Present | Dog Absent | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Min | Median | Mean | Max | S.D. | Min | Median | Mean | Max | S.D. | |
| Daily distance (km) | 4.2 | 11.1 | 11.6 | 21.0 | 3.12 | 1.9 | 8.9 | 9.2 | 17.6 | 2.73 |
| Percent Active (%/day) | 45.3 | 69.9 | 69.3 | 89.5 | 6.25 | 39.5 | 69.3 | 68.7 | 85.1 | 6.35 |
| Maximum THI | 18.4 | 23.4 | 23.4 | 27.3 | 2.23 | 21.2 | 27.7 | 26.7 | 30.6 | 2.57 |
| Minimum THI | 4.0 | 10.8 | 10.2 | 16.2 | 3.12 | 6.2 | 13.7 | 13.4 | 19.0 | 3.87 |
| Water Visits | 0 | 1 | 1.1 | 4 | 0.73 | 0 | 1 | 0.9 | 4 | 0.83 |
| Parameter | Estimate | S.E. | p-Value |
|---|---|---|---|
| Intercept | 1.359 | 0.068 | 0.041 |
| Slope for maximum THI | −0.026 | 0.001 | <0.001 |
| Slope for dog presence | 0.119 | 0.009 | <0.001 |
| Slope for paddock (wethers) | 0.196 | 0.034 | <0.001 |
| Slope for water visits | 0.076 | 0.005 | <0.001 |
| Slope for time spent active | 0.020 | 0.001 | <0.001 |
| Wethers | Ewes | |||||
|---|---|---|---|---|---|---|
| Parameter | Estimate | S.E. | p-Value | Estimate | S.E. | p-Value |
| Intercept | 1.583 | 0.124 | <0.001 | 1.375 | 0.083 | 0.013 |
| Slope for THI max | −0.022 | 0.002 | <0.001 | −0.029 | 0.002 | <0.001 |
| Slope for dog presence | 0.136 | 0.015 | <0.001 | 0.111 | 0.012 | <0.001 |
| Slope for water visits | 0.081 | 0.009 | <0.001 | 0.078 | 0.006 | <0.001 |
| Slope for active | 0.018 | 0.001 | <0.001 | 0.021 | 0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, C.A.; Trotter, M.G.; Manning, J.K. Sensor-Based Detection of Predator Influence on Livestock: A Case Study Exploring the Impacts of Wild Dogs (Canis familiaris) on Rangeland Sheep. Animals 2022, 12, 219. https://doi.org/10.3390/ani12030219
Evans CA, Trotter MG, Manning JK. Sensor-Based Detection of Predator Influence on Livestock: A Case Study Exploring the Impacts of Wild Dogs (Canis familiaris) on Rangeland Sheep. Animals. 2022; 12(3):219. https://doi.org/10.3390/ani12030219
Chicago/Turabian StyleEvans, Caitlin A., Mark G. Trotter, and Jaime K. Manning. 2022. "Sensor-Based Detection of Predator Influence on Livestock: A Case Study Exploring the Impacts of Wild Dogs (Canis familiaris) on Rangeland Sheep" Animals 12, no. 3: 219. https://doi.org/10.3390/ani12030219
APA StyleEvans, C. A., Trotter, M. G., & Manning, J. K. (2022). Sensor-Based Detection of Predator Influence on Livestock: A Case Study Exploring the Impacts of Wild Dogs (Canis familiaris) on Rangeland Sheep. Animals, 12(3), 219. https://doi.org/10.3390/ani12030219