
Sex Determination in Japanese Quails (Coturnix japonica) Using Geometric Morphometrics of the Skull




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sample
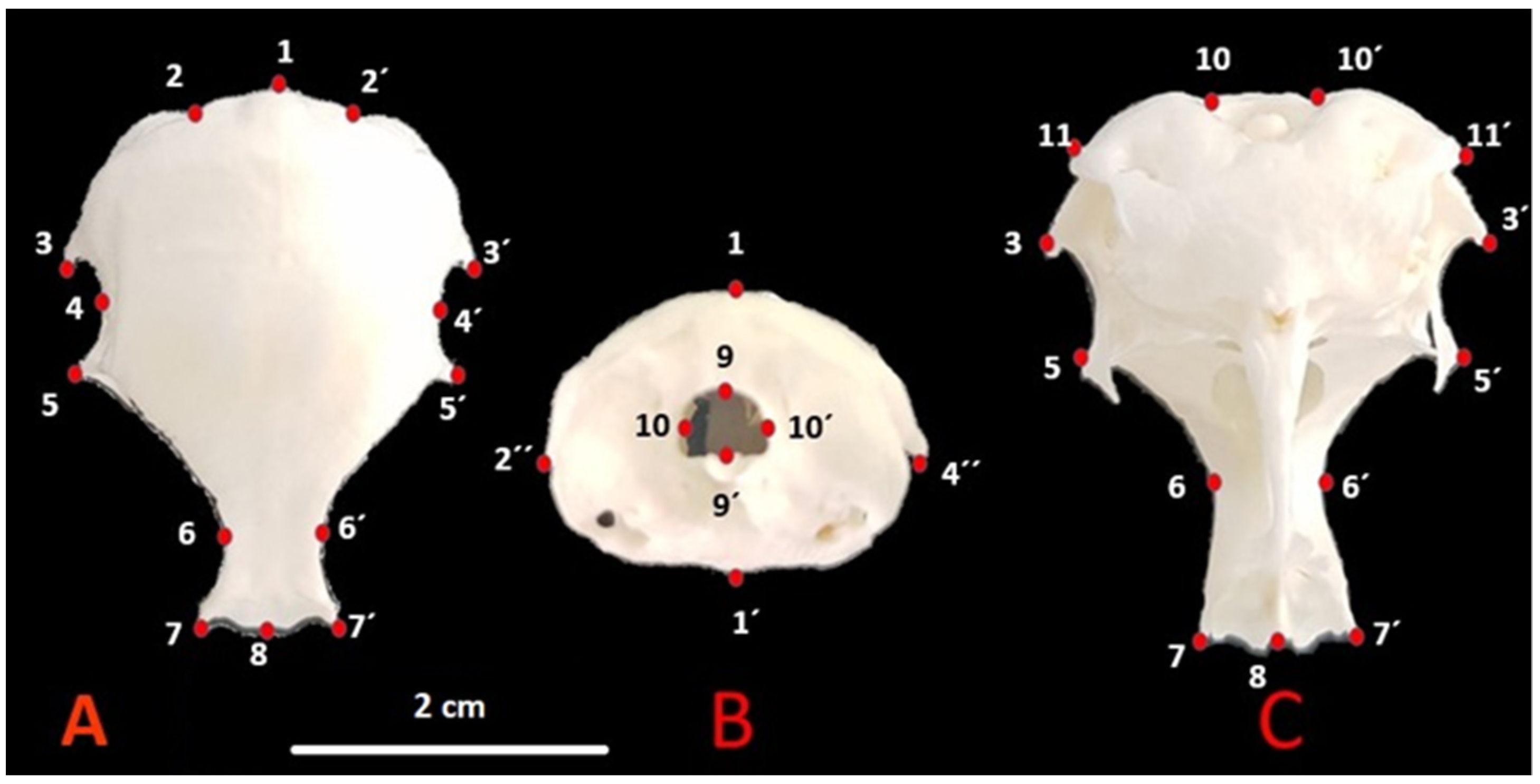
2.2. Landmarks and Data Collections
- (A).
- 1—Cerebellar prominentia; 2, 2′—Exoccipital bone; 3, 3′—Proc. suprameaticus of squamosal bone; 4, 4′—Temporal fossa; 5, 5′—Postorbital process; 6, 6′—Dorsal middle point of supraorbital margin; 7, 7′—Craniolateral terminal point of frontal bone; 8—Middle point of frontonasal suture;
- (B).
- 1—Cerebellar prominentia; 1′—Basilar tuberculum of basioccipital bone; 2″, 4″—Postorbital process; 9, 9′—Foramen magnum high; 10, 10′—Foramen magnum width;
- (C).
- 10, 10′—Foramen magnum width; 11, 11′—Paraoccipital process; 3, 3′—Procc. suprameaticus of squamosal bone; 5, 5′—Postorbital process; 6, 6′—Dorsal middle point of supraorbital margin; 7, 7′—Craniolateral terminal point of frontal bone; 8—Middle point of frontonasal suture.
2.3. Statistical Analysis
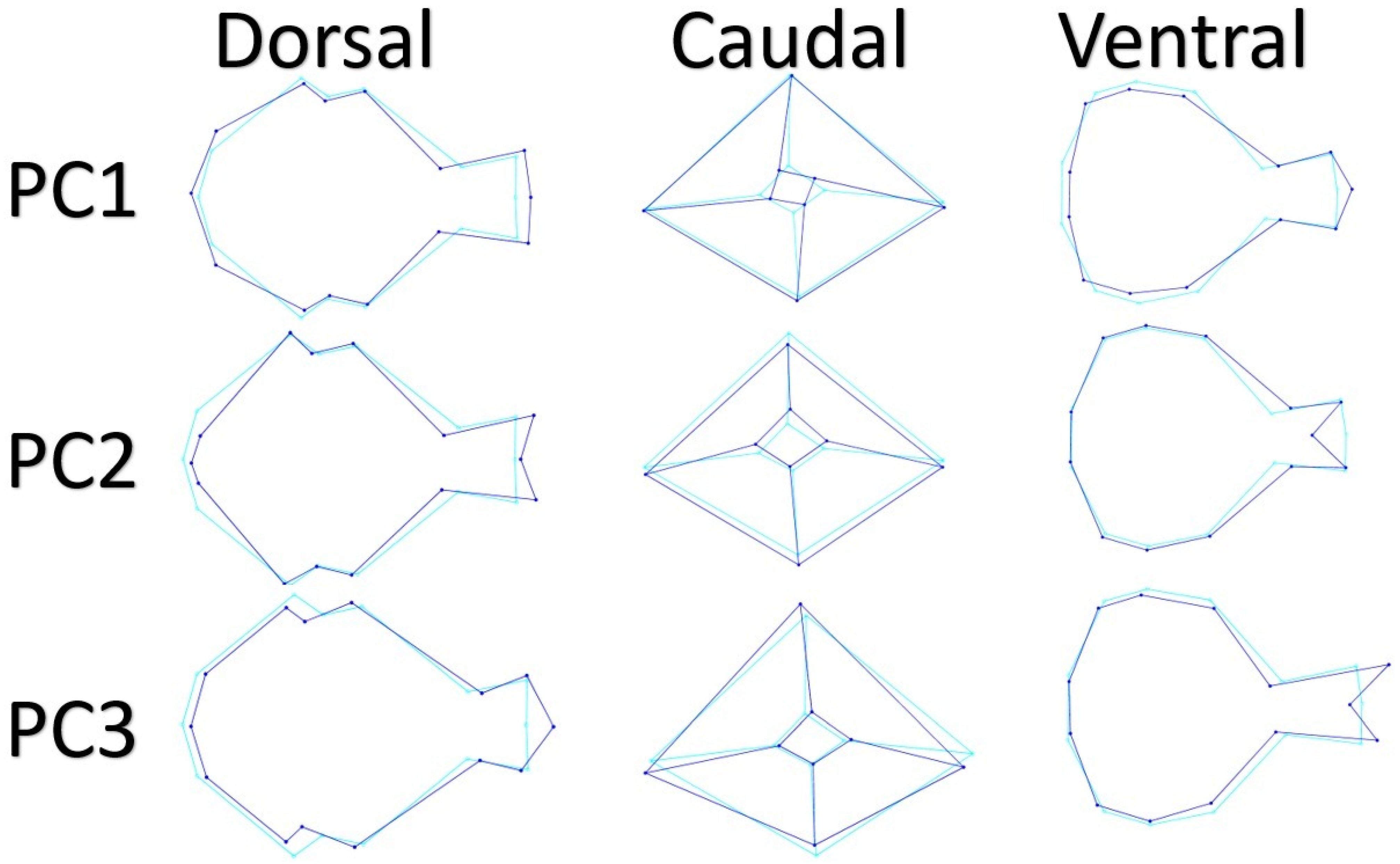
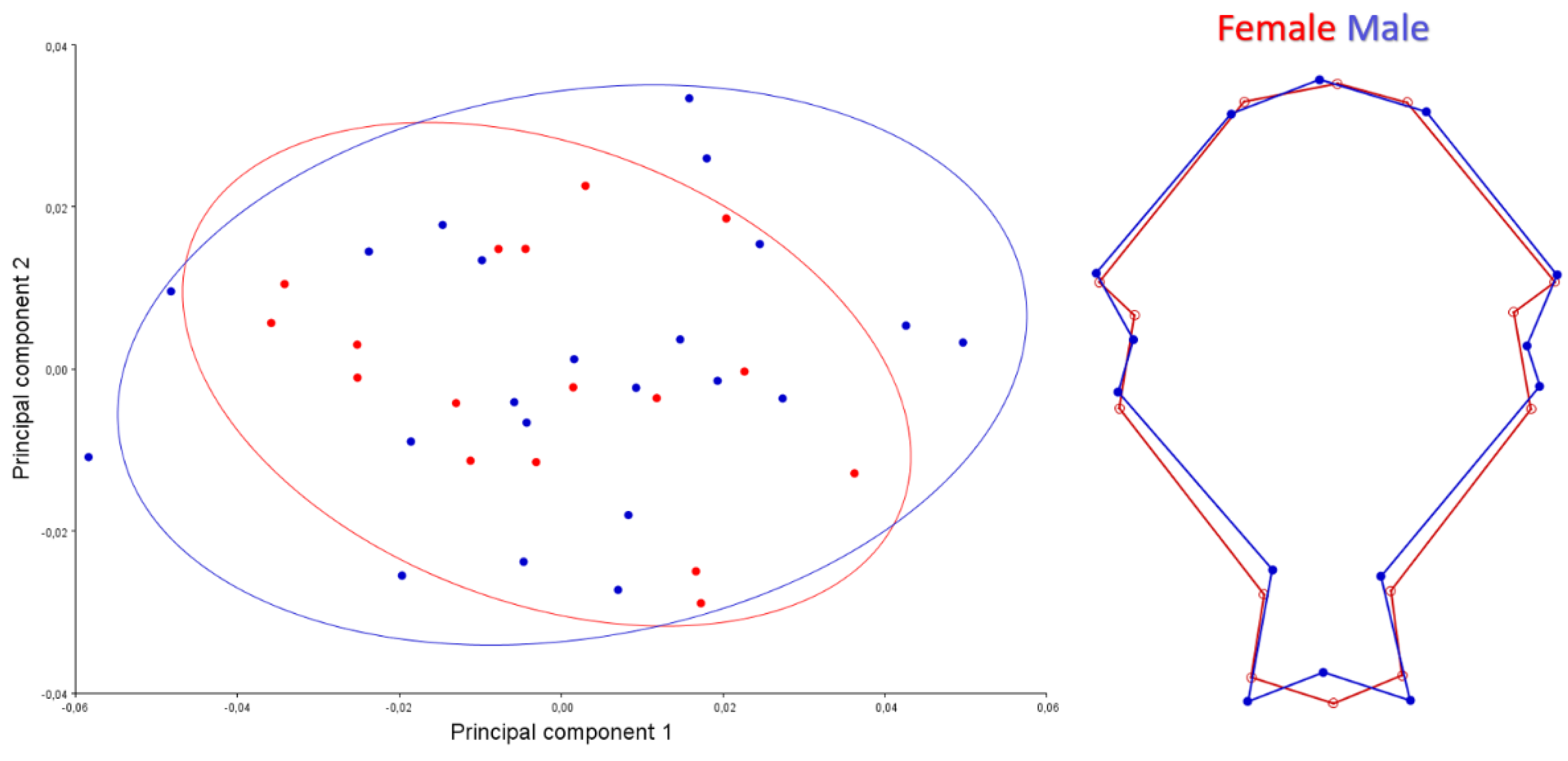
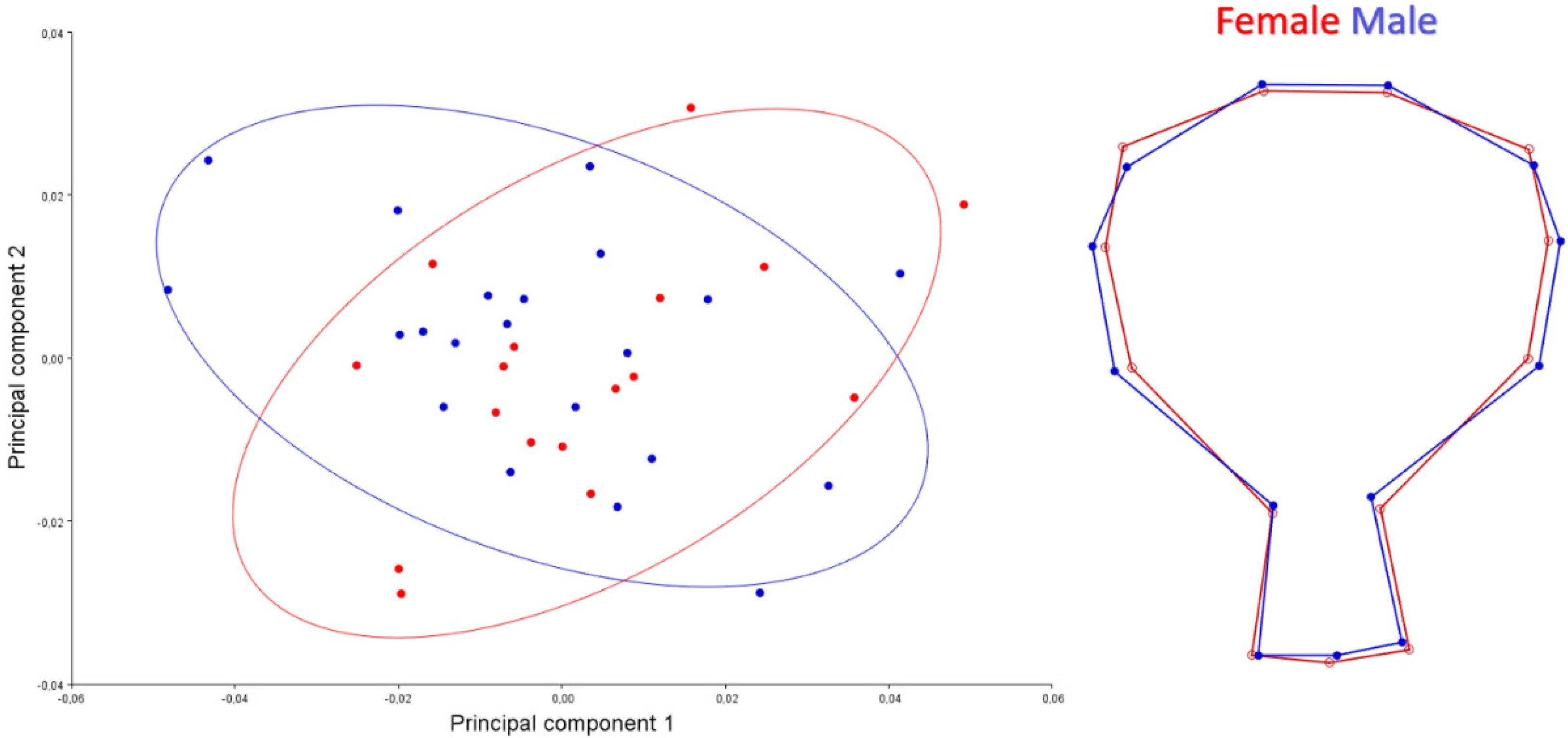
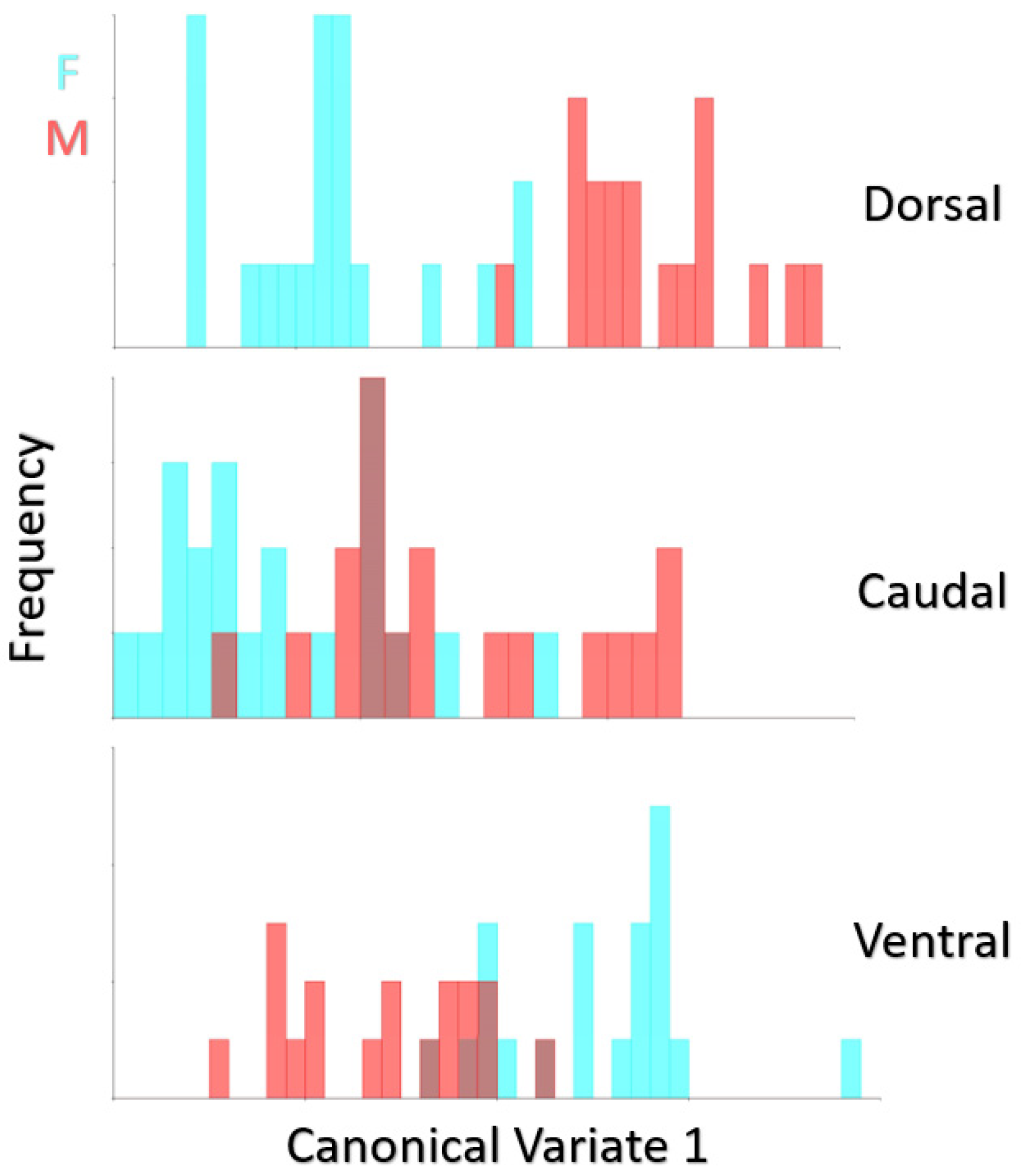
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vali, N. The Japanese quail: A review. Int. J. Poult. Sci. 2008, 7, 925–931. [Google Scholar] [CrossRef] [Green Version]
- Çağlayan, T.; Şeker, E. Dağ nanesinin (Mentha caucasica) japon bıldırcınlarının (Coturnix coturnix Japonica) performans, bazı vücut ölçüleri ve canlı ağırlık arasındak ilişkilerine etkisi. Eurasian J. Vet. Sci. 2015, 31, 33–42. [Google Scholar]
- Mizutani, M. The Japanese Quail. Available online: https://www.angrin.tlri.gov.tw/apec2003/Chapter5JPQuail.pdf/21/02/2017 (accessed on 20 December 2021).
- Minvielle, F. The future of Japanese quail for research and production. World’s Poult. Sci. J. 2004, 60, 500–507. [Google Scholar] [CrossRef]
- Peters, J.L. Check-List of Birds of the World; Harvard University Press: Cambridge, MA, USA, 1934; Volume 2, p. 92. [Google Scholar]
- Gill, F.; Donsker, D.; Rasmussen, P. IOC World Bird List (v11.2). 2021. Available online: https://www.gbif.org/dataset/c696e5ee-9088-4d11-bdae-ab88daffab78 (accessed on 20 December 2021).
- Barilani, M.; Derégnaucourt, S.; Gallego, S.; Galli, L.; Mucci, N.; Piombo, R.; Puigcerver, M.; Rimondi, S.; Rodriguez-Teijeiro, J.D.; Spano, S.; et al. Detecting hybridization in wild (Coturnix c. coturnix) and domesticated (Coturnix c. japonica) quail populations. Biol. Conserv. 2005, 126, 445–455. [Google Scholar] [CrossRef]
- Dacke, C.G.; Arkle, S.; Cook, D.J.; Wormstone, I.M.; Jones, S.; Zaidi, M.; Bascal, Z.A. Medullary bone and avian calcium regulation. J. Exp. Biol. 1993, 184, 63–88. [Google Scholar] [CrossRef]
- Koenig, H.E.; Korbel, R.; Liebich, H.G.; Klupiec, C. Avian Anatomy: Textbook and Colour Atlas; 5M Books Ltd.: Sheffield, UK, 2016. [Google Scholar]
- Baumel, J.J. Handbook of Avian Anatomy: Nomina Anatomica Avium; Publications of the Nuttall Ornithological Club: Cambridge, MA, USA, 1993. [Google Scholar]
- Koch, T. Anatomy of the Chicken and Domestic Birds; Iowa State University Press: Ames, IA, USA, 1973; pp. 48–51. [Google Scholar]
- Nickel, R.; Schummer, A.; Seiferle, E. Anatomy of the Domestic Birds; Verlag Paul Parey: Berlin/Hamburg, Germany, 1977. [Google Scholar]
- McLelland, J. A Colour Atlas of Avian Anatomy; Wolfe Medical Publications Ltd.: London, UK, 1990. [Google Scholar]
- Bärmann, E.V.; Wronski, T.; Lerp, H.; Azanza, B.; Börner, S.; Erpenbeck, D.; Rössner, G.E.; Wörheide, G. Morphometric and genetic framework for Gazella. Zool. J. Linn. Soc. 2013, 169, 673–696. [Google Scholar] [CrossRef] [Green Version]
- Kobryńczuk, F.; Krasińska, M.; Szara, T. Polarization of skull shapes in adult Lowland European bison, Bison bonasus bonasus. Ann. Zool. Fenn. 2008, 45, 341–346. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; Marugán-Lobón, J. Evolutionary covariation in geometric morphometric data: Analyzing integration, modularity, and allometry in a phylogenetic context. Syst. Biol. 2013, 62, 591–610. [Google Scholar] [CrossRef]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D.; Fink, W.L. Geometric Morphometrics for Biologists: A Primer, 2nd ed.; Elsevier Academic Press: San Diego, CA, USA, 2004. [Google Scholar] [CrossRef]
- Kimmerle, E.H.; Ross, A.; Slice, D. Sexual dimorphism in America: Geometric morphometric analysis of the craniofacial region. J. Forensic. Sci. 2008, 53, 54–57. [Google Scholar] [CrossRef]
- Mitteroecker, P.; Gunz, P. Advances in geometric morphometrics. Evol. Biol. 2009, 36, 235–247. [Google Scholar] [CrossRef] [Green Version]
- Bigoni, L.; Velemínská, J.; Brůžek, J. Three-dimensional geometric morphometric analysis of cranio-facial sexual dimorphism in a Central European sample of known sex. Homo 2010, 61, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Demircioglu, İ.; Demiraslan, Y.; Gurbuz, İ.; Dayan, M.O. Geometric Morphometric Analysis of Skull and Mandible in Awassi Ewe and Ram. Kafkas Üniversitesi Vet. Fakültesi Derg. 2021, 27, 43–49. [Google Scholar]
- Parés-Casanova, P.M.; Martínez, S. Geometric morphometrics for the study of hemicoxae sexual dimorphism in a local domestic equine breed. Int. J. Morphol. 2013, 31, 623–628. [Google Scholar] [CrossRef] [Green Version]
- Parés-Casanova, P.M.; Domènech-Domènech, X. A comparative analysis of sphenoid bone between domestic sheep (ovis aries) and goat (capra hircus) using geometric morphometrics. Anat. Histol. Embryol. 2021, 50, 556–561. [Google Scholar] [CrossRef] [PubMed]
- Siddiq, A.B.; Pares-Casanova, P.M.; Öncü, Ö.E.; Kar, H.; Onar, V. High level of fluctuating asymmetry in the Byzantine dogs from the Theodosius Harbor, Istanbul, Turkey. Turk. J. Vet. Anim. Sci. 2021, 45, 248–256. [Google Scholar]
- Gürbüz, İ.; Aytek, A.İ.; Demiraslan, Y.; Onar, V.; Özgel, Ö. Geometric morphometric analysis of cranium of wolf (Canis lupus) and German shepherd dog (Canis lupus familiaris). Kafkas Üniversitesi Vet. Fakültesi Derg. 2020, 26, 525–532. [Google Scholar]
- Rohlf, F.J. The tps series of software. Hystrix 2015, 26, 9–12. [Google Scholar] [CrossRef]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Gündemir, O.; Duro, S.; Jashari, T.; Kahvecioğlu, O.; Demircioğlu, İ.; Mehmeti, H. A study on morphology and morphometric parameters on skull of the Bardhoka autochthonous sheep breed in Kosovo. Anat. Histol. Embryol. 2020, 49, 365–371. [Google Scholar] [CrossRef]
- Süzer, B.; Serbest, A.; Arican, İ.; Yonkova, P.; Yilmaz, B. A morphometric study on the skull of the turkeys (Meleagris gallopavo). Uludağ Üniversitesi Vet. Fakültesi Derg. 2018, 37, 93–100. [Google Scholar] [CrossRef]
- Özkan, E.; Siddiq, A.B.; Kahvecioğlu, K.O.; Öztürk, M.; Onar, V. Morphometric analysis of the skulls of domestic cattle (Bos taurus L.) and water buffalo (Bubalus bubalis L.) in Turkey. Turk. J. Vet. Anim. Sci. 2019, 43, 532–539. [Google Scholar] [CrossRef]
- Jashari, T.; Duro, S.; Gündemir, O.; Szara, T.; Ilieski, V.; Mamuti, D.; Choudhary, O.P. Morphology, morphometry and some aspects of clinical anatomy in the skull and mandible of Sharri sheep. Biologia 2021, 1–11. [Google Scholar] [CrossRef]
- Pares-Casanova, P.M.; Siddiq, A.B.; Onar, V. Cranial size and shape sexual dimorphism in the Kangal dog from Turkey. Turk. J. Vet. Anim. Sci. 2020, 44, 396–403. [Google Scholar] [CrossRef]
- Demiraslan, Y.; Özgel, Ö.; Gürbüz, İ.; Kaştan, Ö. The mandibles of the Honamli and Hair goat (Capra hircus); a geometric morphometric study. Ank. Üniversitesi Vet. Fakültesi Derg. 2021, 68, 321–328. [Google Scholar] [CrossRef]
- Parés-Casanova, P.M. Geometric Morphometrics to the Study of Skull Sexual Dimorphism in a Local Domestic Goat Breed. J. Fish. Livest. Prod. 2015, 3, 1–4. [Google Scholar]
- Kraft, E. Vergleichend Morphologische Untersuchungen an Einzelknochen Nord-und Mitteleuropaischer Kleinerer Huhnervogel. Ph.D. Thesis, Ludwig-Maximilians-Universität, München, Germany, 1972. [Google Scholar]
- Moselhy, A.A.; Mohamed, S.K.A.; El-Ghazali, H.M. Anatomical features of bones and bony cavities of the ostrich skull (Struthio camelus). Int. J. Anat. Res. 2018, 6, 5390–5398. [Google Scholar]
- Gündemir, O.; Özkan, E.; Dayan, M.O.; Aydoğdu, S. Sexual analysis in turkey (Meleagris gallopavo) neurocranium using geometric morphometric methods. Turk. J. Vet. Anim. Sci. 2020, 44, 681–687. [Google Scholar] [CrossRef]
- Chovalopoulou, M.E.; Bertsatos, A. Estimating sex of modern Greeks based on the foramen magnum region. J. Anthropol. 2017, 2017, 9129801. [Google Scholar] [CrossRef] [Green Version]
- Lau, C.; Asahara, M.; Han, S.Y.; Kimura, J. Sexual dimorphism of the Eurasian otter (Lutra lutra) in South Korea: Craniodental geometric morphometry. J. Vet. Med. Sci. 2016, 78, 1007–1011. [Google Scholar] [CrossRef] [Green Version]
- Livezey, B.C.; Humphrey, P.S. Sexual dimorphism in continental steamer-ducks. Condor 1984, 86, 368–377. [Google Scholar] [CrossRef]
- Johnston, R.F. Variation in size and shape in pigeons, Columba livia. Wilson Bull. 1990, 102, 213–225. [Google Scholar]
- Mills, A.D.; Crawford, L.L.; Domjan, M.; Faure, J.M. The behavior of the Japanese or domestic quail Coturnix japonica. Neurosci. Biobehav. Rev. 1997, 21, 261–281. [Google Scholar] [CrossRef]
- Bagh, J.; Panigrahi, B.; Panda, N.; Pradhan, C.R.; Mallik, B.K.; Majhi, B.; Rout, S.S. Body weight, egg production, and egg quality traits of gray, brown, and white varieties of Japanese quail (Coturnix coturnix japonica) in coastal climatic condition of Odisha. Vet. World 2016, 9, 832–836. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PC | Dorsal | Caudal | Ventral | |||
|---|---|---|---|---|---|---|
| Eigenvalue | % V | Eigenvalue | % V | Eigenvalue | % V | |
| 1 | 0.000532077 | 33.046 | 0.00235293 | 41.206 | 0.000484881 | 34.495 |
| 2 | 0.000228691 | 14.203 | 0.00195548 | 34.246 | 0.000318786 | 22.679 |
| 3 | 0.000208817 | 12.969 | 0.000515875 | 9.344 | 0.000128329 | 9.1295 |
| 4 | 0.000125772 | 7.8113 | 0.000364257 | 6.3791 | 0.000106107 | 7.5486 |
| 5 | 0.000105952 | 6.5804 | 0.000159492 | 2.7931 | 7.22557 × 10−5 | 5.1404 |
| 6 | 8.52045E-05 | 5.2918 | 0.000106751 | 1.8695 | 5.58247 × 10−5 | 3.9714 |
| Individuals | Database | F | p-Value | |
|---|---|---|---|---|
| Sex | Dorsal | Centroid Size | 0.01 | 0.9136 |
| Shape | 1.83 | 0.0092 | ||
| Caudal | Centroid Size | 4.87 | 0.0336 | |
| Shape | 0.56 | 0.8717 | ||
| Ventral | Centroid Size | 2.07 | 0.1584 | |
| Shape | 0.79 | 0.7406 |
| Dorsal | Caudal | Ventral | ||
|---|---|---|---|---|
| Female | Mean | 14,975.44 | 8230.163 | 16,311.38 |
| SD | 6112.519 | 2656.018 | 6623.777 | |
| Male | Mean | 17,561.24 | 7113.946 | 15,894.75 |
| SD | 526.8477 | 3880 | 6784.003 |
| MD | MD-P | PD | PD-P | |
|---|---|---|---|---|
| Dorsal | 3.5694 | <0.0001 | 0.0178 | 0.0510 |
| Caudal | 1.3809 | 0.0751 | 0.0167 | 0.4923 |
| Ventral | 2.2510 | <0.0001 | 0.0100 | 0.5791 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szara, T.; Duro, S.; Gündemir, O.; Demircioğlu, İ. Sex Determination in Japanese Quails (Coturnix japonica) Using Geometric Morphometrics of the Skull. Animals 2022, 12, 302. https://doi.org/10.3390/ani12030302
Szara T, Duro S, Gündemir O, Demircioğlu İ. Sex Determination in Japanese Quails (Coturnix japonica) Using Geometric Morphometrics of the Skull. Animals. 2022; 12(3):302. https://doi.org/10.3390/ani12030302
Chicago/Turabian StyleSzara, Tomasz, Sokol Duro, Ozan Gündemir, and İsmail Demircioğlu. 2022. "Sex Determination in Japanese Quails (Coturnix japonica) Using Geometric Morphometrics of the Skull" Animals 12, no. 3: 302. https://doi.org/10.3390/ani12030302
APA StyleSzara, T., Duro, S., Gündemir, O., & Demircioğlu, İ. (2022). Sex Determination in Japanese Quails (Coturnix japonica) Using Geometric Morphometrics of the Skull. Animals, 12(3), 302. https://doi.org/10.3390/ani12030302