
Associations of Transcription Factor 21 Gene Polymorphisms with the Growth and Body Composition Traits in Broilers

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. ChIP-Seq Analysis
2.2. Experimental Populations and Phenotype Measurements
2.3. Genotyping of SNPs
2.4. Transcription Factor Binding Site Analysis
2.5. Statistical Analyses
3. Results
3.1. Identification of Genes Associated with Growth and Body Composition Traits in Broilers
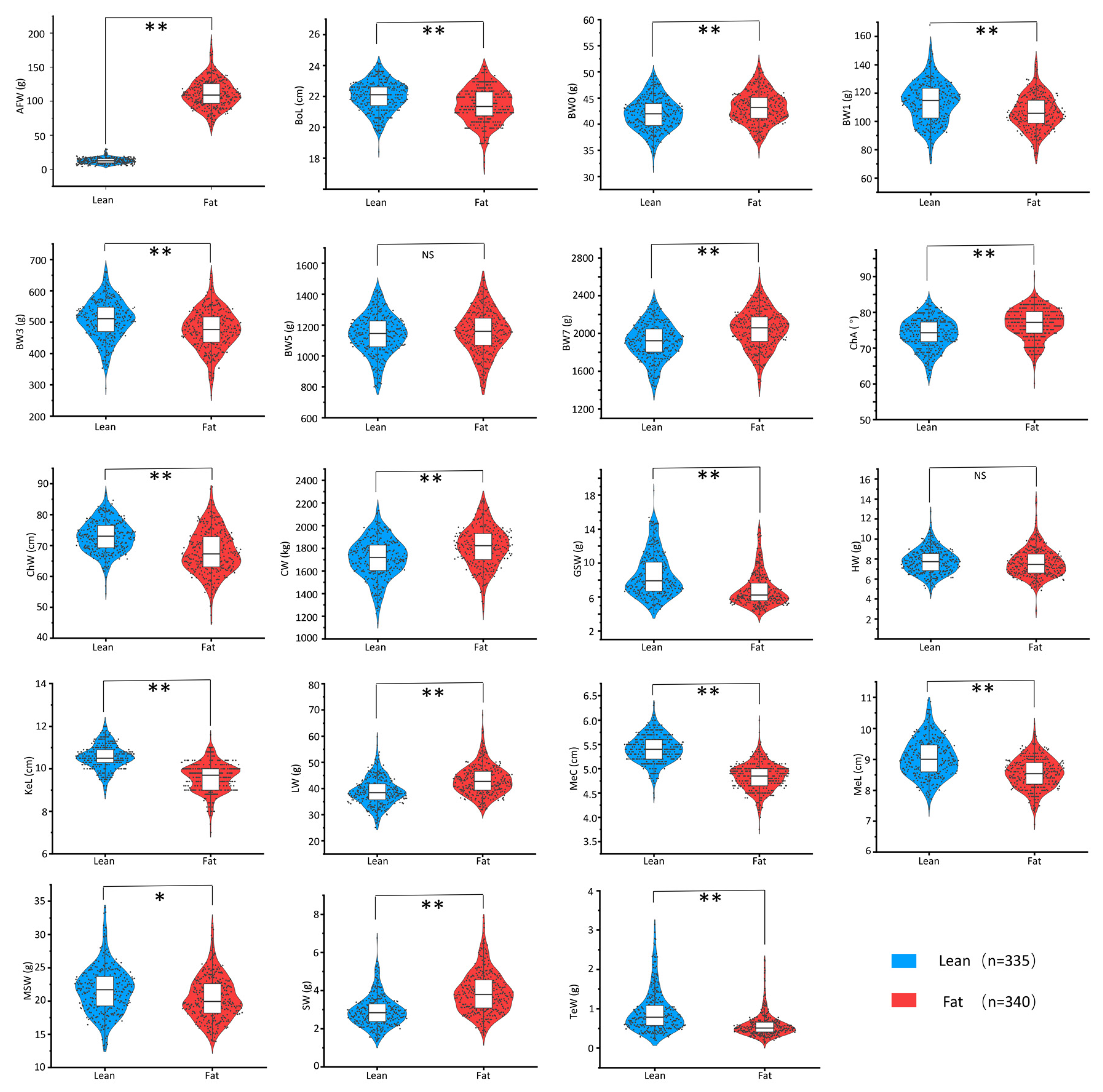
3.2. NEAUHLF Is an Ideal Test Material for Studying the Correlation between Growth and Body Composition Traits in Broilers
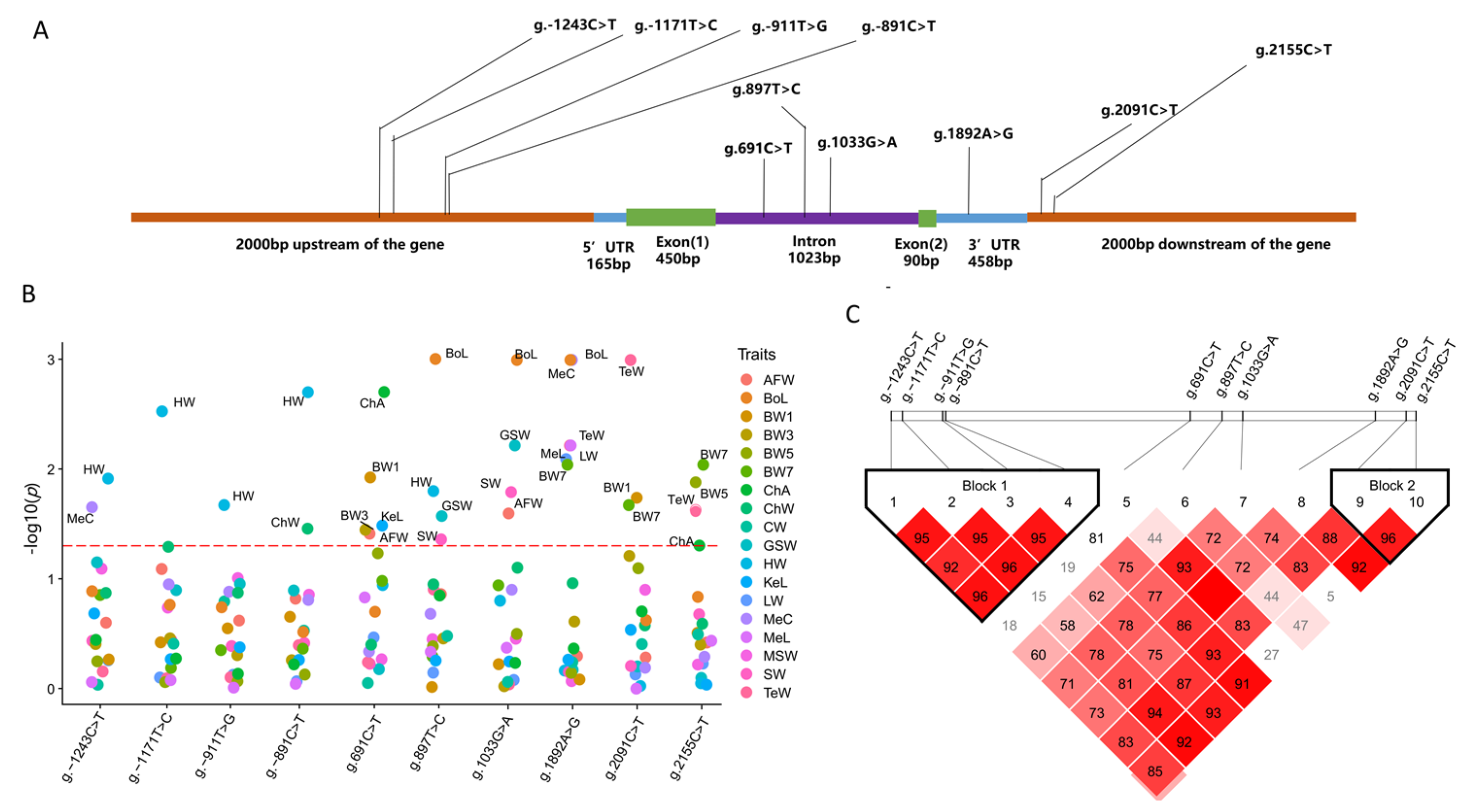
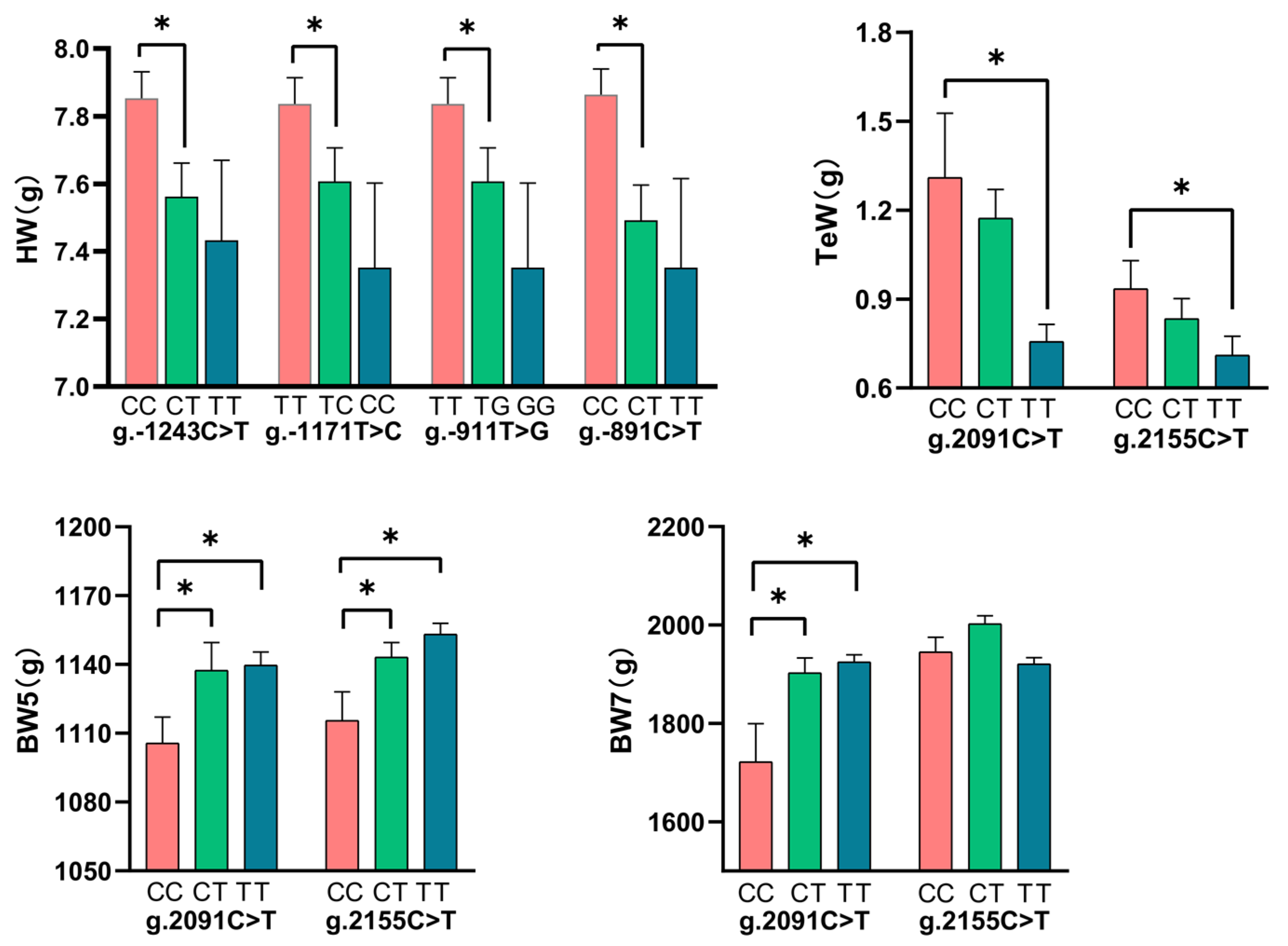
3.3. Associations of TCF21 Gene Polymorphisms with Growth and Body Composition Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dierick, E.; Hirvonen, O.P.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F.; Goossens, E. Rapid growth predisposes broilers to necrotic enteritis. Avian Pathol. 2019, 48, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Deeb, J.; Lamont, S. Genetic Architecture of Growth and Body Composition in Unique Chicken Populations. J. Hered. 2002, 93, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Khare, V.; Khare, A. Modern Approach in Animal Breeding by Use of Advanced Molecular Genetic Techniques. Int. J. Livest. Res. 2017, 7, 1–22. [Google Scholar] [CrossRef]
- Williams, J. The use of marker-assisted selection in animal breeding and biotechnology. Rev. Sci. Tech. Oie. 2005, 24, 379–391. [Google Scholar] [CrossRef]
- Wolc, A. Understanding genomic selection in poultry breeding. World Poult. Sci. J. 2014, 70, 309–314. [Google Scholar] [CrossRef] [Green Version]
- Vignal, A.; Milan, D.; SanCristobal, M.; Eggen, A. A review on SNP and other types of molecular markers and their use in animal genetics. Genet. Sel. Evol. 2002, 34, 275–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Wang, F.; Liu, L.; Li, Q.; Liu, R.; Zheng, M.; Cui, H.; Wen, J.; Zhao, G. Genetic Mutation Analysis of High and Low IgY Chickens by Capture Sequencing. Animals 2019, 9, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, B.; Li, Q.; Liu, R.; Zheng, M.; Wen, J.; Zhao, G. Genome-Wide Association Study of H/L Traits in Chicken. Animals 2019, 9, 260. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zheng, M.; Zhao, G.; Wang, J.; Liu, J.; Wang, S.; Feng, F.; Liu, D.; Zhu, D.; Li, Q.; et al. Identification of QTL regions and candidate genes for growth and feed efficiency in broilers. Genet. Sel. Evol. 2021, 53, 13. [Google Scholar] [CrossRef]
- Braitsch, C.; Combs, M.; Quaggin, S.; Yutzey, K. Pod1/Tcf21 is regulated by retinoic acid signaling and inhibits differentiation of epicardium-derived cells into smooth muscle in the developing heart. Dev. Biol. 2012, 368, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Na, W.; Zhang, H.-L.; Wang, N.; Du, Z.-Q.; Wang, S.-Z.; Wang, Z.-P.; Zhang, Z.; Li, H. TCF21 is related to testis growth and development in broiler chickens. Genet. Sel. Evol. 2017, 49, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Cheng, B.; Ma, Y.; Liu, Y.; Wang, N.; Zhang, H.; Li, Y.; Wang, Y.; Luan, P.; Cao, Z.; et al. Genome-wide survey and functional analysis reveal TCF21 promotes chicken preadipocyte differentiation by directly upregulating HTR2A. Biochem. Biophys. Res. Commun. 2022, 587, 131–138. [Google Scholar] [CrossRef]
- Kern, C.; Wang, Y.; Xu, X.; Pan, Z.; Halstead, M.; Chanthavixay, G.; Saelao, P.; Waters, S.; Xiang, R.; Chamberlain, A.; et al. Functional annotations of three domestic animal genomes provide vital resources for comparative and agricultural research. Nat. Commun. 2021, 12, 1821. [Google Scholar] [CrossRef]
- Quinlan, A.R. BEDTools: The Swiss-Army Tool for Genome Feature Analysis. Curr. Protoc. Bioinform. 2014, 47, 11–12. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; He, Q.-Y. ChIPseeker: An R/Bioconductor package for ChIP peak annotation, comparison and visualization. Bioinformatics 2015, 31, 2382–2383. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Hu, X.; Wang, Z.; Zhang, Y.; Wang, S.-Z.; Wang, N.; Leng, L.; Wang, S.; Wang, Q.; Wang, Y.; et al. Selection Signature Analysis Implicates the PC1/PCSK1 Region for Chicken Abdominal Fat Content. PLoS ONE 2012, 7, e40736. [Google Scholar] [CrossRef] [PubMed]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Bian, L.H.; Wang, S.Z.; Wang, Q.G.; Zhang, S.; Wang, Y.X.; Li, H. Variation at the insulin-like growth factor 1 gene and its association with body weight traits in the chicken. J. Anim. Breed. Genet. 2008, 125, 265–270. [Google Scholar] [CrossRef]
- Dybus, A. Associations between Leu/Val polymorphism of growth hormone gene and milk production traits in Black-and-White cattle. Arch. Anim. Breed. 2002, 45, 421–428. [Google Scholar] [CrossRef] [Green Version]
- Yano, Y.; Ozono, R.; Oishi, Y.; Kambe, M.; Yoshizumi, M.; Ishida, T.; Omura, S.; Oshima, T.; Igarashi, K. Genetic ablation of the transcription repressor Bach1 leads to myocardial protection against ischemia/reperfusion in mice. Genes Cells 2006, 11, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Sunagawa, Y.; Funamoto, M.; Honda, H.; Katanasaka, Y.; Murai, N.; Kawase, Y.; Hirako, Y.; Katagiri, T.; Yabe, H.; et al. The Selective Serotonin 2A Receptor Antagonist Sarpogrelate Prevents Cardiac Hypertrophy and Systolic Dysfunction via Inhibition of the ERK1/2-GATA4 Signaling Pathway. Pharmaceuticals 2021, 14, 1268. [Google Scholar] [CrossRef] [PubMed]
- Cicenia, M.; Alesi, V.; Orlando, V.; Magliozzi, M.; Di Tommaso, S.; Iodice, F.G.; Pompei, E.; Toscano, A.; Digilio, M.C.; Drago, F.; et al. 8p23.1 deletion: Look out for left ventricular hypertrabeculation and not only congenital heart diseases. Single-center experience and literature revision. Am. J. Med. Genet. A 2021. [Google Scholar] [CrossRef]
- Liang, Y.; Xu, Y.; Ding, L.; Chen, X.; Li, H. Urotensin II Induces Cardiac Fibrosis through the TGF-β/Smad Signaling Pathway during the Development of Cardiac Hypertrophy. Int. Heart J. 2021, 62, 1135–1144. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.M.; Wang, S.S.; Xu, Y.J.; Zhao, C.M.; Qiao, X.H.; Yang, C.X.; Liu, X.Y.; Yang, Y.Q. SOX17 Loss-of-Function Mutation Underlying Familial Pulmonary Arterial Hypertension. Int. Heart J. 2021, 62, 566–574. [Google Scholar] [CrossRef]
- Zhao, L.; Jiang, W.F.; Yang, C.X.; Qiao, Q.; Xu, Y.J.; Shi, H.Y.; Qiu, X.B.; Wu, S.H.; Yang, Y.Q. SOX17 loss-of-function variation underlying familial congenital heart disease. Eur. J. Med. Genet. 2021, 64, 104211. [Google Scholar] [CrossRef]
- Zerehdaran, S.; Vereijken, A.L.; van Arendonk, J.A.; van der Waaijt, E.H. Estimation of genetic parameters for fat deposition and carcass traits in broilers. Poult. Sci. 2004, 83, 521–525. [Google Scholar] [CrossRef]
- Xiang, H.; Gan, J.; Zeng, D.; Li, J.; Yu, H.; Zhao, H.; Yang, Y.; Tan, S.; Li, G.; Luo, C.; et al. Specific Microbial Taxa and Functional Capacity Contribute to Chicken Abdominal Fat Deposition. Front. Microbiol. 2021, 12, 643025. [Google Scholar] [CrossRef] [PubMed]
- Quaggin, S.E.; Schwartz, L.; Cui, S.; Igarashi, P.; Deimling, J.; Post, M.; Rossant, J. The basic-helix-loop-helix protein pod1 is critically important for kidney and lung organogenesis. Development 1999, 126, 5771–5783. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, N.; Walden, T.B.; Shabalina, I.G.; Timmons, J.A.; Cannon, B.; Nedergaard, J. Chronic peroxisome proliferator-activated receptor gamma (PPARgamma) activation of epididymally derived white adipocyte cultures reveals a population of thermogenically competent, UCP1-containing adipocytes molecularly distinct from classic brown adipocytes. J. Biol. Chem. 2010, 285, 7153–7164. [Google Scholar] [CrossRef] [Green Version]
- Akama, T.; Chun, T.H. Transcription factor 21 (TCF21) promotes proinflammatory interleukin 6 expression and extracellular matrix remodeling in visceral adipose stem cells. J. Biol. Chem. 2018, 293, 6603–6610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambeth, L.S.; Smith, C.A. Disorders of sexual development in poultry. Sex. Dev. 2012, 6, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.R.; Hirst, C.E.; Major, A.T.; Ezaz, T.; Ford, M.; Bibby, S.; Doran, T.J.; Smith, C.A. Gonadal and Endocrine Analysis of a Gynandromorphic Chicken. Endocrinology 2018, 159, 3492–3502. [Google Scholar] [CrossRef]
- Zheng, J.X.; Liu, Z.Z.; Yang, N. Deficiency of growth hormone receptor does not affect male reproduction in dwarf chickens. Poult. Sci. 2007, 86, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Ross, A.; Stallings, N.; Parker, K.L.; Capel, B.; Quaggin, S.E. Disrupted gonadogenesis and male-to-female sex reversal in Pod1 knockout mice. Development 2004, 131, 4095–4105. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, R.; Sadler-Riggleman, I.; Clement, T.; Skinner, M. Basic Helix-Loop-Helix Transcription Factor TCF21 Is a Downstream Target of the Male Sex Determining Gene SRY. PLoS ONE 2011, 6, e19935. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, R.; Schinke, E.; Haque, M.; Sadler-Riggleman, I.; Skinner, M. SRY Induced TCF21 Genome-Wide Targets and Cascade of bHLH Factors During Sertoli Cell Differentiation and Male Sex Determination in Rats. Biol. Reprod. 2012, 87, 131. [Google Scholar] [CrossRef]
- Lin, H.Y.; Chou, P.C.; Chen, Y.H.; Lai, L.S.; Chung, T.K.; Walzem, R.L.; Huang, S.Y.; Chen, S.E. Dietary Supplementation of 25-Hydroxycholecalciferol Improves Livability in Broiler Breeder Hens-Amelioration of Cardiac Pathogenesis and Hepatopathology. Animals 2019, 9, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, H.; Beaulaurier, J.; Lum, P.; Molony, C.; Yang, X.; MacNeil, D.; Weingarth, D.; Zhang, B.; Greenawalt, D.; Dobrin, R.; et al. Liver and Adipose Expression Associated SNPs Are Enriched for Association to Type 2 Diabetes. PLoS Genet. 2010, 6, e1000932. [Google Scholar] [CrossRef] [Green Version]
- Consortium, T.; Kanoni, S.; Willenborg, C.; Farrall, M.; Assimes, T.; Thompson, J.; Ingelsson, E.; Saleheen, D.; Erdmann, J.; Goldstein, B.; et al. Large-scale association analysis identifies new risk loci for coronary artery disease. Nat. Genet. 2012, 45, 25–33. [Google Scholar] [CrossRef]
- Hidai, H.; Bardales, R.; Goodwin, R.; Quertermous, T.; Quertermous, E. Cloning of capsulin, a basic helix-loop-helix factor expressed in progenitor cells of the pericardium and the coronary arteries1. Mech. Develop. 1998, 73, 33–43. [Google Scholar] [CrossRef]
- Mann, A.; Bhatia, S. Zebrafish: A Powerful Model for Understanding the Functional Relevance of Noncoding Region Mutations in Human Genetic Diseases. Biomedicines 2019, 7, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwan, A.R.; MacKenzie, A. Perspective: Quality Versus Quantity; Is It Important to Assess the Role of Enhancers in Complex Disease from an In Vivo Perspective? Int. J. Mol. Sci. 2020, 21, 7856. [Google Scholar] [CrossRef] [PubMed]
- Soltani-Fard, E.; Taghvimi, S.; Abedi Kichi, Z.; Weber, C.; Shabaninejad, Z.; Taheri-Anganeh, M.; Hossein Khatami, S.; Mousavi, P.; Movahedpour, A.; Natarelli, L. Insights into the Function of Regulatory RNAs in Bacteria and Archaea. Int. J. Transl. Med. 2021, 1, 403–423. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP ID | Primers (5′ to 3′) | Size of Product (bp) | Annealing Conditions | Number of Cyclesr | Endonuclease |
|---|---|---|---|---|---|
| g.-1243C>T (rs741031208) | F: CTGAATAGTTGGATTTTCCCCTGCC R:CTGATGGAGTCGAAGAGGGGTTTTA | 389 | 61 °C 35 s | 35 | HPyCH4IV |
| g.-1171T>G (rs312348545) | F:AGGTGTGTGAAGAAGGAAGGAGATACTGGGGAAGGC R:GATAAGGTCCCTGGCTGTGGGGCTGCATGC | 307 | 64 °C 30 s | 35 | HinP1I |
| g.-911T>G (rs739858013) | F: CCTTATCTTGCCTGTTTACTC R: GCACTTGGACCTCGCTATT | 379 | 55 °C 35 s | 35 | HinFI |
| g.-891>T (rs735029013) | F: GGAGCCCTCTCTTCCCCTCTTCTT R:CGTGCTGTCTAAACGCTGTCCTGTA | 332 | 61.8 °C 35 s | 35 | RsaI |
| g.691C>T (rs16282937) | F: TCCACTGGTCCCCACTGTCCCGT R: GGTTGGGCACAAAGCCTCAAAAGC | 273 | 57.8 °C 35 s | 35 | BstNI |
| g.897T>C (rs316577617) | F: TCCACTGGTCCCCACTGTCCCGT R: GGTTGGGCACAAAGCCTCAAAAGC | 273 | 60 °C 35 s | 35 | PvuII |
| g.1033G>A (rs16282934) | F: GTCCCCTCCACTGGTCCCCACTGT R: GGGAGTGCTTTCTGGTGTGGCCG | 394 | 60 °C 35 s | 40 | HinP1I |
| g.1892A>G (rs793941727) | F:TTGTCTGAGACCTGTGGAATATGTAGATGCCTTGA R:GGCAATAATCCTCAGCCCCACACCGA | 512 | 63.9 °C 30 s | 35 | BpuEI |
| g.2091C>T (rs314263759) | F: TACTTTTCGTTTCCAACTCACCAGG R:GACATCTTGTAAACAGTGCGGTCATAAC | 536 | 60 °C 30 s | 35 | AhdI |
| g.2155C>T (rs16282929) | F:ATTGAGTGCGTGTGCAGTCGAGTGTC R:CTCAGAGTTGACCCTCCTTGGGGAGTC | 432 | 62 °C 30 s | 35 | HpaII |
| SNPs | Strain | Genotype Frequency (No. of Birds) | Allele Frequency | χ2 | |||
|---|---|---|---|---|---|---|---|
| g.-1243C>T | CC | TC | TT | C | T | ||
| Lean line | 0.701 (235) | 0.278 (93) | 0.021 (7) | 0.84 | 0.16 | 21.66577 | |
| Fat line | 0.547 (186) | 0.379 (129) | 0.074 (25) | 0.737 | 0.263 | (p < 0.01) | |
| g.-1171T>C | TT | TC | CC | T | C | ||
| Lean line | 0.728 (244) | 0.266 (89) | 0.006 (2) | 0.861 | 0.139 | 28.45446 | |
| Fat line | 0.562 (190) | 0.367 (124) | 0.071 (24) | 0.746 | 0.254 | (p < 0.01) | |
| g.-911T>G | TT | TG | GG | T | G | ||
| Lean line | 0.697 (232) | 0.294 (98) | 0.009 (3) | 0.844 | 0.156 | 20.67613 | |
| Fat line | 0.56 (190) | 0.366 (124) | 0.074 (25) | 0.743 | 0.257 | (p < 0.01) | |
| g.-891C>T | CC | CT | TT | C | T | ||
| Lean line | 0.77 (258) | 0.227 (76) | 0.003 (1) | 0.884 | 0.116 | 39.40273 | |
| Fat line | 0.574 (195) | 0.356 (121) | 0.07 (24) | 0.751 | 0.249 | (p < 0.01) | |
| g.691C>T | CC | CT | TT | C | T | ||
| Lean line | 0.49 (164) | 0.409 (137) | 0.101 (34) | 0.694 | 0.306 | 103.2983 | |
| Fat line | 0.2 (68) | 0.438 (149) | 0.362 (123) | 0.419 | 0.581 | (p < 0.01) | |
| g.897T>C | CC | CT | TT | G | T | ||
| Lean line | 0 (1) | 0.3 (101) | 0.7 (230) | 0.155 | 0.845 | 133.2732 | |
| Fat line | 0.212 (72) | 0.465 (158) | 0.323 (110) | 0.444 | 0.556 | (p < 0.01) | |
| g.1033G>A | AA | AG | GG | A | G | ||
| Lean line | 0.62 (207) | 0.32 (101) | 0.06 (20) | 0.78 | 0.22 | 69.4667 | |
| Fat line | 0.34 (116) | 0.45 (151) | 0.21 (71) | 0.567 | 0.433 | (p < 0.01) | |
| g.1892A>G | AA | AG | GG | A | G | ||
| Lean line | 0.184 (59) | 0.763 (245) | 0.053 (17) | 0.565 | 0.435 | 38.83882 | |
| Fat line | 0.467 (154) | 0.527 (174) | 0.006 (2) | 0.73 | 0.27 | (p < 0.01) | |
| g.2091C>T | TT | TG | CC | T | C | ||
| Lean line | 0.8445 (277) | 0.137 (45) | 0.018 (6) | 0.909 | 0.091 | 61.71857 | |
| Fat line | 1 (340) | 0 (0) | 0 (0) | 1 | 0 | (p < 0.01) | |
| g.2155C>T | TT | TC | CC | T | C | ||
| Lean line | 0.728 (244) | 0.238 (80) | 0.033 (11) | 0.848 | 0.152 | 63.82669 | |
| Fat line | 0.45 (153) | 0.421 (143) | 0.129 (44) | 0.66 | 0.34 | (p < 0.01) | |
| Traits (Unit) | Heritability | Genetic Correlation |
|---|---|---|
| AFW (g) | 0.485 ± 0.096 | 1 |
| BoL (cm) | 0.069 ± 0.063 | −0.716 ± 0.308 |
| BW1 (g) | 0.500 ± 0.102 | −0.332 ± 0.160 |
| BW3 (g) | 0.218 ± 0.080 | −0.515 ± 0.194 |
| BW5 (g) | 0.304 ± 0.086 | −0.345 ± 0.200 |
| BW7 (g) | 0.224 ± 0.080 | −0.565 ± 0.190 |
| ChA (°) | 0.096 ± 0.043 | 0.213 ± 0.132 |
| ChW (cm) | 0.083 ± 0.062 | 0.240 ± 0.306 |
| CW (g) | 0.273 ± 0.091 | −0.155 ± 0.255 |
| GSW (g) | 0.537 ± 0.097 | −0.340 ± 0.150 |
| HW (g) | 0.187 ± 0.072 | 0.165 ± 0.216 |
| KeL (cm) | 0.193 ± 0.073 | −0.329 ± 0.204 |
| LW (g) | 0.131 ± 0.066 | 0.696 ± 0.223 |
| MeC (cm) | 0.250 ± 0.084 | −0.133 ± 0.198 |
| MeL (cm) | 0.176 ± 0.076 | −0.438 ± 0.208 |
| MSW (g) | 0.662 ± 0.094 | −0.181 ± 0.145 |
| TeW (g) | 0.413 ± 0.106 | 0.199 ± 0.238 |
| SNPs | Base Group | Transcription Factors | Function |
|---|---|---|---|
| g.-1243C>T | C | BACH1 | Myocardial ischemia [20] |
| g.-1171T>C | - | - | - |
| g.-911T>G | T | GATA4 | Key regulators of heart gene expression [21,22] |
| SMAD1 | protects cardiomyocytes from ischemia-reperfusion injury [23] | ||
| SOX17 | Early heart development in mouse embryos [24,25] | ||
| g.-891C>T | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, L.; Yu, J.; Ge, Y.; Li, H.; Li, Y.; Cao, Z.; Luan, P.; Xiao, F.; Gao, H.; Zhang, H. Associations of Transcription Factor 21 Gene Polymorphisms with the Growth and Body Composition Traits in Broilers. Animals 2022, 12, 393. https://doi.org/10.3390/ani12030393
Shen L, Yu J, Ge Y, Li H, Li Y, Cao Z, Luan P, Xiao F, Gao H, Zhang H. Associations of Transcription Factor 21 Gene Polymorphisms with the Growth and Body Composition Traits in Broilers. Animals. 2022; 12(3):393. https://doi.org/10.3390/ani12030393
Chicago/Turabian StyleShen, Linyong, Jiaqiang Yu, Yaowen Ge, Hui Li, Yumao Li, Zhiping Cao, Peng Luan, Fan Xiao, Haihe Gao, and Hui Zhang. 2022. "Associations of Transcription Factor 21 Gene Polymorphisms with the Growth and Body Composition Traits in Broilers" Animals 12, no. 3: 393. https://doi.org/10.3390/ani12030393
APA StyleShen, L., Yu, J., Ge, Y., Li, H., Li, Y., Cao, Z., Luan, P., Xiao, F., Gao, H., & Zhang, H. (2022). Associations of Transcription Factor 21 Gene Polymorphisms with the Growth and Body Composition Traits in Broilers. Animals, 12(3), 393. https://doi.org/10.3390/ani12030393