
Comprehensive Analyses of Bone and Cartilage Transcriptomes Evince Ion Transport, Inflammation and Cartilage Development-Related Genes Involved in Chickens’ Femoral Head Separation


 , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Bioinformatics, Differential Expression Analyses and Functional Annotation
2.2. qPCR Validation
2.3. Integrated Analysis
2.4. Polymorphism Identification Using the RNA-Seq Data
3. Results
3.1. RNA-Sequencing, Mapping and Characterization
3.2. Differentially Expressed Genes in AC and GP Transcriptomes
3.3. qPCR Validation
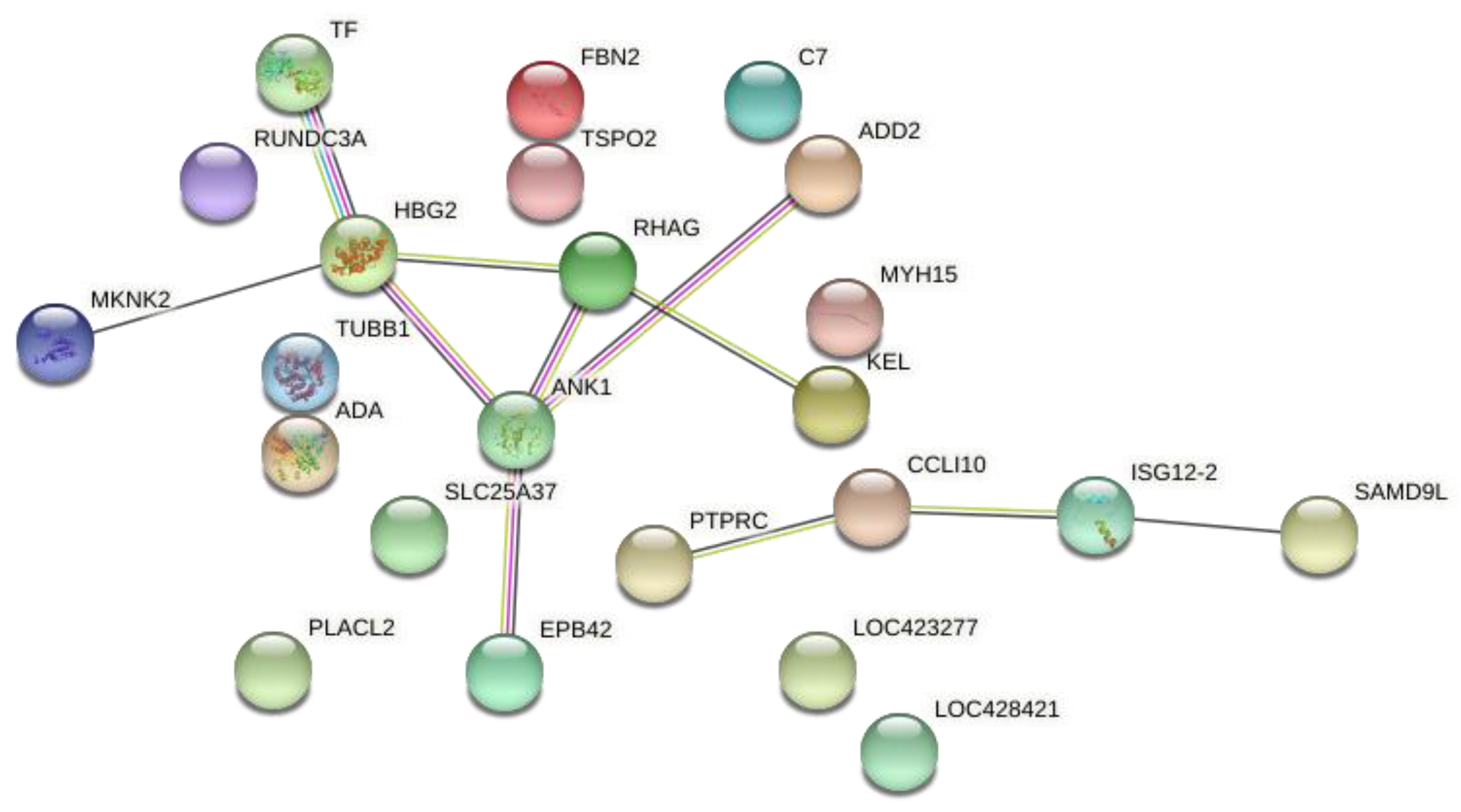
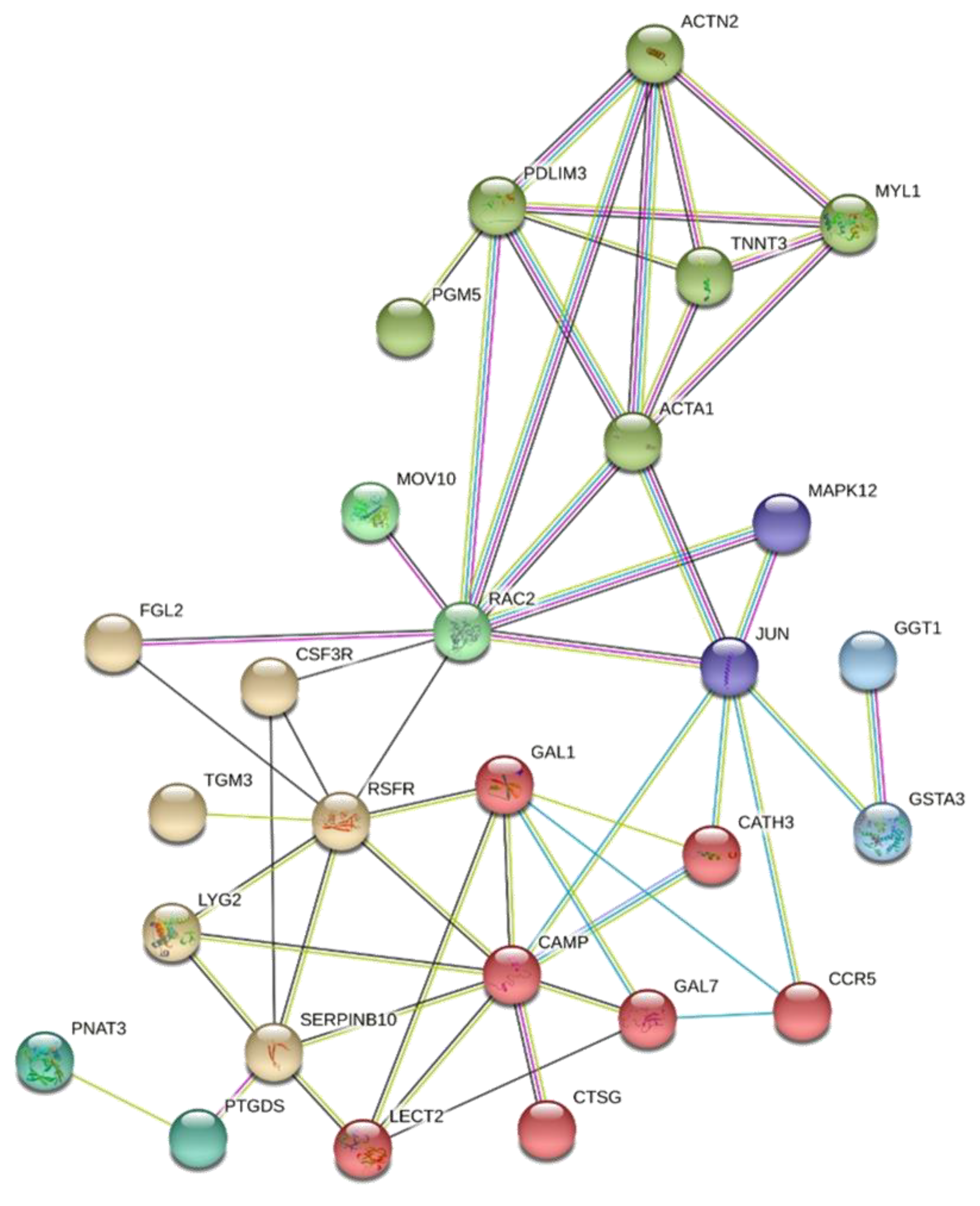
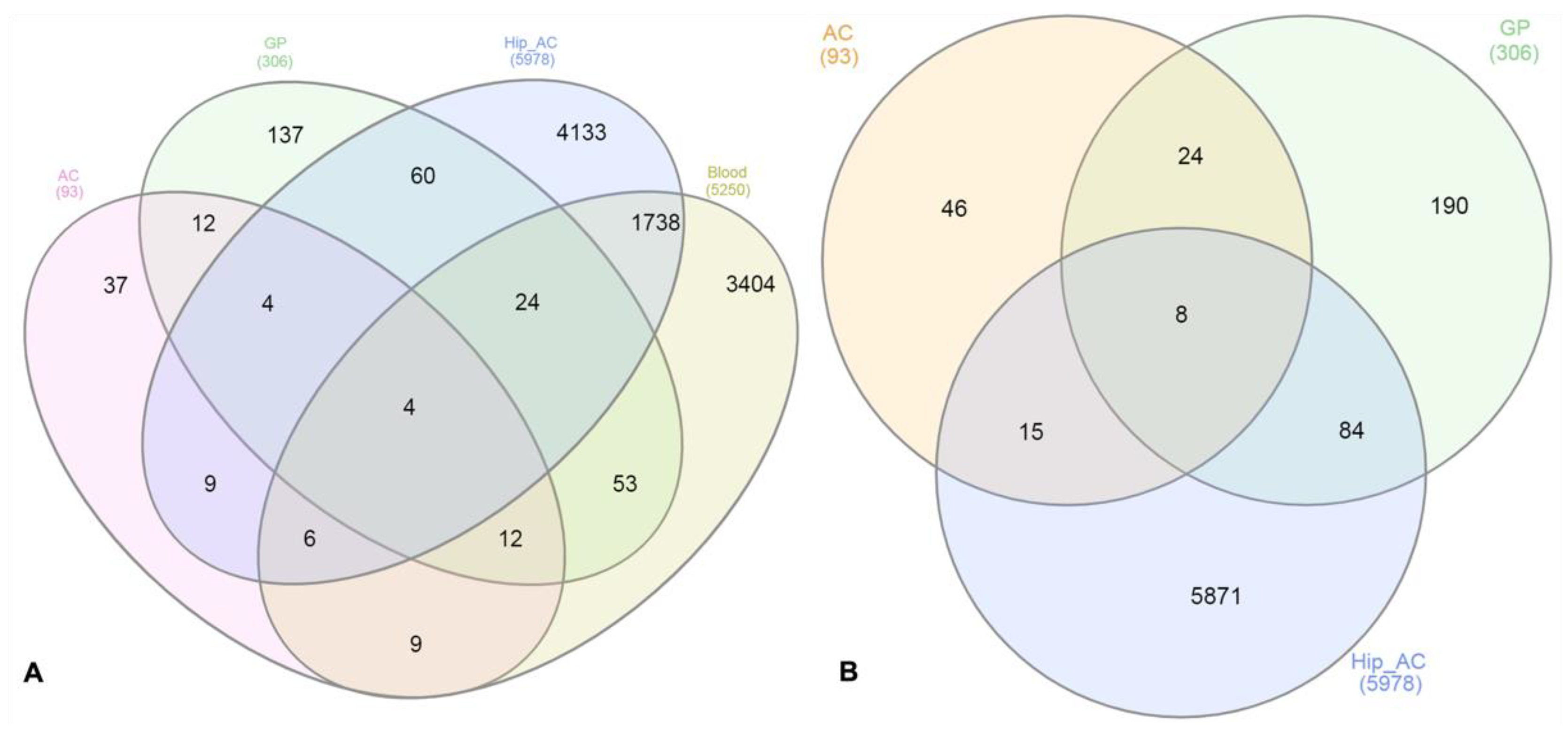
3.4. Comparison of the DE Genes from AC and GP Tissues with Different Datasets
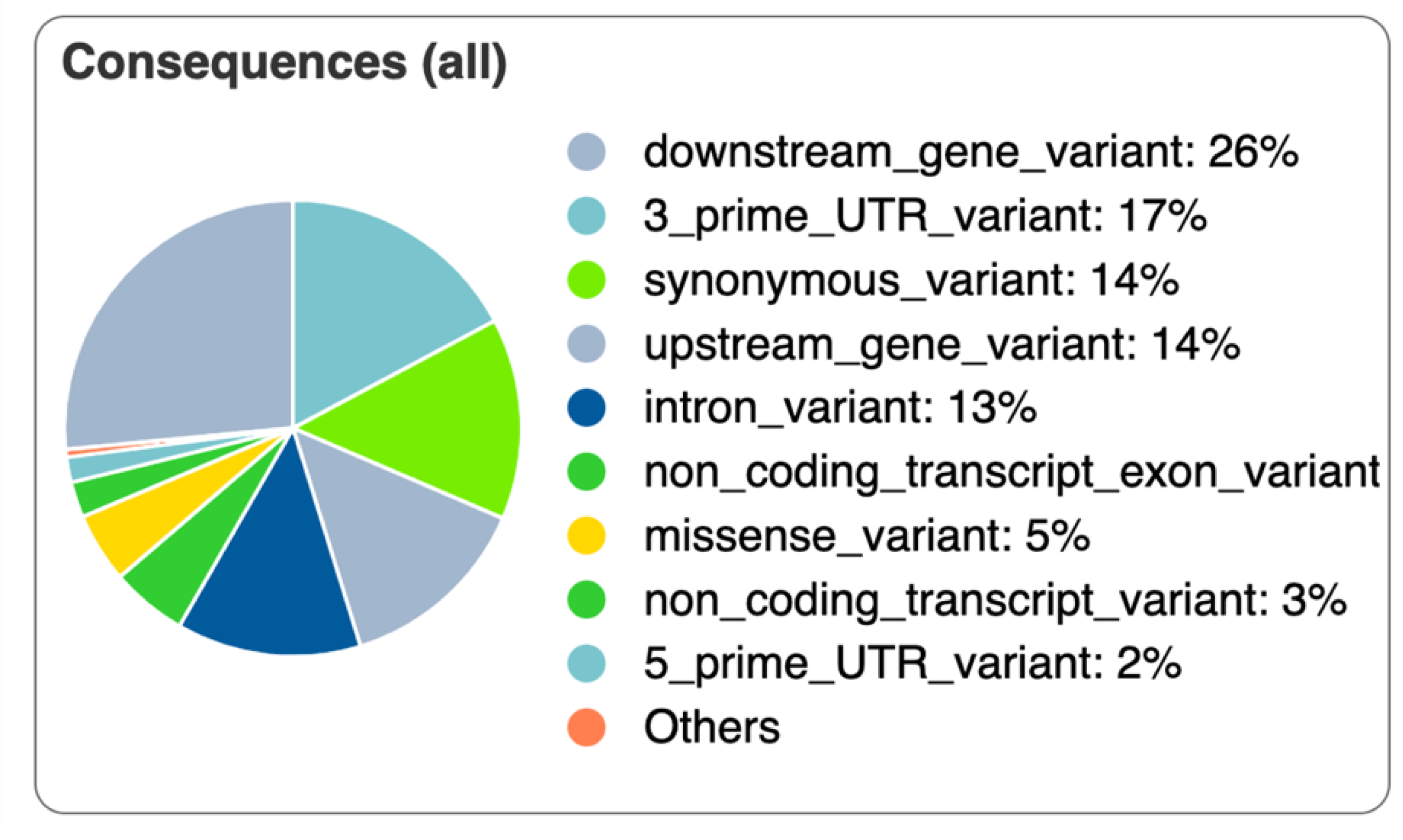
3.5. Variant Identification and Annotation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Voila, M.; Triches, D. A cadeia de carne de frango: Uma análise dos mercados brasileiro e mundial de 2002 a 2012. Rev. Teor. E Evidência Econômica 2015, 21, 126–148. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, D.; Berger, B.; Filiberto, D.; Newton, M.; Wolfe, B.; Karabinakis, E.; Clark, S.; Poon, E.; Abbett, E.; Nandagopal, S. Water resources: Agricultural and environmental issues. Bioscience 2004, 54, 909–918. [Google Scholar] [CrossRef] [Green Version]
- Barbut, S.; Sosnicki, A.A.; Lonergan, S.M.; Knapp, T.; Ciobanu, D.C.; Gatcliffe, L.J.; Huff-Lonergan, E.; Wilson, E.W. Progress in reducing the pale, soft and exudative (PSE) problem in pork and poultry meat. Meat Sci. 2008, 79, 46–63. [Google Scholar] [CrossRef]
- Knowles, T.G.; Kestin, S.C.; Haslam, S.M.; Brown, S.N.; Green, L.E.; Butterworth, A.; Pope, S.J.; Pfeiffer, D.; Nicol, C.J. Leg Disorders in Broiler Chickens: Prevalence, Risk Factors and Prevention. PLoS ONE 2008, 3, e1545. [Google Scholar] [CrossRef] [Green Version]
- Tallentire, C.W.; Leinonen, I.; Kyriazakis, I. Breeding for efficiency in the broiler chicken: A review. Agron. Sustain. Dev. 2016, 36, 66. [Google Scholar] [CrossRef] [Green Version]
- Paz, I.C.D.L.A. Problemas locomotores em frangos de corte—Revisão./locomotor problems in broiler chickens—A review. Rev. Bras. Eng. Biossistemas 2008, 2, 263. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, R.H.; Kirkden, R.D.; Broom, D.M. A Review of the Aetiology and Pathology of Leg Weakness in Broilers in Relation to Welfare. Avian Poult. Biol. Rev. 2002, 13, 45–103. [Google Scholar] [CrossRef] [Green Version]
- Wideman, R.F. Bacterial chondronecrosis with osteomyelitis and lameness in broilers: A review. Poult. Sci. 2016, 95, 325–344. [Google Scholar] [CrossRef]
- Packialakshmi, B.; Rath, N.C.; Huff, W.E.; Huff, G.R. Poultry Femoral Head Separation and Necrosis: A Review. Avian Dis. 2015, 59, 349–354. [Google Scholar] [CrossRef]
- Li, P.F.; Zhou, Z.L.; Shi, C.Y.; Hou, J.F. Downregulation of basic fibroblast growth factor is associated with femoral head necrosis in broilers. Poult. Sci. 2015, 94, 1052–1059. [Google Scholar] [CrossRef]
- Durairaj, V.; Okimoto, R.; Rasaputra, K.; Clark, F.D.; Rath, N.C. Histopathology and Serum Clinical Chemistry Evaluation of Broilers With Femoral Head Separation Disorder. Avian Dis. 2009, 53, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Wideman Bone circulatory disturbances in the development of spontaneous bacterial chondronecrosis with osteomyelitis: A translational model for the pathogenesis of femoral head necrosis. Front. Endocrinol. (Lausanne) 2013, 3, 183. [CrossRef] [Green Version]
- Paludo, E.; Ibelli, A.M.G.; Peixoto, J.O.; Tavernari, F.C.; Lima-Rosa, C.A.V.; Pandolfi, J.R.C.; Ledur, M.C. The involvement of RUNX2 and SPARC genes in the bacterial chondronecrosis with osteomyelitis in broilers. Animal 2017, 11, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Petry, B.; Savoldi, I.R.; Ibelli, A.M.G.; Paludo, E.; de Oliveira Peixoto, J.; Jaenisch, F.R.F.; de Córdova Cucco, D.; Ledur, M.C. New genes involved in the Bacterial Chondronecrosis with Osteomyelitis in commercial broilers. Livest. Sci. 2018, 208, 33–39. [Google Scholar] [CrossRef]
- Peixoto, J.D.O.; Savoldi, I.R.; Ibelli, A.M.G.; Cantão, M.E.; Jaenisch, F.R.F.; Giachetto, P.F.; Settles, M.L.; Zanella, R.; Marchesi, J.A.P.; Pandolfi, J.R.; et al. Proximal femoral head transcriptome reveals novel candidate genes related to epiphysiolysis in broiler chickens. BMC Genom. 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- de Peixoto, J.O. Improving skeletal structure in meat-type chickens given fast weight gain and large breast yield through genetic selection. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2020, 15. [Google Scholar] [CrossRef]
- Hul, L.M.; Ibelli, A.M.G.; Savoldi, I.R.; Marcelino, D.E.P.; Fernandes, L.T.; Peixoto, J.O.; Cantão, M.E.; Higa, R.H.; Giachetto, P.F.; Coutinho, L.L.; et al. Differentially expressed genes in the femur cartilage transcriptome clarify the understanding of femoral head separation in chickens. Sci. Rep. 2021, 11, 17965. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, S.; Huang, J.; Guo, W.; Chen, J.; Zhang, L.; Zhao, B.; Peng, J.; Wang, A.; Wang, Y.; et al. The ECM-cell interaction of cartilage extracellular matrix on chondrocytes. BioMed Res. Int. 2014, 2014, 648459. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. Revigo summarizes and visualizes long lists of gene ontology terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heberle, H.; Meirelles, V.G.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Kestin, S.C.; Gordon, S.; Su, G.; Sørensen, P. Relationships in broiler chickens between lameness, liveweight, growth rate and age. Vet. Rec. 2001, 148, 195–197. [Google Scholar] [CrossRef]
- Jiang, T.; Mandal, R.K.; Wideman, R.F.; Khatiwara, A.; Pevzner, I.; Min Kwon, Y. Molecular Survey of Bacterial Communities Associated with Bacterial Chondronecrosis with Osteomyelitis (BCO) in Broilers. PLoS ONE 2015, 10, e0124403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Q.; Liu, Z.; Shen, C.; Li, H.; Ding, J.; Jin, F.; Sha, L.; Zhang, Z. Microarray study of gene expression profile to identify new candidate genes involved in the molecular mechanism of leptin-induced knee joint osteoarthritis in rat. Hereditas 2018, 155, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcantara, J.; Schryvers, A.B. Transferrin binding protein two interacts with both the N-lobe and C-lobe of ovotransferrin. Microb. Pathog. 1996, 20, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Bullen, J.J.; Rogers, H.J.; Griffiths, E. Role of iron in bacterial infection. Curr. Top. Microbiol. Immunol. 1978, 80, 1–35. [Google Scholar] [PubMed]
- Giansanti, F.; Leboffe, L.; Angelucci, F.; Antonini, G. The nutraceutical properties of ovotransferrin and its potential utilization as a functional food. Nutrients 2015, 7, 9105–9115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlevaro, M.F.; Albini, A.; Ribatti, D.; Gentili, C.; Benelli, R.; Cermelli, S.; Cancedda, R.; Cancedda, F.D. Transferrin promotes endothelial cell migration and invasion: Implication in cartilage neovascularization. J. Cell Biol. 1997, 136, 1375–1384. [Google Scholar] [CrossRef]
- HBG2 Gene—GeneCards | HBG2 Protein | HBG2 Antibody.
- Alev, C.; Shinmyozu, K.; McIntyre, B.A.S.; Sheng, G. Genomic organization of zebra finch alpha and beta globin genes and their expression in primitive and definitive blood in comparison with globins in chicken. Dev. Genes Evol. 2009, 219, 353–360. [Google Scholar] [CrossRef] [PubMed]
- SLC25A37 Gene—GeneCards | MFRN1 Protein | MFRN1 Antibody.
- Pizzagalli, M.D.; Bensimon, A.; Superti-Furga, G. A guide to plasma membrane solute carrier proteins. FEBS J. 2021, 288, 2784–2835. [Google Scholar] [CrossRef] [PubMed]
- Chigaev, A. Does aberrant membrane transport contribute to poor outcome in adult acute myeloid leukemia? Front. Pharmacol. 2015, 6, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Luo, Q.; Shu, Y.; Zeng, Z.; Huang, B.; Feng, Y.; Zhang, B.; Wang, X.; Lei, Y.; Ye, Z.; et al. Transcriptomic landscape regulated by the 14 types of bone morphogenetic proteins (BMPs) in lineage commitment and differentiation of mesenchymal stem cells (MSCs). Genes Dis. 2019, 6, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Michaely, P.; Tomchick, D.R.; Machius, M.; Anderson, R.G.W. Crystal structure of a 12 ANK repeat stack from human ankyrinR. EMBO J. 2002, 21, 6387–6396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagnato, P.; Barone, V.; Giacomello, E.; Rossi, D.; Sorrentino, V. Binding of an ankyrin-1 isoform to obscurin suggests a molecular link between the sarcoplasmic reticulum and myofibrils in striated muscles. J. Cell Biol. 2003, 160, 245–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nummenmaa, E.; Hämäläinen, M.; Pemmari, A.; Moilanen, L.J.; Tuure, L.; Nieminen, R.M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Transient Receptor Potential Ankyrin 1 (TRPA1) Is Involved in Upregulating Interleukin-6 Expression in Osteoarthritic Chondrocyte Models. Int. J. Mol. Sci. 2020, 22, 87. [Google Scholar] [CrossRef] [PubMed]
- Scheper, G.C.; Parra, J.L.; Wilson, M.; van Kollenburg, B.; Vertegaal, A.C.O.; Han, Z.-G.; Proud, C.G. The N and C Termini of the Splice Variants of the Human Mitogen-Activated Protein Kinase-Interacting Kinase Mnk2 Determine Activity and Localization. Mol. Cell. Biol. 2003, 23, 5692–5705. [Google Scholar] [CrossRef] [Green Version]
- Chrestensen, C.A.; Eschenroeder, A.; Ross, W.G.; Ueda, T.; Watanabe-Fukunaga, R.; Fukunaga, R.; Sturgill, T.W. Loss of MNK function sensitizes fibroblasts to serum-withdrawal induced apoptosis. Genes Cells 2007, 12, 1133–1140. [Google Scholar] [CrossRef] [PubMed]
- Daneshyar, M.; Kermanshahi, H.; Golian, A. The effects of turmeric supplementation on antioxidant status, blood gas indices and mortality in broiler chickens with T3-induced ascites. Br. Poult. Sci. 2012, 53, 379–385. [Google Scholar] [CrossRef]
- Acharya, K.R.; Ackerman, S.J. Eosinophil Granule Proteins: Form and Function. J. Biol. Chem. 2014, 289, 17406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davoine, F.; Lacy, P. Eosinophil cytokines, chemokines, and growth factors: Emerging roles in immunity. Front. Immunol. 2014, 5, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jong, E.C.; Henderson, W.R.; Klebanoff, S.J. Bactericidal activity of eosinophil peroxidase. J. Immunol. 1980, 124, 1378–1382. [Google Scholar] [PubMed]
- Johnston, R.B.; Baehner, R.L. Chronic granulomatous disease: Correlation between pathogenesis and clinical findings. Pediatrics 1971, 48, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Zhang, D.E.; Sun, Z.; Albee, E.A.; Nagata, S.; Tenen, D.G.; Ackerman, S.J. Functional characterization of the promoter for the gene encoding human eosinophil peroxidase. J. Biol. Chem. 1994, 269, 19410–19419. [Google Scholar] [CrossRef]
- Celi, P.; Verlhac, V.; Pérez Calvo, E.; Schmeisser, J.; Kluenter, A.M. Biomarkers of gastrointestinal functionality in animal nutrition and health. Anim. Feed Sci. Technol. 2019, 250, 9–31. [Google Scholar] [CrossRef]
- Hay, E.D. Collagen and Other Matrix Glycoproteins in Embryogenesis. In Cell Biology of Extracellular Matrix; Springer US: New York, NY, USA, 1991; pp. 419–462. [Google Scholar]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velleman, S.G. The Role of the Extracellular Matrix in Skeletal Muscle Development. Poult. Sci. 2000, 79, 985–989. [Google Scholar] [CrossRef] [PubMed]
- Aran, J.M.; Colomer, D.; Matutes, E.; Vives-Corrons, J.L.; Franco, R. Presence of adenosine deaminase on the surface of mononuclear blood cells: Immunochemical localization using light and electron microscopy. J. Histochem. Cytochem. 1991, 39, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Desrosiers, M.D.; Cembrola, K.M.; Fakir, M.J.; Stephens, L.A.; Jama, F.M.; Shameli, A.; Mehal, W.Z.; Santamaria, P.; Shi, Y. Adenosine Deamination Sustains Dendritic Cell Activation in Inflammation. J. Immunol. 2007, 179, 1884–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitkovsky, M.V.; Lukashev, D.; Apasov, S.; Kojima, H.; Koshiba, M.; Caldwell, C.; Ohta, A.; Thiel, M. Physiological Control of Immune Response and Inflammatory Tissue Damage by Hypoxia Inducible Factors and Adenosine A2A Receptors. Annu. Rev. Immunol. 2004, 22, 657–682. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sharma, A. Adenosine: An endogenous modulator of innate immune system with therapeutic potential. Eur. J. Pharmacol. 2009, 616, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.H.; Alabanza, L.M.; Mahamed, D.A.; Bynoe, M.S. Extracellular adenosine signaling induces CX3CL1 expression in the brain to promote experimental autoimmune encephalomyelitis. J. Neuroinflammation 2012, 9, 683. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, Y.; Luo, W.; Song, X.; Huang, L.; Xiao, J.; Jin, F.; Ren, Z.; Wang, Y. Roles of long non-coding RNAs and emerging RNA-binding proteins in innate antiviral responses. Theranostics 2020, 10, 9407–9424. [Google Scholar] [CrossRef]
- Strazzulla, L.C.; Cronstein, B.N. Regulation of bone and cartilage by adenosine signaling. Purinergic Signal. 2016, 12, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Manson, D.; Diamond, L.; Oudjhane, K.; Hussain, F.B.; Roifman, C.; Grunebaum, E. Characteristic scapular and rib changes on chest radiographs of children with ADA-deficiency SCIDS in the first year of life. Pediatr. Radiol. 2013, 43, 589–592. [Google Scholar] [CrossRef]
- Sauer, A.V.; Mrak, E.; Hernandez, R.J.; Zacchi, E.; Cavani, F.; Casiraghi, M.; Grunebaum, E.; Roifman, C.M.; Cervi, M.C.; Ambrosi, A.; et al. ADA-deficient SCID is associated with a specific microenvironment and bone phenotype characterized by RANKL/OPG imbalance and osteoblast insufficiency. Blood 2009, 114, 3216–3226. [Google Scholar] [CrossRef] [PubMed]
- ADA Gene—GeneCards | ADA Protein | ADA Antibody.
- Ananko, E.A.; Kondrakhin, Y.V.; Merkulova, T.I.; Kolchanov, N.A. Recognition of interferon-inducible sites, promoters, and enhancers. BMC Bioinform. 2007, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Tillgren, V.; Ho, J.C.S.; Önnerfjord, P.; Kalamajski, S. The Novel Small Leucine-rich Protein Chondroadherin-like (CHADL) Is Expressed in Cartilage and Modulates Chondrocyte Differentiation. J. Biol. Chem. 2015, 290, 918–925. [Google Scholar] [CrossRef] [Green Version]
- Kania, K.; Colella, F.; Riemen, A.H.K.; Wang, H.; Howard, K.A.; Aigner, T.; Dell’Accio, F.; Capellini, T.D.; Roelofs, A.J.; De Bari, C. Regulation of Gdf5 expression in joint remodelling, repair and osteoarthritis. Sci. Rep. 2020, 10, 157. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.X.; Li, Y.S.; Tu, C.; Xie, P.; Li, Y.H.; Qi, L.; Li, Z.H. Identification of a potential gene target for osteoarthritis based on bioinformatics analyses. J. Orthop. Surg. Res. 2020, 15, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Coorens, M.; Banaschewski, B.J.H.; Baer, B.J.; Yamashita, C.; van Dijk, A.; Haagsman, H.P.; Veldhuizen, R.A.W.; Veldhuizen, E.J.A. Killing of Pseudomonas aeruginosa by Chicken Cathelicidin-2 Is Immunogenically Silent, Preventing Lung Inflammation In Vivo. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Ensembl ID | Primer (5′ to 3′) |
|---|---|---|
| C7 | ENSGALG00000014835 | F: GTGGTCCTTCCCTGGACATC |
| Gallus gallus complement C7 | R: GTTGGATCGCGCTTCACATT | |
| EPB42 | ENSGALG00000021230 | F: GCTACAGACCTGGTACTTGGA |
| Gallus gallus erythrocyte membrane protein band 4.2 | R: GACTTGCAGGGTTCTAACTTCG | |
| MKNK2 | ENSGALG00000003845 | F: CATCCTGCAGAGGAACAGCA |
| Gallus gallus MAPK interacting serine/threonine kinase 2 | R: CCTGGAGGCTGCTTTGATGA | |
| SLC4A1 | ENSGALG00000039978 | F:TCGAGCTCAAATGCCTGGAC |
| Gallus gallus solute carrier family 4 member 1 (Diego blood group) | R: GGCATCTGCACCTCGTTGTA | |
| SPTB | ENSGALG00000036805 | F: CTGGATGCTGATGAGGCCAA |
| Gallus gallus spectrin beta, erythrocytic | R: CTTTCCTCGTCCTTCGGCTT | |
| ANK1 * | ENSGALG00000029534 | F: CCACCATCCCACCATTCAGT |
| Ankyrin 1 | R: ACGGTCACAAACTCCAGCAT | |
| ADA * | ENSGALG00000004170 | F:TTCGGCAAGAAAAGAGGGGT |
| adenosine deaminase | R: GTGTTTGGTAGCTGACGTGC | |
| RHAG * | ENSGALG00000016684 | F:TCTGGAGATCACGGCCTTTG |
| Rh associated glycoprotein | R:GCTCCAATATCTGTGGCCTGA |
| AC | GP | |||
|---|---|---|---|---|
| Annotated Transcripts | Transcript Number | % | Transcript Number | % |
| IG_V_gene | 3 | 0.02% | 20 | 0.16% |
| LncRNA | 413 | 3.40% | 454 | 3.59% |
| MiRNA | 20 | 0.16% | 22 | 0.17% |
| misc_RNA | 1 | 0.01% | 2 | 0.02% |
| Mt_Rrna | 2 | 0.02% | 2 | 0.02% |
| protein_coding | 11,577 | 95.03% | 11,942 | 94.54% |
| Pseudogene | 155 | 1.27% | 162 | 1.28% |
| rRNA | 1 | 0.01% | 0 | 0 |
| scaRNA | 1 | 0.01% | 0 | 0 |
| snoRNA | 8 | 0.07% | 26 | 0.21% |
| snRNA | 1 | 0.01% | 1 | 0.01% |
| sRNA | 1 | 0.01% | ||
| Total annotated transcripts | 12,182 | 12,632 | ||
| AC | GP | ||||
|---|---|---|---|---|---|
| Ensembl ID | Gene Name | logFC | FDR | logFC | FDR |
| ENSGALG00000043254 | EPX | 4.25 | 4.11 × 10−9 | 2.59 | 0.0009 |
| ENSGALG00000003212 | TSPO2 | 3.66 | 7.20 × 10−5 | 1.54 | 0.005 |
| ENSGALG00000040279 | RHCE | 2.02 | 0.0008 | 1.29 | 0.002 |
| ENSGALG00000026518 | RUNDC3A | 2.44 | 0.003 | 1.10 | 0.02 |
| ENSGALG00000039978 | SLC4A1 | 2.70 | 0.003 | 1.20 | 0.01 |
| ENSGALG00000007447 | TUBB1 | 1.99 | 0.003 | 1.28 | 0.01 |
| ENSGALG00000011190 | PLACL2 | 2.54 | 0.005 | 1.10 | 0.0002 |
| ENSGALG00000042105 | PLCB2 | 1.81 | 0.005 | 0.67 | 0.03 |
| ENSGALG00000014585 | CCL26 | 2.36 | 0.008 | 1.12 | 0.04 |
| ENSGALG00000021230 | EPB42 | 2.69 | 0.008 | 1.33 | 0.02 |
| ENSGALG00000004170 | ADA | 1.71 | 0.008 | 1.01 | 0.001 |
| ENSGALG00000015358 | MYH15 | −1.91 | 0.01 | −1.38 | 0.03 |
| ENSGALG00000016684 | RHAG | 2.43 | 0.01 | 1.34 | 0.006 |
| ENSGALG00000029857 | GIMAP6 | 1.79 | 0.01 | 0.76 | 0.01 |
| ENSGALG00000013575 | IFI6 | 2.87 | 0.01 | 2.22 | 0.01 |
| ENSGALG00000009479 | SAMD9L | 3.10 | 0.01 | 1.78 | 0.01 |
| ENSGALG00000000378 | SLC25A37 | 1.76 | 0.01 | 1.32 | 0.001 |
| ENSGALG00000041693 | 2.81 | 0.01 | 1.33 | 0.04 | |
| ENSGALG00000003845 | MKNK2 | 1.40 | 0.01 | 0.63 | 0.009 |
| ENSGALG00000026948 | ADD2 | 2.12 | 0.01 | 1.27 | 1.85 |
| ENSGALG00000036805 | SPTB | 2.19 | 0.01 | 1.09 | 0.005 |
| ENSGALG00000051068 | 2.38 | 0.02 | 1.41 | 0.0005 | |
| ENSGALG00000003594 | ANK1 | 2.11 | 0.02 | 0.99 | 0.01 |
| ENSGALG00000014736 | KEL | 1.99 | 0.02 | 1.21 | 0.005 |
| ENSGALG00000043671 | −1.24 | 0.02 | −1.03 | 0.02 | |
| ENSGALG00000028273 | HBE1 | 3.74 | 0.02 | 2.24 | 0.04 |
| ENSGALG00000039269 | RNF213 | 2.06 | 0.027 | 1.06 | 0.03 |
| ENSGALG00000014835 | C7 | 2.84 | 0.027 | 1.55 | 0.001 |
| ENSGALG00000028357 | LOC428421 | 2.50 | 0.036 | 1.49 | 0.006 |
| ENSGALG00000002192 | PTPRC | 1.22 | 0.037 | 0.61 | 0.03 |
| ENSGALG00000045776 | CPN2 | 1.77 | 0.037 | 0.90 | 0.02 |
| ENSGALG00000014686 | FBN2 | −1.08 | 0.037 | −1.72 | 0.0005 |
| ENSGALG00000030247 | TMOD4 | 2.05 | 0.04 | 1.06 | 0.01 |
| ENSGALG00000009552 | LOC423277 | 1.78 | 0.04 | 1.14 | 0.04 |
| ENSGALG00000044326 | LOC426820 | 1.54 | 0.04 | 0.80 | 0.01 |
| ENSGALG00000006453 | TF | 3.04 | 0.04 | 0.67 | 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldoni, I.; Ibelli, A.M.G.; Fernandes, L.T.; Peixoto, J.d.O.; Hul, L.M.; Cantão, M.E.; Gouveia, J.J.d.S.; Ledur, M.C. Comprehensive Analyses of Bone and Cartilage Transcriptomes Evince Ion Transport, Inflammation and Cartilage Development-Related Genes Involved in Chickens’ Femoral Head Separation. Animals 2022, 12, 788. https://doi.org/10.3390/ani12060788
Goldoni I, Ibelli AMG, Fernandes LT, Peixoto JdO, Hul LM, Cantão ME, Gouveia JJdS, Ledur MC. Comprehensive Analyses of Bone and Cartilage Transcriptomes Evince Ion Transport, Inflammation and Cartilage Development-Related Genes Involved in Chickens’ Femoral Head Separation. Animals. 2022; 12(6):788. https://doi.org/10.3390/ani12060788
Chicago/Turabian StyleGoldoni, Iara, Adriana Mércia Guaratini Ibelli, Lana Teixeira Fernandes, Jane de Oliveira Peixoto, Ludmila Mudri Hul, Maurício Egídio Cantão, João José de Simoni Gouveia, and Mônica Corrêa Ledur. 2022. "Comprehensive Analyses of Bone and Cartilage Transcriptomes Evince Ion Transport, Inflammation and Cartilage Development-Related Genes Involved in Chickens’ Femoral Head Separation" Animals 12, no. 6: 788. https://doi.org/10.3390/ani12060788
APA StyleGoldoni, I., Ibelli, A. M. G., Fernandes, L. T., Peixoto, J. d. O., Hul, L. M., Cantão, M. E., Gouveia, J. J. d. S., & Ledur, M. C. (2022). Comprehensive Analyses of Bone and Cartilage Transcriptomes Evince Ion Transport, Inflammation and Cartilage Development-Related Genes Involved in Chickens’ Femoral Head Separation. Animals, 12(6), 788. https://doi.org/10.3390/ani12060788