
Reduction in Pathogenicity in Yeast-like Fungi by Farnesol in Quail Model

 ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain
2.2. Reagents
2.3. C. albicans Processing with Farnesol
2.4. Experimental Infection Quails with Candida albicans ATCC 10231
2.4.1. Animals
2.4.2. Hematological Analysis of Blood Samples
2.4.3. Serum Biochemical Analysis
2.5. Changes in Microbial Composition in Quail Feces
2.6. Statistics
3. Results
3.1. Phenotypic Characteristics of C. albicans Strain Used in the Study
3.2. Determination of Biofilm-Forming Potential of C. albicans
3.3. Experimental Infection in Quails with Candida albicans ATCC 10231
3.3.1. Histopathology of Quail Candida albicans Infection
3.3.2. Hematological and Biochemical Analysis in the Experiment and Control Groups
3.4. Changes in Microbial Composition in Quail Feces
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rudenko, P.; Vatnikov, Y.; Sachivkina, N.; Rudenko, A.; Kulikov, E.; Lutsay, V.; Notina, E.; Bykova, I.; Petrov, A.; Drukovskiy, S.; et al. Search for Promising Strains of Probiotic Microbiota Isolated from Different Biotopes of Healthy Cats for Use in the Control of Surgical Infections. Pathogens 2021, 10, 667. [Google Scholar] [CrossRef] [PubMed]
- Hallen-Adams, H.E.; Suhr, M.J. Fungi in the healthy human gastrointestinal tract. Virulence 2017, 8, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Sachivkina, N.; Podoprigora, I.; Bokov, D. Morphological characteristics of Candida albicans, Candida krusei, Candida guilliermondii, and Candida glabrata biofilms, and response to Farnesol. Vet. World 2021, 14, 1608–1614. [Google Scholar] [CrossRef] [PubMed]
- Sachivkina, N.; Lenchenko, E.; Blumenkrants, D.; Ibragimova, A.; Bazarkina, O. Effects of farnesol and lyticase on the formation of Candida albicans biofilm. Vet. World 2020, 13, 1030–1036. [Google Scholar] [CrossRef]
- Vatnikov, Y.; Donnik, I.; Kulikov, E.; Karamyan, A.; Sachivkina, N.; Rudenko, P.; Tumanyan, A.; Khairova, N.; Romanova, E.; Gurina, R.; et al. Research on the antibacterial and antimycotic effect of the Phyto preparation Farnesol on biofilm-forming microorganisms in veterinary medicine. Int. J. Pharm. Res. 2020, 12, 1481–1492. [Google Scholar]
- Rudenko, P.; Sachivkina, N.; Vatnikov, Y.; Shabunin, S.; Engashev, S.; Kontsevaya, S.; Karamyan, A.; Bokov, D.; Kuznetsova, O.; Vasilieva, E. Role of microorganisms isolated from cows with mastitis in Moscow region in biofilm formation. Vet. World 2021, 14, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Lenchenko, E.; Blumenkrants, D.; Sachivkina, N.; Shadrova, N.; Ibragimova, A. Morphological and adhesive properties of Klebsiella pneumoniae biofilms. Vet. World 2020, 13, 197–200. [Google Scholar] [CrossRef] [Green Version]
- Sachivkina, N.P.; Lenchenko, E.M.; Marakhova, A.I. Study of the formation of Candida albicans and Escherichia coli biofilms. Farmatsiya 2019, 68, 26–30. [Google Scholar] [CrossRef]
- Jabra-Rizk, M.A.; Shirtliff, M.; James, C.; Meiller, T. Effect of farnesol on Candida dubliniensis biofilm formation and fluconazole resistance. FEMS Yeast Res. 2006, 6, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Langford, M.L.; Kenneth, S.H.; Nickerson, W.; Atkin, A.L. Activity and toxicity of farnesol towards Candida albicans are dependent on growth conditions. Antimicrob. Agents Chemother. 2010, 54, 940–942. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.; Henriques, M.; Azeredo, J.; Rocha, S.M.; Coimbra, M.A. Morphogenesis control in Candida albicans and Candida dubliniensis through signaling molecules produced by planktonic and biofilm cells. Eukaryot. Cell 2007, 12, 2429–2436. [Google Scholar] [CrossRef] [Green Version]
- Paguigan, N.D.; Rivera-Chávez, J.; Stempin, J.J.; Augustinović, M.; Noras, A.I.; Raja, H.A.; Todd, D.A.; Triplett, K.D.; Day, C.; Figueroa, M.; et al. Prenylated Diresorcinols Inhibit Bacterial Quorum Sensing. J. Nat. Prod. 2019, 82, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Hirooka, K.; Shioda, S.; Okada, M. Identification of critical residues for the catalytic activity of ComQ, a Bacillus prenylation enzyme for quorum sensing, by using a simple bioassay system. Biosci. Biotechnol. Biochem. 2020, 84, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, Y.G.; Khadke, S.K.; Yamano, A.; Woo, J.T.; Lee, J. Antimicrobial and antibiofilm activities of prenylated flavanones from Macaranga tanarius. Phytomedicine 2019, 63, 153033. [Google Scholar] [CrossRef] [PubMed]
- Marakasova, E.S.; Akhmatova, N.K.; Amaya, M.; Eisenhaber, B.; Eisenhaber, F.; van Hoek, M.L.; Baranova, A.V. Prenylation: From bacteria to eukaryotes. Mol. Biol. 2013, 47, 717–730. [Google Scholar] [CrossRef]
- Sachivkina, N.P.; Lenchenko, E.M.; Mannapova, R.T.; Strizhakov, A.A.; Romanova, E.V.; Lukina, D.M. Candida Biofilm Modeling: Past and Present. Farmatsiya 2019, 68, 18–22. [Google Scholar] [CrossRef]
- Sachivkina, N.P.; Kravtsov, E.G.; Vasilyeva, E.A.; Anokhina, I.V.; Dalin, M.V. Study of antimycotic activity of lyticase. Bull. Exp. Biol. Med. 2009, 148, 214–216. [Google Scholar] [CrossRef]
- Committee for the Update of the Guide for the Care and Use of Laboratory Animals of the Institute for Laboratory Animal Research, Division on Earth and Life Studies; National Research Council. Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, DC, USA, 2011. Available online: https://grants.nih.gov/grants/olaw/guide-for-the-care-anduse-of-laboratory-animals.pdf (accessed on 28 December 2020).
- Morozov, I.A.; Sachivkina, N.P.; Kravtsov, E.G.; Vasilyeva, E.A.; Anokhina, I.V.; Yashina, N.V.; Dalin, M.V. Damaging effects of lyticase on Candida albicans and changes in the response of rat alveolar macrophages to the contact with yeast-like fungi. Bull. Exp. Biol. Med. 2011, 151, 705–708. [Google Scholar] [CrossRef]
- Sachivkina, N.P.; Kravtsov, E.G.; Vasileva, E.A.; Anokchina, I.V.; Dalin, M.V. Efficiency of lyticase (bacterial enzyme) in experimental candidal vaginitis in mice. Bull. Exp. Biol. Med. 2010, 149, 727–730. [Google Scholar] [CrossRef]
- Sachivkina, N.; Lenchenko, E.; Strizakov, A.; Zimina, V.; Gnesdilova, L.; Gavrilov, V.; Byakhova, V.; Germanova, S.; Zharov, A.; Molchanova, M. The Evaluation of formation of biomembrane by microscopic Fungi of the Candida Genus. Int. J. Pharm. Res. 2018, 10, 738–744. [Google Scholar]
- Rossignol, T.; Logue, M.E.; Reynolds, K.; Grenon, M.; Lowndes, N.F.; Butler, G. Transcriptional response of Candida parapsilosis following exposure to Farnesol. Antimicrob. Agents Chemother. 2007, 51, 2304–2312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, T.; Miyazaki, T.; Sumiyoshi, M.; Ashizawa, N.; Takazono, T.; Yamamoto, K.; Imamura, Y.; Izumikawa, K.; Yanagihara, K.; Kohno, S.; et al. ERG3-Encoding Sterol C5,6- DESATURASE in Candida albicans Is Required for Virulence in an Enterically Infected Invasive Candidiasis Mouse Model. Pathogens 2021, 10, 23. [Google Scholar] [CrossRef]
- Sachivkina, N.P.; Karamyan, A.S.; Kuznetsova, O.M.; Byakhova, V.M.; Bondareva, I.B.; Molchanova, M.A. Development of therapeutic transdermal systems for microbial biofilm destruction. FEBS Open Bio 2019, 9 (Suppl. 1), 386. [Google Scholar]
- Ramage, G.; Saville, S.P.; Wickes, B.L.; Lopez-Ribot, J.L. Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl. Environ. Microbiol. 2002, 68, 5459–5463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenchenko, E.; Blumenkrants, D.; Vatnikov, Y.; Kulikov, E.; Khai, V.; Sachivkina, N.; Gnezdilova, L.; Sturov, N.; Sakhno, N.; Kuznetsov, V.; et al. Poultry Salmonella sensitivity to antibiotics. Sys. Rev. Pharm. 2020, 11, 170–175. [Google Scholar]
- Jacobsen, I.D.; Wilson, D.; Wachtler, B.; Brunke, S.L.; Naglik, J.R.; Hube, B. Candida albicans dimorphism as a therapeutic target. Expert Rev. Anti-Infect. Ther. 2012, 10, 85–93. [Google Scholar] [CrossRef] [Green Version]
- Witchley, J.N.; Penumetcha, P.; Abon, N.V.; Woolford, C.A.; Mitchell, A.P.; Noble, S.M. Candida albicans Morphogenesis Programs Control the Balance between Gut Commensalism and Invasive Infection. Cell Host Microbe. 2019, 25, 432–443. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; He, H.; Liu, J.; Xie, S.; Han, J. Inhibiting roles of farnesol and HOG in morphological switching of Candida albicans. Am. J. Transl. Res. 2020, 12, 6988–7001. [Google Scholar]
- Singkum, P.; Muangkaew, W.; Suwanmanee, S.; Pumeesat, P.; Wongsuk, T.; Luplertlop, N. Suppression of the pathogenicity of Candida albicans by the quorum-sensing molecules farnesol and tryptophol. J. Gen. Appl. Microbiol. 2020, 31, 277–283. [Google Scholar] [CrossRef] [Green Version]
- Zawrotniak, M.; Wojtalik, K.; Rapala-Kozik, M. Farnesol, a Quorum-Sensing Molecule of Candida Albicans Triggers the Release of Neutrophil Extracellular Traps. Cells 2019, 8, 1611. [Google Scholar] [CrossRef] [Green Version]
- Hasim, S.; Vaughn, E.N.; Donohoe, D.; Gordon, D.M.; Pfiffner, S.; Reynolds, T.B. Influence of phosphatidylserine and phosphatidylethanolamine on farnesol tolerance in Candida albicans. Yeast 2018, 35, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornby, J.M.; Kebaara, B.W.; Nickerson, K.W. Farnesol biosynthesis in Candida albicans: Cellular response to sterol inhibition by zaragozic acid B. Antimicrob. Agents Chemother. 2003, 47, 2366–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, R.A.; Monteiro, D.R.; Arias, L.S.; Fernandes, G.L.; Delbem, A.C.B.; Barbosa, D.B. Virulence Factors in Candida albicans and Streptococcus mutans Biofilms Mediated by Farnesol. Indian J. Microbiol. 2018, 58, 138–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Kim, Y.G.; Khadke, S.K.; Lee, J. Antibiofilm and antifungal activities of medium-chain fatty acids against Candida albicans via mimicking of the quorum-sensing molecule farnesol. Microb. Biotechnol. 2021, 14, 1353–1366. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Microorganism Culture | Optical Density (OD) | |
|---|---|---|
| Control | Experiment | |
| C. albicans | 0.497 ± 0.05 | 0.243 ± 0.06 |
| Indicators | ||||||||
|---|---|---|---|---|---|---|---|---|
| RBC, 1012 | HGB, g/L | LEU, 109 | PSEU, % | EOS, % | MONO, % | BAS, % | LYM, % | |
| 5 days experiment | 1.86 ± 0.13 | 85.4 ± 4.3 | 40.5 ± 1.9 | 22.2 ± 1.8 | 3.3 ± 0.4 | 3.4 ± 0.6 | 0.30 ± 0.05 | 71.5 ± 6.4 |
| Control 5 days | 1.90 ± 0.11 | 86.2 ± 3.9 | 42.1 ± 1.4 | 28.1 ± 2.4 | 3.2 ± 0.6 | 2.9 ± 0.8 | 0.42 ± 0.06 | 73.4 ± 4.9 |
| 10 days experiment | 1.72 ± 0.12 | 75.4 ± 6.3 | 30.2 ± 0.9 | 24.7 ± 2.6 | 3.0 ± 0.4 | 2.4 ± 0.7 | 0.36 ± 0.08 | 79.0 ± 6.5 |
| Control 10 days | 1.84 ± 0.11 | 82.7 ± 5.8 | 38.5 ± 1.7 | 20.0 ± 1.9 | 3.5 ± 0.7 | 2.5 ± 0.7 | 0.38 ± 0.11 | 68.4 ± 7.1 |
| 15 days experiment | 1.69 ± 0.15 | 72.9 ± 5.1 | 34.5 ± 2.1 | 26.5 ± 2.8 | 2.8 ± 0.8 | 2.0 ± 0.6 | 0.36 ± 0.05 | 77.6 ± 5.4 |
| Control 15 days | 1.87 ± 0.13 | 83.1 ± 3.4 | 35.5 ± 1.6 | 18.6 ± 1.7 | 3.6 ± 0.5 | 2.1 ± 0.3 | 0.35 ± 0.09 | 70.4 ± 6.1 |
| Indicators | ||||||||
|---|---|---|---|---|---|---|---|---|
| CRE mmol/L | AST ME/L | ALT ME/L | ALP ME/L | GLU mmol/L | CH mmol/L | TG g/L | TBIL mmol/L | |
| 5 days experiment | 20.6 ± 0.5 | 2.1 ± 0.3 | 0.11 ± 0.02 | 3.7 ± 0.3 | 13.7 ± 0.4 | 4.3 ± 0.6 | 1.11 ± 0.14 | 1.6 ± 0.4 |
| Control 5 days | 20.4 ± 0.4 | 2.3 ± 0.3 | 0.07 ± 0.02 | 3.5 ± 0.4 | 14.2 ± 0.5 | 3.7 ± 0.4 | 0.99 ± 0.10 | 1.8 ± 0.4 |
| 10 days experiment | 23.1 ± 0.3 | 2.7 ± 0.4 | 0.17 ± 0.03 | 5.3 ± 0.2 | 11.1 ± 0.4 | 2.8 ± 0.4 | 0.96 ± 0.12 | 2.3 ± 0.3 |
| Control 10 days | 18.8 ± 0.4 | 1.7 ± 0.3 | 0.11 ± 0.02 | 4.2 ± 0.3 | 12.3 ± 0.4 | 2.6 ± 0.4 | 1.2 ± 0.11 | 2.9 ± 0.6 |
| 15 days experiment | 25.9 ± 0.3 | 2.5 ± 0.4 | 0.18 ± 0.02 | 5.9 ± 0.4 | 12.3 ± 0.4 | 2.5 ± 0.4 | 1.2 ± 0.11 | 2.5 ± 0.5 |
| Control 15 days | 17.9 ± 0.4 | 1.8 ± 0.3 | 0.09 ± 0.02 | 4.2 ± 0.3 | 12.6 ± 0.4 | 2.6 ± 0.4 | 1.2 ± 0.13 | 2.9 ± 0.5 |
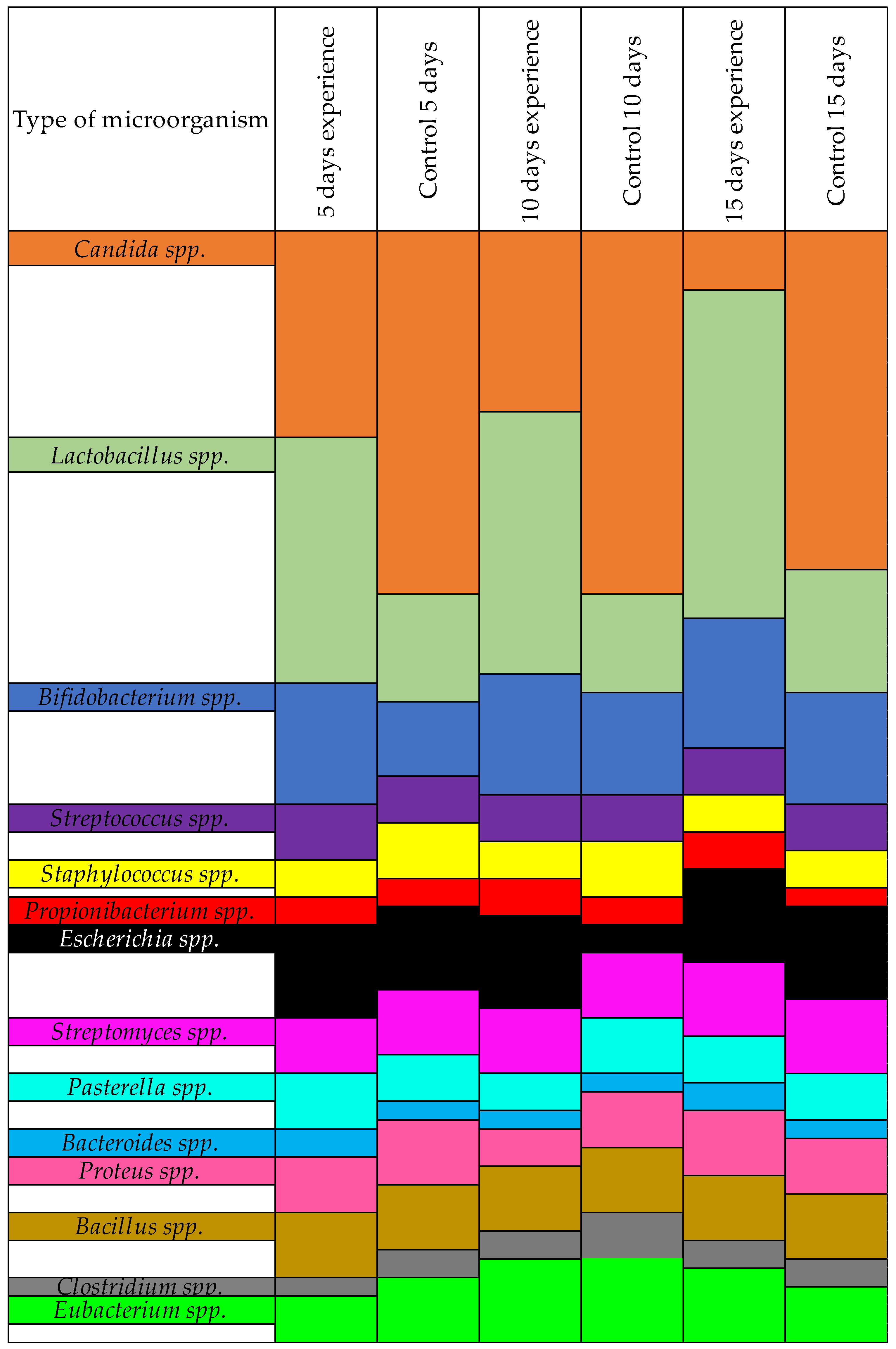
| Type of Microorganism | Number of Isolates | |||||
|---|---|---|---|---|---|---|
| 5 Days Experiment | Control 5 Days | 10 Days Experiment | Control 10 Days | 15 Days Experiment | Control 15 Days | |
| Candida spp. | 4.13 ± 0.21 | 6.12 ± 0.36 | 3.13 ± 0.24 | 6.27 ± 0.27 | 1.13 ± 0.16 | 5.88 ± 0.39 |
| Lactobacillus spp. | 6.28 ± 0.35 | 4.13 ± 0.25 | 6.43 ± 0.51 | 4.01 ± 0.32 | 5.89 ± 0.27 | 4.34 ± 0.31 |
| Bifidobacterium spp. | 5.67 ± 0.28 | 3.42 ± 0.36 | 5.51 ± 0.29 | 4.59 ± 0.41 | 5.81 ± 0.38 | 5.02 ± 0.25 |
| Streptococcus spp. | 2.61 ± 0.48 | 2.34 ± 0.40 | 2.27 ± 0.36 | 2.11 ± 0.40 | 2.33 ± 0.29 | 2.08 ± 0.35 |
| Staphylococcus spp. | 1.91 ± 0.35 | 2.47 ± 0.21 | 1.87 ± 0.31 | 2.70 ± 0.33 | 1.90 ± 0.37 | 1.71 ± 0.29 |
| Propionibacterium spp. | 1.65 ± 0.19 | 1.50 ± 0.11 | 2.10 ± 0.28 | 1.63 ± 0.22 | 1.73 ± 0.25 | 1.80 ± 0.19 |
| Escherichia spp. | 4.31 ± 0.22 | 3.97 ± 0.35 | 4.24 ± 0.40 | 4.00 ± 0.29 | 4.33 ± 0.32 | 4.37 ± 0.25 |
| Streptomyces spp. | 2.84 ± 0.23 | 3.14 ± 0.31 | 3.02 ± 0.19 | 2.94 ± 0.26 | 3.18 ± 0.20 | 3.41 ± 0.23 |
| Pasterella spp. | 2.59 ± 0.32 | 2.31 ± 0.44 | 2.63 ± 0.27 | 2.58 ± 0.39 | 2.11 ± 0.36 | 2.37 ± 0.40 |
| Bacteroides spp. | 1.47 ± 0.25 | 0.91 ± 0.30 | 1.23 ± 0.24 | 1.07 ± 0.19 | 1.25 ± 0.25 | 1.16 ± 0.22 |
| Proteus spp. | 2.54 ± 0.39 | 3.05 ± 0.24 | 2.78 ± 0.31 | 2.60 ± 0.27 | 3.10 ± 0.24 | 2.90 ± 0.29 |
| Bacillus spp. | 2.97 ± 0.13 | 2.97 ± 0.13 | 2.97 ± 0.13 | 2.97 ± 0.13 | 2.97 ± 0.13 | 2.97 ± 0.13 |
| Clostridium spp. | 0.93 ± 0.16 | 1.21 ± 0.20 | 1.33 ± 0.25 | 1.34 ± 0.28 | 1.51 ± 0.26 | 1.60 ± 0.21 |
| Eubacterium spp. | 2.68 ± 0.22 | 2.66 ± 0.24 | 2.74 ± 0.21 | 2.51 ± 0.30 | 2.70 ± 0.31 | 2.49 ± 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sachivkina, N.; Vasilieva, E.; Lenchenko, E.; Kuznetsova, O.; Karamyan, A.; Ibragimova, A.; Zhabo, N.; Molchanova, M. Reduction in Pathogenicity in Yeast-like Fungi by Farnesol in Quail Model. Animals 2022, 12, 489. https://doi.org/10.3390/ani12040489
Sachivkina N, Vasilieva E, Lenchenko E, Kuznetsova O, Karamyan A, Ibragimova A, Zhabo N, Molchanova M. Reduction in Pathogenicity in Yeast-like Fungi by Farnesol in Quail Model. Animals. 2022; 12(4):489. https://doi.org/10.3390/ani12040489
Chicago/Turabian StyleSachivkina, Nadezhda, Elena Vasilieva, Ekaterina Lenchenko, Olga Kuznetsova, Arfenia Karamyan, Alfia Ibragimova, Natalia Zhabo, and Maria Molchanova. 2022. "Reduction in Pathogenicity in Yeast-like Fungi by Farnesol in Quail Model" Animals 12, no. 4: 489. https://doi.org/10.3390/ani12040489
APA StyleSachivkina, N., Vasilieva, E., Lenchenko, E., Kuznetsova, O., Karamyan, A., Ibragimova, A., Zhabo, N., & Molchanova, M. (2022). Reduction in Pathogenicity in Yeast-like Fungi by Farnesol in Quail Model. Animals, 12(4), 489. https://doi.org/10.3390/ani12040489