
Effects of Environmental Heat Load on Endocannabinoid System Components in Adipose Tissue of High Yielding Dairy Cows

 ,
, 

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Procedures and Collection of Blood and Milk
2.1.1. Seasonal Effects on eCB ‘Tone’ in Adipose Tissue of Early Postpartum Cows (S vs. W)
2.1.2. Comparing Not Cooled (Heat-Stressed) vs. Cooled Late-Lactation Cows in Summer (HS vs. CL)
2.2. Analysis of Circulating NEFA, Oxidative Stress and Inflammatory Markers
2.3. Biopsy of Subcutaneous AT
2.4. Measurements of eCBs in AT
2.5. Relative Gene Expression in AT by Quantitative Real-Time PCR
2.6. Protein Abundance in AT by Immunoblot Analysis
2.7. Homology Modeling and Active Site Prediction of CB1 and CB2
Molecular Interaction of 2-AG and AEA with CB1 and CB2 Receptors
2.8. Statistical Analysis
3. Results
3.1. Heat Load Affects Plasma Indicators of Metabolism, Inflammation, Oxidative Stress, and Lactation Performance in Periparturient Cows (S vs. W)
3.2. Heat Load Impacts Transcription of ECS Components in AT of S vs. W Calving Cows
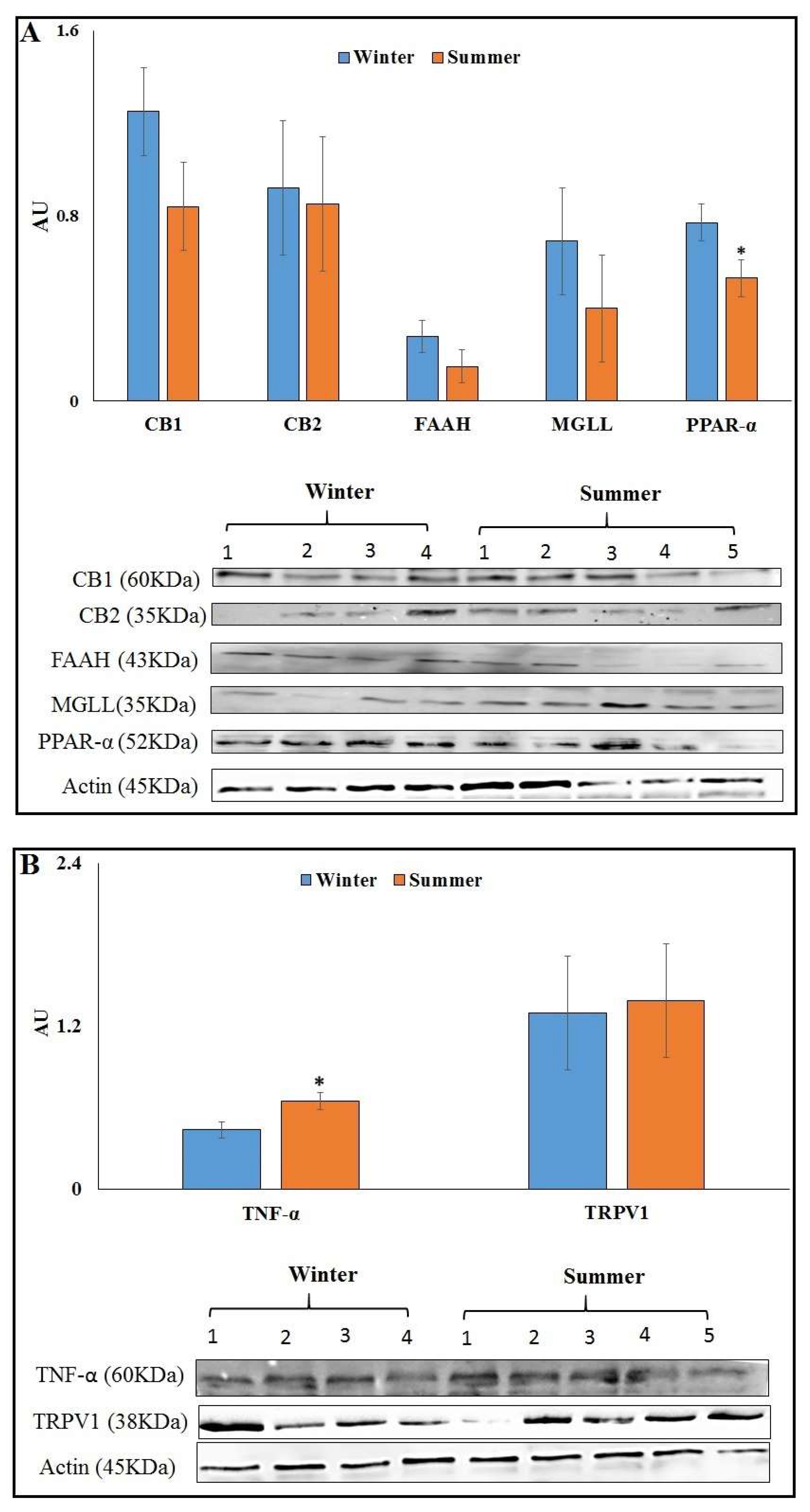
3.3. Heat Load and ECS Components’ Protein Abundance in AT of S vs. W Calving Cows
3.4. Levels of eCBs in AT and Plasma at 7 d PP of Summer vs. Winter
3.5. Abating Heat Load by Cooling Affects Idices of Heat Stress in Late-Lactation Cows (HS vs. CL)
3.6. Minimizing Heat Load by Cooling Did Not Affect Gene Expression of ECS Components in AT of HS vs. CL
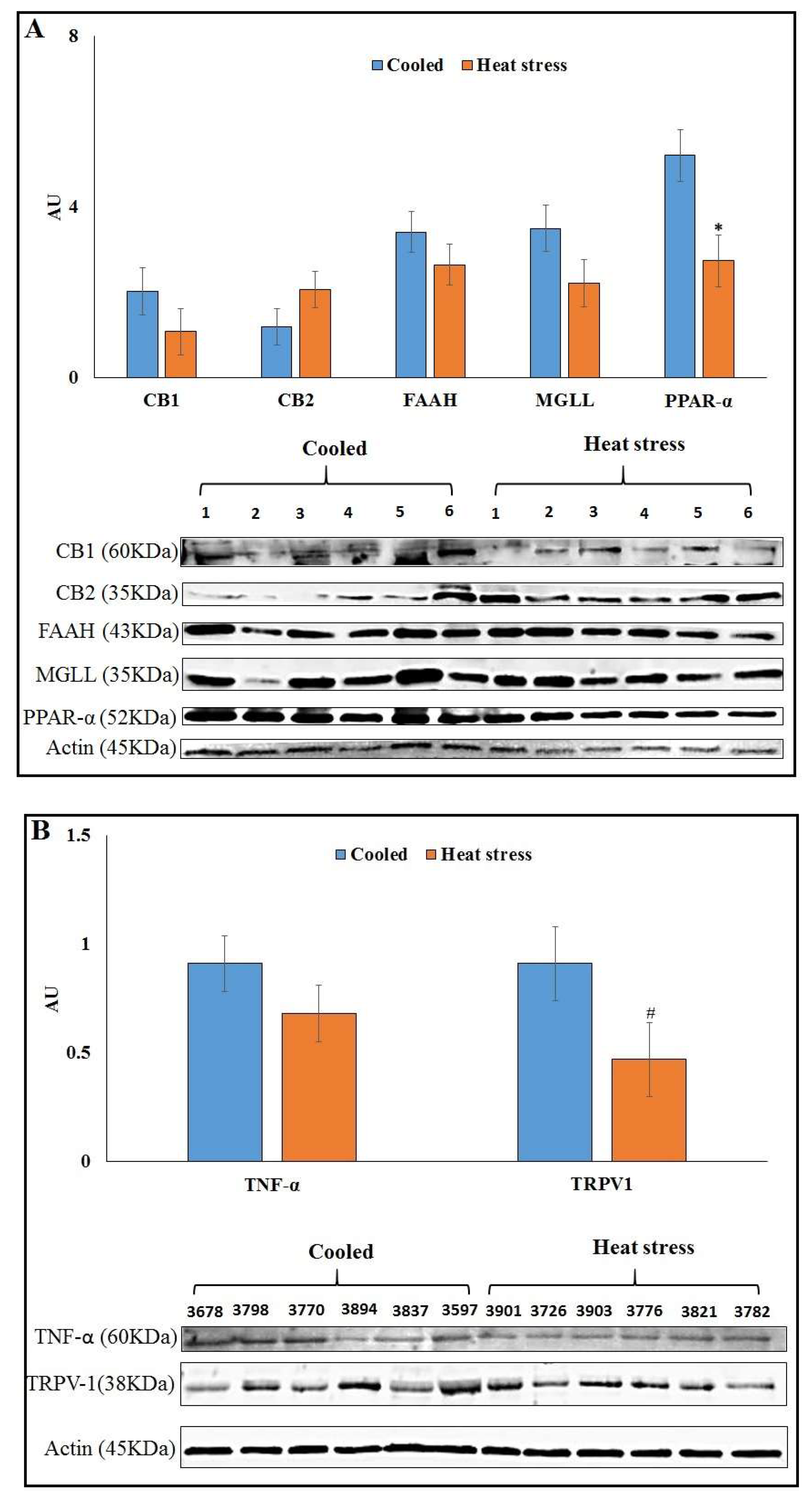
3.7. Minimizing Heat Load by Cooling Affects Protein Abundance of ECS Components in AT of HS vs. CL
3.8. Levels of eCBs in AT, Plasma and Milk of HS vs. Cooled Cows
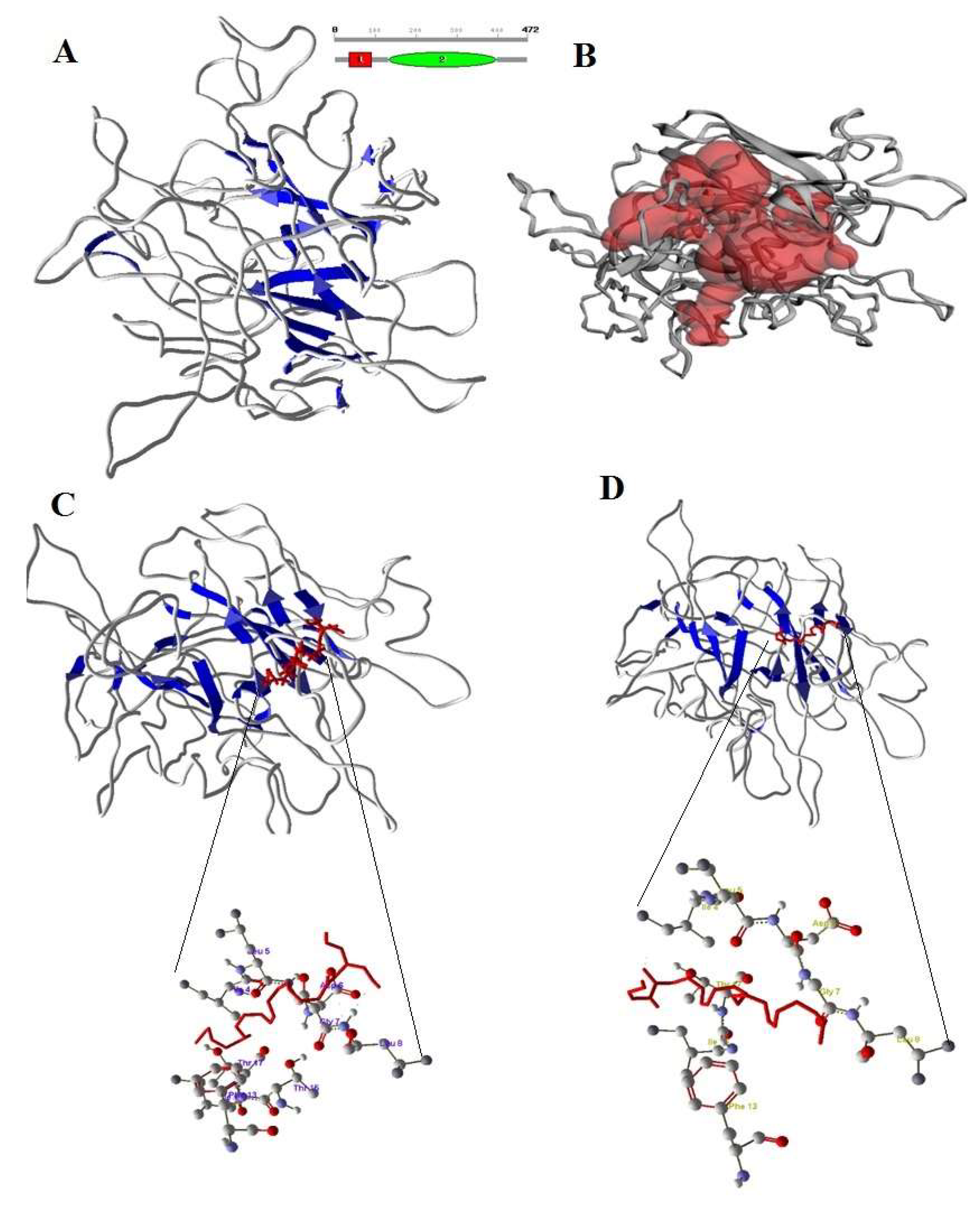
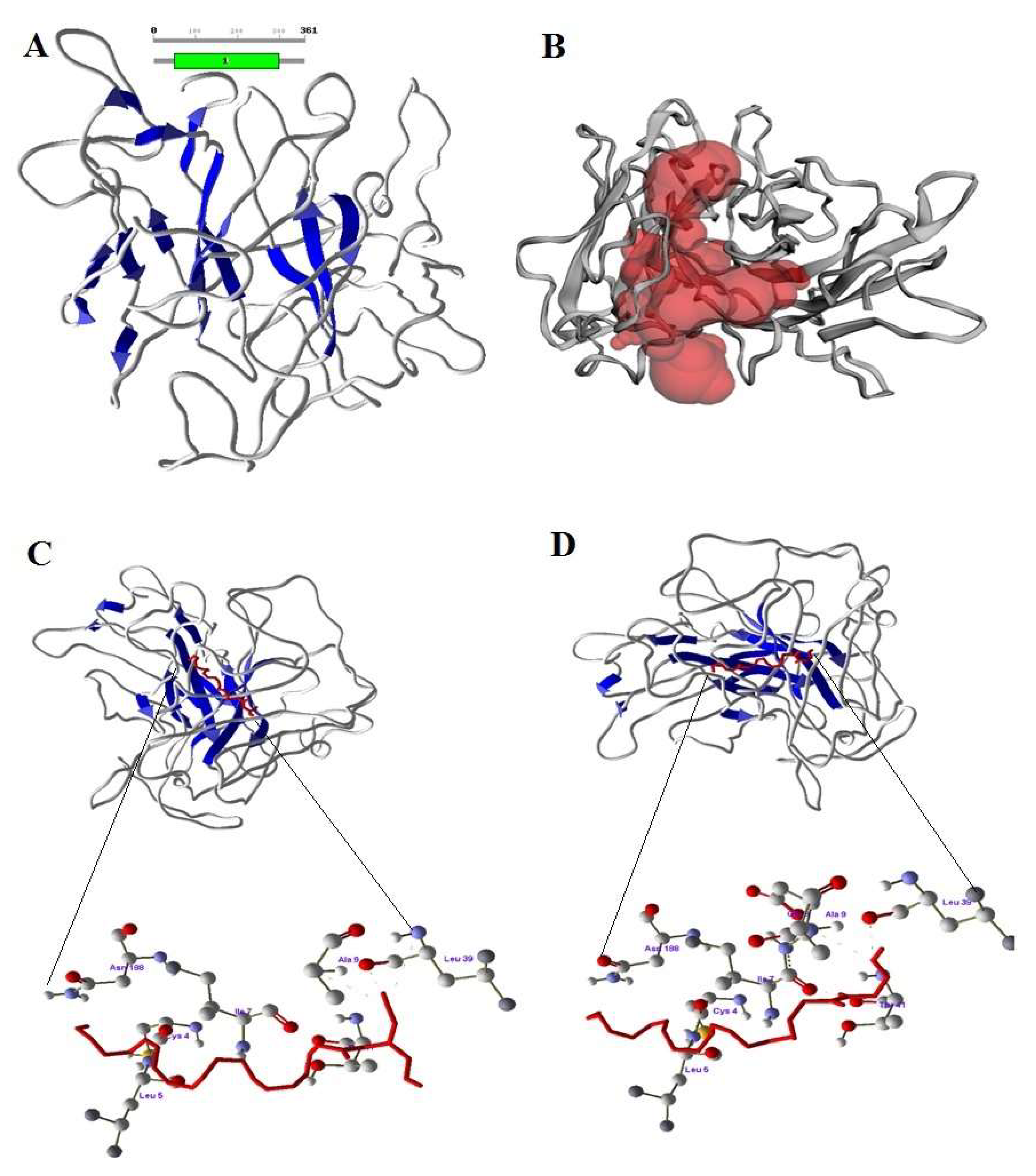
3.9. Modeling the Bovine CB1 and CB2 Receptors and Interactions of 2-AG and AEA
4. Discussion
4.1. Effects of HL on Feed Intake, Metabolic Response, Oxidative Stress and Inflammation and the Possible Role of the ECS in These Responses
4.2. Effects of HS on the ECS Receptors, MGLL and PPAR-α in AT
4.3. eCB Levels in Plasma, Milk and AT of PP Dairy Cows
4.4. Effects of HL on TRPV1 in AT
4.5. Molecular Interaction of Bovine CB1, CB2 Receptors with eCBs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Solomon, S.; Manning, M.; Marquis, M.; Qin, D. Climate Change 2007-the Physical Science Basis: Working Group I Contribution to the Fourth Assessment Report of the IPCC; Cambridge University Press: Cambridge, UK, 2007; Volume 4, ISBN 0521705967. [Google Scholar]
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic Losses from Heat Stress by US Livestock Industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef] [Green Version]
- Bernabucci, U.; Basiricò, L.; Morera, P.; Dipasquale, D.; Vitali, A.; Piccioli Cappelli, F.; Calamari, L. Effect of summer season on milk protein fractions in Holstein cows. J. Dairy Sci. 2015, 98, 1815–1827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, S.; Orellana Rivas, R.M.; Marins, T.N.; Chen, Y.-C.; Gao, J.; Bernard, J.K. Impact of heat stress on lactational performance of dairy cows. Theriogenology 2020, 150, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.; Dahl, G.E. Invited review: Heat stress effects during late gestation on dry cows and their calves. J. Dairy Sci. 2013, 96, 4079–4093. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.J.; Weng, X.G.; Wang, J.F.; Zhou, D.; Zhang, W.; Zhai, C.C.; Hou, Y.X.; Zhu, Y.H. Effects of temperature-humidity index and chromium supplementation on antioxidant capacity, heat shock protein 72, and cytokine responses of lactating cows. J. Anim. Sci. 2014, 92, 3026–3034. [Google Scholar] [CrossRef] [PubMed]
- do Amaral, B.C.; Connor, E.E.; Tao, S.; Hayen, M.J.; Bubolz, J.W.; Dahl, G.E. Heat stress abatement during the dry period influences metabolic gene expression and improves immune status in the transition period of dairy cows. J. Dairy Sci. 2011, 94, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Zachut, M.; Kra, G.; Livshitz, L.; Portnick, Y.; Yakoby, S.; Friedlander, G.; Levin, Y. Seasonal heat stress affects adipose tissue proteome toward enrichment of the Nrf2-mediated oxidative stress response in late-pregnant dairy cows. J. Proteom. 2017, 158, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Myers, M.N.; Zachut, M.; Tam, J.; Contreras, G.A. A proposed modulatory role of the endocannabinoid system on adipose tissue metabolism and appetite in periparturient dairy cows. J. Anim. Sci. Biotechnol. 2021, 12, 21. [Google Scholar] [CrossRef]
- Silvestri, C.; Di Marzo, V. The Endocannabinoid System in Energy Homeostasis and the Etiopathology of Metabolic Disorders. Cell Metab. 2013, 17, 475–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachut, M.; Kra, G.; Moallem, U.; Livshitz, L.; Levin, Y.; Udi, S.; Nemirovski, A.; Tam, J. Characterization of the endocannabinoid system in subcutaneous adipose tissue in periparturient dairy cows and its association to metabolic profiles. PLoS ONE 2018, 13, e0205996. [Google Scholar] [CrossRef] [PubMed]
- Kuhla, B.; Kaever, V.; Tuchscherer, A.; Kuhla, A. Involvement of Plasma Endocannabinoids and the Hypothalamic Endocannabinoid System in Increasing Feed Intake after Parturition of Dairy Cows. Neuroendocrinology 2020, 110, 246–257. [Google Scholar] [CrossRef]
- Bensaid, M.; Gary-Bobo, M.; Esclangon, A.; Maffrand, J.P.; Le Fur, G.; Oury-Donat, F.; Soubrié, P. The cannabinoid CB1 receptor antagonist SR141716 increases Acrp30 mRNA expression in adipose tissue of obese fa/fa rats and in cultured adipocyte cells. Mol. Pharmacol. 2003, 63, 908–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cota, D.; Marsicano, G.; Tschöp, M.; Grübler, Y.; Flachskamm, C.; Schubert, M.; Auer, D.; Yassouridis, A.; Thöne-Reineke, C.; Ortmann, S.; et al. The endogenous cennabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis. J. Clin. Investig. 2003, 112, 423–431. [Google Scholar] [CrossRef]
- Matias, I.; Gonthier, M.-P.; Orlando, P.; Martiadis, V.; De Petrocellis, L.; Cervino, C.; Petrosino, S.; Hoareau, L.; Festy, F.; Pasquali, R.; et al. Regulation, Function, and Dysregulation of Endocannabinoids in Models of Adipose and β-Pancreatic Cells and in Obesity and Hyperglycemia. J. Clin. Endocrinol. Metab. 2006, 91, 3171–3180. [Google Scholar] [CrossRef]
- Blüher, M.; Engeli, S.; Klöting, N.; Berndt, J.; Fasshauer, M.; Bátkai, S.; Pacher, P.; Schön, M.R.; Jordan, J.; Stumvoll, M. Dysregulation of the peripheral and adipose tissue endocannabinoid system in human abdominal obesity. Diabetes 2006, 55, 3053–3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devane, W.A.; Hanus, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Ben-Shabat, S.; Hanus, L.; Ligumsky, M.; Kaminski, N.E.; Schatz, A.R.; Gopher, A.; Almog, S.; Martin, B.R.; Compton, D.R. Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem. Pharmacol. 1995, 50, 83–90. [Google Scholar] [CrossRef]
- Sugiura, T.; Kondo, S.; Sukagawa, A.; Nakane, S.; Shinoda, A.; Itoh, K.; Yamashita, A.; Waku, K. 2-Arachidonoylglycerol: A possible endogenous cannabinoid receptor ligand in brain. Biochem. Biophys. Res. Commun. 1995, 215, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Silver, R.J. The Endocannabinoid System of Animals. Animals 2019, 9, 686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, S.; Dittel, B.N. Unraveling the complexities of cannabinoid receptor 2 (CB2) immune regulation in health and disease. Immunol. Res. 2011, 51, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Dirandeh, E.; Ghorbanalinia, M.; Rezaei-Roodbari, A.; Colazo, M.G. Relationship between body condition score loss and mRNA of genes related to fatty acid metabolism and the endocannabinoid system in adipose tissue of periparturient cows. Animal 2020, 14, 1724–1732. [Google Scholar] [CrossRef] [PubMed]
- Izzo, A.A.; Piscitelli, F.; Capasso, R.; Marini, P.; Cristino, L.; Petrosino, S.; Di Marzo, V. Basal and fasting/refeeding-regulated tissue levels of endogenous PPAR-alpha ligands in Zucker rats. Obesity 2010, 18, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R.; Tamir, R.; Klionsky, L.; Norman, M.H.; Louis, J.-C.; Wild, K.D.; Treanor, J.J.S. Proton Activation Does Not Alter Antagonist Interaction with the Capsaicin-Binding Pocket of TRPV1. Mol. Pharmacol. 2005, 68, 1524–1533. [Google Scholar] [CrossRef] [Green Version]
- Christie, S.; Wittert, G.A.; Li, H.; Page, A.J. Involvement of TRPV1 Channels in Energy Homeostasis. Front. Endocrinol. 2018, 9, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szallasi, A.; Cortright, D.N.; Blum, C.A.; Eid, S.R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. Nat. Rev. Drug Discov. 2007, 6, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Ho, K.W.; Ward, N.J.; Calkins, D.J. TRPV1: A stress response protein in the central nervous system. Am. J. Neurodegener. Dis. 2012, 1, 1–14. [Google Scholar] [PubMed]
- Zachut, M.; Kra, G.; Nemes-Navon, N.; Ben-Aharon, N.; Moallem, U.; Lavon, Y.; Jacoby, S. Seasonal heat load is more potent than the degree of body weight loss in dysregulating immune function by reducing white blood cell populations and increasing inflammation in Holstein dairy cows. J. Dairy Sci. 2020, 103, 10809–10822. [Google Scholar] [CrossRef]
- Bohmanova, J.; Misztal, I.; Cole, J.B. Temperature-Humidity Indices as Indicators of Milk Production Losses due to Heat Stress. J. Dairy Sci. 2007, 90, 1947–1956. [Google Scholar] [CrossRef] [PubMed]
- Moallem, U.; Kamer, H.; Hod, A.; Lifshitz, L.; Kra, G.; Jacoby, S.; Portnick, Y.; Zachut, M. Reducing milking frequency from thrice to twice daily in early lactation improves the metabolic status of high-yielding dairy cows with only minor effects on yields. J. Dairy Sci. 2019, 102, 9468–9480. [Google Scholar] [CrossRef] [PubMed]
- Council, N.R. Nutrient Requirements of Dairy Cattle, 7th ed.; The National Academies Press: Washington, DC, USA, 2001; pp. 18–19. ISBN 978-0-309-06997-7. [Google Scholar]
- Edmonoson, A.J.; Lean, I.J.; Weaker, L.D.; Farver, T.; Webster, G. A Body Condition Scoring Chart for Holstein Dairy Cows. J. Dairy Sci. 1989, 72, 68–78. [Google Scholar] [CrossRef]
- Feldman, E. Thiobarbituric acid reactive substances (TBARS) Assay. Anim. Models Diabet. Complicat. Consort. 2004, 1, 1–3. [Google Scholar]
- Zachut, M.; Honig, H.; Striem, S.; Zick, Y.; Boura-Halfon, S.; Moallem, U. Periparturient dairy cows do not exhibit hepatic insulin resistance, yet adipose-specific insulin resistance occurs in cows prone to high weight loss. J. Dairy Sci. 2013, 96, 5656–5669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aran, A.; Cassuto, H.; Lubotzky, A.; Wattad, N.; Hazan, E. Brief Report: Cannabidiol-Rich Cannabis in Children with Autism Spectrum Disorder and Severe Behavioral Problems—A Retrospective Feasibility Study. J. Autism Dev. Disord. 2019, 49, 1284–1288. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daddam, J.R.; Dowlathabad, M.R.; Panthangi, S.; Jasti, P. Molecular docking and P-glycoprotein inhibitory activity of Flavonoids. Interdiscip. Sci. Comput. Life Sci. 2014, 6, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Thepbandit, W.; Papathoti, N.K.; Daddam, J.R.; Thumanu, K.; Siriwong, S.; Le Thanh, T.; Buensanteai, N. Identification of Salicylic Acid Mechanism against Leaf Blight Disease in Oryza sativa by SR-FTIR Microspectroscopic and Docking Studies. Pathogens 2021, 10, 652. [Google Scholar] [CrossRef]
- Daddam, J.R.; Sreenivasulu, B.; Umamahesh, K.; Peddanna, K.; Rao, D.M. In Silico Studies on Anti-Stress Compounds of Ethanolic Root Extract of Hemidesmus indicus L. Curr. Pharm. Biotechnol. 2019, 21, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Rayalu, D.J.; Selvaraj, C.; Singh, S.K.; Ganeshan, R.; Udaya Kumar, N.; Seshapani, P. Homology modeling, active site prediction, and targeting the anti hypertension activity through molecular docking on endothelin–B receptor domain. Bioinformation 2012, 8, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, B.C.K.; Kuo, W.W.; Day, C.H.; Hsieh, D.J.Y.; Kuo, C.H.; Daddam, J.; Chen, R.J.; Padma, V.V.; Wang, G.; Huang, C.Y. The soybean bioactive peptide VHVV alleviates hypertension-induced renal damage in hypertensive rats via the SIRT1-PGC1α/Nrf2 pathway. J. Funct. Foods 2020, 75, 104255. [Google Scholar] [CrossRef]
- Kurjogi, M.; Satapute, P.; Jogaiah, S.; Abdelrahman, M.; Daddam, J.R.; Ramu, V.; Tran, L.S.P. Computational modeling of the staphylococcal enterotoxins and its interaction with natural antitoxin compounds. Int. J. Mol. Sci. 2018, 19, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, H.; Wang, W.; Guo, L.; Bindelle, J. Recent Advances on Early Detection of Heat Strain in Dairy Cows Using Animal-Based Indicators: A Review. Animals 2021, 11, 980. [Google Scholar] [CrossRef] [PubMed]
- Shwartz, G.; Rhoads, M.L.; VanBaale, M.J.; Rhoads, R.P.; Baumgard, L.H. Effects of a supplemental yeast culture on heat-stressed lactating Holstein cows1. J. Dairy Sci. 2009, 92, 935–942. [Google Scholar] [CrossRef] [Green Version]
- Tam, J.; Vemuri, V.K.; Liu, J.; Bátkai, S.; Mukhopadhyay, B.; Godlewski, G.; Osei-Hyiaman, D.; Ohnuma, S.; Ambudkar, S.V.; Pickel, J.; et al. Peripheral CB1 cannabinoid receptor blockade improves cardiometabolic risk in mouse models of obesity. J. Clin. Investig. 2010, 120, 2953–2966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, J.; Cinar, R.; Liu, J.; Godlewski, G.; Wesley, D.; Jourdan, T.; Szanda, G.; Mukhopadhyay, B.; Chedester, L.; Liow, J.-S.; et al. Peripheral cannabinoid-1 receptor inverse agonism reduces obesity by reversing leptin resistance. Cell Metab. 2012, 16, 167–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jastrebski, S.F.; Lamont, S.J.; Schmidt, C.J. Chicken hepatic response to chronic heat stress using integrated transcriptome and metabolome analysis. PLoS ONE 2017, 12, e0181900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz de Azua, I.; Mancini, G.; Srivastava, R.K.; Rey, A.A.; Cardinal, P.; Tedesco, L.; Zingaretti, C.M.; Sassmann, A.; Quarta, C.; Schwitter, C.; et al. Adipocyte cannabinoid receptor CB1 regulates energy homeostasis and alternatively activated macrophages. J. Clin. Investig. 2017, 127, 4148–4162. [Google Scholar] [CrossRef] [PubMed]
- Joyeux, M.; Arnaud, C.; Godin-Ribuot, D.; Demenge, P.; Lamontagne, D.; Ribuot, C. Endocannabinoids are implicated in the infarct size-reducing effect conferred by heat stress preconditioning in isolated rat hearts. Cardiovasc. Res. 2002, 55, 619–625. [Google Scholar] [CrossRef] [Green Version]
- Morena, M.; Patel, S.; Bains, J.S.; Hill, M.N. Neurobiological Interactions Between Stress and the Endocannabinoid System. Neuropsychopharmacology 2016, 41, 80–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drori, A.; Gammal, A.; Azar, S.; Hinden, L.; Hadar, R.; Wesley, D.; Nemirovski, A.; Szanda, G.; Salton, M.; Tirosh, B.; et al. CB1R regulates soluble leptin receptor levels via CHOP, contributing to hepatic leptin resistance. eLife 2020, 9, e60771. [Google Scholar] [CrossRef]
- Karatsoreos, I.N.; McEwen, B.S. Psychobiological allostasis: Resistance, resilience and vulnerability. Trends Cogn. Sci. 2011, 15, 576–584. [Google Scholar] [CrossRef]
- Bonsale, R.; Seyed Sharifi, R.; Dirandeh, E.; Hedayat, N.; Mojtahedin, A.; Ghorbanalinia, M.; Abolghasemi, A. Endocannabinoids as endometrial inflammatory markers in lactating Holstein cows. Reprod. Domest. Anim. 2018, 53, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Sumislawski, J.J.; Ramikie, T.S.; Patel, S. Reversible Gating of Endocannabinoid Plasticity in the Amygdala by Chronic Stress: A Potential Role for Monoacylglycerol Lipase Inhibition in the Prevention of Stress-Induced Behavioral Adaptation. Neuropsychopharmacology 2011, 36, 2750–2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouveia-Figueira, S.; Nording, M.L. Development and validation of a sensitive UPLC-ESI-MS/MS method for the simultaneous quantification of 15 endocannabinoids and related compounds in milk and other biofluids. Anal. Chem. 2014, 86, 1186–1195. [Google Scholar] [CrossRef] [PubMed]
- Gouveia-Figueira, S.; Nording, M.L. Validation of a tandem mass spectrometry method using combined extraction of 37 oxylipins and 14 endocannabinoid-related compounds including prostamides from biological matrices. Prostaglandins Other Lipid Mediat. 2015, 121, 110–121. [Google Scholar] [CrossRef]
- Muller, C.; Lynch, D.L.; Hurst, D.P.; Reggio, P.H. TRPV1 Activation by Anandamide via a Unique Lipid Pathway. J. Chem. Inf. Model. 2021, 61, 5742–5746. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, X.; Nie, Y.; Tian, Y.; Xiao, X.; Yang, F. Endocannabinoid activation of the TRPV1 ion channel is distinct from activation by capsaicin. J. Biol. Chem. 2021, 297, 101022. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.K.; Pakkkianathan, B.C.; Kumar, M.; Daddam, J.R.; Jayavel, S.; Kannan, M.; Pillai, G.G.; Krishnan, M. Computational studies on molecular interactions of 6-thioguanosine analogs with anthrax toxin receptor 1. Interdiscip. Sci. Comput. Life Sci. 2012, 4, 183–189. [Google Scholar] [CrossRef]
- Lin, Y.-M.; Badrealam, K.F.; Kuo, C.-H.; Daddam, J.; Shibu, M.A.; Lin, K.-H.; Ho, T.-J.; Viswanadha, V.P.; Kuo, W.-W.; Huang, C.-Y. Small Molecule Compound Nerolidol attenuates Hypertension induced hypertrophy in spontaneously hypertensive rats through modulation of Mel-18-IGF-IIR signalling. Phytomedicine 2021, 84, 153450. [Google Scholar] [CrossRef] [PubMed]
- Desai, T.D.; Wen, Y.-T.; Daddam, J.R.; Cheng, F.; Chen, C.-C.; Pan, C.-L.; Lin, K.-L.; Tsai, R.-K. Long term therapeutic effects of icariin-loaded PLGA microspheres in an experimental model of optic nerve ischemia via modulation of CEBP-β/G-CSF/noncanonical NF-κB axis. Bioeng. Transl. Med. 2021, e10289. [Google Scholar] [CrossRef]
- Kra, G.; Daddam, J.R.; Gabay, H.; Yosefi, S.; Zachut, M. Antioxidant Resveratrol Increases Lipolytic and Reduces Lipogenic Gene Expression under In Vitro Heat Stress Conditions in Dedifferentiated Adipocyte-Derived Progeny Cells from Dairy Cows. Antioxidants 2021, 10, 905. [Google Scholar] [CrossRef] [PubMed]
- Shaik, A.H.; Shaik, S.R.; Daddam, J.R.; Ali, D.; Manoharadas, S.; Arafah, M.W.; Kodidhela, L.D. Maslinic acid and gallic acid protective efficacy on lipids, lipoproteins and lipid metabolizing enzymes against isoproterenol administered cardiotoxicity: An in vivo and in silico molecular docking evidences. J. King Saud Univ.-Sci. 2021, 33, 101230. [Google Scholar] [CrossRef]
- Ali, A.; Kuo, W.W.; Kuo, C.H.; Lo, J.F.; Chen, M.Y.C.; Daddam, J.R.; Ho, T.J.; Viswanadha, V.P.; Shibu, M.A.; Huang, C.Y. E3 ligase activity of Carboxyl terminus of Hsc70 interacting protein (CHIP) in Wharton’s jelly derived mesenchymal stem cells improves their persistence under hyperglycemic stress and promotes the prophylactic effects against diabetic cardiac damages. Bioeng. Transl. Med. 2021, 6, e10234. [Google Scholar] [CrossRef] [PubMed]
- Daddam, J.R.; Sreenivasulu, B.; Peddanna, K.; Umamahesh, K. Designing, docking and molecular dynamics simulation studies of novel cloperastine analogues as anti-allergic agents: Homology modeling and active site prediction for the human histamine H1 receptor. RSC Adv. 2020, 10, 4745–4754. [Google Scholar] [CrossRef] [Green Version]
- Saleh, N.; Hucke, O.; Kramer, G.; Schmidt, E.; Montel, F.; Lipinski, R.; Ferger, B.; Clark, T.; Hildebrand, P.W.; Tautermann, C.S. Multiple Binding Sites Contribute to the Mechanism of Mixed Agonistic and Positive Allosteric Modulators of the Cannabinoid CB1 Receptor. Angew. Chem. Int. Ed. Engl. 2018, 57, 2580–2585. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Winter | Summer | SEM | p | |
|---|---|---|---|---|
| Dry matter intake 30 d, kg/d | 25.0 a | 18.5 b | 0.8 | <0.0001 |
| Milk production 30 d, kg/d | 41.7 | 37.5 | 1.7 | 0.11 |
| Fat corrected milk (4%) 30 d, kg/d | 40.7 a | 34.0 b | 1.9 | 0.02 |
| Energy balance 30 d, Mcal/d | 0.6 a | −4.5 b | 1.7 | 0.01 |
| NEFAs, µEq/L | 414.0 | 437.0 | 47.7 | 0.74 |
| Insulin, pg/mL | 20.1 | 14.2 | 2.0 | 0.06 |
| Cortisol, ng/mL | 7.8 | 6.8 | 1.8 | 0.69 |
| MDA, µM | 224.3 | 388.2 | 91.6 | 0.25 |
| TNF-α, pg/mL | 375.8 b | 1289.3 a | 161.6 | 0.001 |
| RQ | Summer | Winter | SEM | p |
|---|---|---|---|---|
| ECS genes | ||||
| CNR1 | 0.035 a | 0.064 b | 0.007 | 0.01 |
| CNR2 | 0.176 a | 0.331 b | 0.037 | 0.009 |
| MGLL | 0.551 a | 0.992 b | 0.127 | 0.03 |
| FAAH | 0.008 | 0.010 | 0.002 | 0.57 |
| NAPEPLD | 0.012 | 0.015 | 0.003 | 0.42 |
| PPARA | 0.073 | 0.056 | 0.013 | 0.36 |
| Inflammatory genes | ||||
| TRPV1 | 0.210 a | 0.487 b | 0.051 | 0.001 |
| TNFA | 0.003 | 0.003 | 0.001 | 1.00 |
| CD68 | 0.241 | 0.465 | 0.093 | 0.10 |
| NFKB | 0.148 | 0.088 | 0.034 | 0.24 |
| Oxidative stress genes | ||||
| SOD1 | 0.519 | 0.492 | 0.068 | 0.78 |
| NRF2 | 0.244 | 0.151 | 0.069 | 0.36 |
| STIP1 | 0.741 | 0.919 | 0.088 | 0.17 |
| MAP2K | 2.501 a | 1.788 b | 0.171 | 0.009 |
| GSTM1 | 0.106 | 0.086 | 0.040 | 0.73 |
| Winter | Summer | SEM | p | |
|---|---|---|---|---|
| Adipose tissue | ||||
| AEA, fmol/mg | 1.6 | 3.3 | 1.5 | 0.429 |
| 2-AG, nmol/mg | 199.7 | 199.0 | 49.0 | 0.992 |
| OEA, pmol/mg | 148.2 | 238.6 | 84.5 | 0.461 |
| PEA, pmol/mg | 34.3 | 66.7 | 21.2 | 0.295 |
| AA, nmol/mg | 1097.0 | 1023.2 | 176.7 | 0.773 |
| Plasma | ||||
| AEA, fmol/mL | 308.6 | 443.9 | 67.9 | 0.178 |
| 2-AG, nmol/mL | 16.8 | 13.9 | 2.1 | 0.350 |
| OEA, pmol/mL | 65.4 | 62.4 | 5.4 | 0.694 |
| PEA, pmol/mL | 1698.9 | 1186.1 | 579.9 | 0.544 |
| AA, nmol/mL | 251.5 | 260.8 | 28.2 | 0.820 |
| Cooled | Heat Stressed | SEM | p | |
|---|---|---|---|---|
| Dry matter intake, kg/d | 28.7 a | 24.8 b | 0.6 | 0.0005 |
| Milk production, kg/d | 32.3 a | 28.9 b | 0.9 | 0.02 |
| Fat corrected milk (4%), kg/d | 28.1 a | 24.0 b | 1.4 | 0.05 |
| Energy balance, Mcal/d | 15.2 a | 12.6 b | 0.9 | <0.0001 |
| BW gain, kg | 3.5 | 13.2 | 5.9 | 0.26 |
| NEFAs, µEq/L | 136.0 | 120.6 | 6.4 | 0.10 |
| Glucose, mg/dL | 62.8 | 59.3 | 1.8 | 0.19 |
| Cortisol, ng/mL | 1.09 | 1.03 | 0.04 | 0.35 |
| MDA, µM | 3.8 | 4.9 | 0.8 | 0.38 |
| TNF-α, pg/mL | 163.6 | 154.0 | 8.4 | 0.43 |
| Rectal temperature, morning | 37.8 b | 38.5 a | 0.08 | <0.0001 |
| Rectal temperature, afternoon | 38.4 b | 39.1 a | 0.08 | <0.0001 |
| Resting time, min/d | 586.7 | 543.2 | 18.7 | 0.11 |
| Respiration rate, morning | 28.6 b | 43.1 a | 3.2 | 0.005 |
| Respiration rate, afternoon | 48.6 | 58.6 | 3.8 | 0.08 |
| Rumination time, min/d | 517.5 | 512.9 | 9.8 | 0.7 |
| RQ | Cooled | Heat Stressed | SEM | p |
|---|---|---|---|---|
| ECS genes | ||||
| CNR1 | 0.033 | 0.030 | 0.007 | 0.80 |
| CNR2 | 0.303 | 0.283 | 0.038 | 0.71 |
| MGLL | 0.286 | 0.149 | 0.091 | 0.31 |
| FAAH | 0.007 | 0.007 | 0.002 | 0.94 |
| NAPEPLD | 0.028 | 0.017 | 0.007 | 0.30 |
| PPARA | 0.096 | 0.059 | 0.027 | 0.38 |
| Inflammatory genes | ||||
| TRPV1 | 0.091 | 0.080 | 0.015 | 0.63 |
| TNFA | 0.004 | 0.005 | 0.001 | 0.73 |
| NFKB | 0.045 | 0.038 | 0.012 | 0.68 |
| Oxidative stress genes | ||||
| STIP1 | 0.670 | 0.691 | 0.064 | 0.81 |
| MAP2K | 1.116 | 0.746 | 0.204 | 0.23 |
| Cooled | Heat Stressed | SEM | p | |
|---|---|---|---|---|
| Adipose tissue | ||||
| AEA, fmol/mg | 0.3 | 0.2 | 0.06 | 0.27 |
| 2-AG, nmol/mg | 164.2 | 159.6 | 35.6 | 0.93 |
| OEA, pmol/mg | 32.3 | 25.6 | 6.5 | 0.48 |
| PEA, pmol/mg | 5.2 | 6.8 | 1.9 | 0.57 |
| AA, nmol/mg | 0.5 | 0.5 | 0.1 | 1.00 |
| Plasma | ||||
| AEA, fmol/mL | 95.2 | 99.8 | 12.4 | 0.80 |
| 2-AG, nmol/mL | 4.7 | 5.1 | 0.5 | 0.57 |
| OEA, pmol/mL | 25.4 | 28.8 | 6.4 | 0.71 |
| PEA, pmol/mL | 1.3 | 3.1 | 1.2 | 0.31 |
| AA, nmol/mL | 178.8 | 186.0 | 21.1 | 0.81 |
| Milk | ||||
| AEA, fmol/mL | 13.4 | 18.3 | 5.6 | 0.55 |
| 2-AG, nmol/mL | 29.2 | 50.3 | 7.5 | 0.06 |
| OEA, pmol/mL | 5.3 | 5.6 | 0.8 | 0.80 |
| PEA, pmol/mL | 14.4 | 11.0 | 1.8 | 0.20 |
| AA, nmol/mL | 61.6 | 58.7 | 8.4 | 0.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kra, G.; Daddam, J.R.; Moallem, U.; Kamer, H.; Ahmad, M.; Nemirovski, A.; Contreras, G.A.; Tam, J.; Zachut, M. Effects of Environmental Heat Load on Endocannabinoid System Components in Adipose Tissue of High Yielding Dairy Cows. Animals 2022, 12, 795. https://doi.org/10.3390/ani12060795
Kra G, Daddam JR, Moallem U, Kamer H, Ahmad M, Nemirovski A, Contreras GA, Tam J, Zachut M. Effects of Environmental Heat Load on Endocannabinoid System Components in Adipose Tissue of High Yielding Dairy Cows. Animals. 2022; 12(6):795. https://doi.org/10.3390/ani12060795
Chicago/Turabian StyleKra, Gitit, Jayasimha Rayalu Daddam, Uzi Moallem, Hadar Kamer, Majdoleen Ahmad, Alina Nemirovski, G. Andres Contreras, Joseph Tam, and Maya Zachut. 2022. "Effects of Environmental Heat Load on Endocannabinoid System Components in Adipose Tissue of High Yielding Dairy Cows" Animals 12, no. 6: 795. https://doi.org/10.3390/ani12060795
APA StyleKra, G., Daddam, J. R., Moallem, U., Kamer, H., Ahmad, M., Nemirovski, A., Contreras, G. A., Tam, J., & Zachut, M. (2022). Effects of Environmental Heat Load on Endocannabinoid System Components in Adipose Tissue of High Yielding Dairy Cows. Animals, 12(6), 795. https://doi.org/10.3390/ani12060795