
The Novel-miR-659/SPP1 Interaction Regulates Fat Deposition in Castrated Male Pigs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Differentiation
2.2. Plasmid Constructs, RNA Interference, and Cell Transfection
2.3. Testosterone Test
2.4. Oil Red O Staining
2.5. Real-Time PCR
2.6. Dual Luciferase Reporter Assay
2.7. Statistical Analysis
3. Result
3.1. Adipogenic Differentiation of pBMSCs
3.2. SPP1 Overexpression Suppressed the Adipogenic Differentiation of pBMSCs
3.3. Inhibition of SPP1 Promoted the Adipogenic Differentiation of pBMSCs
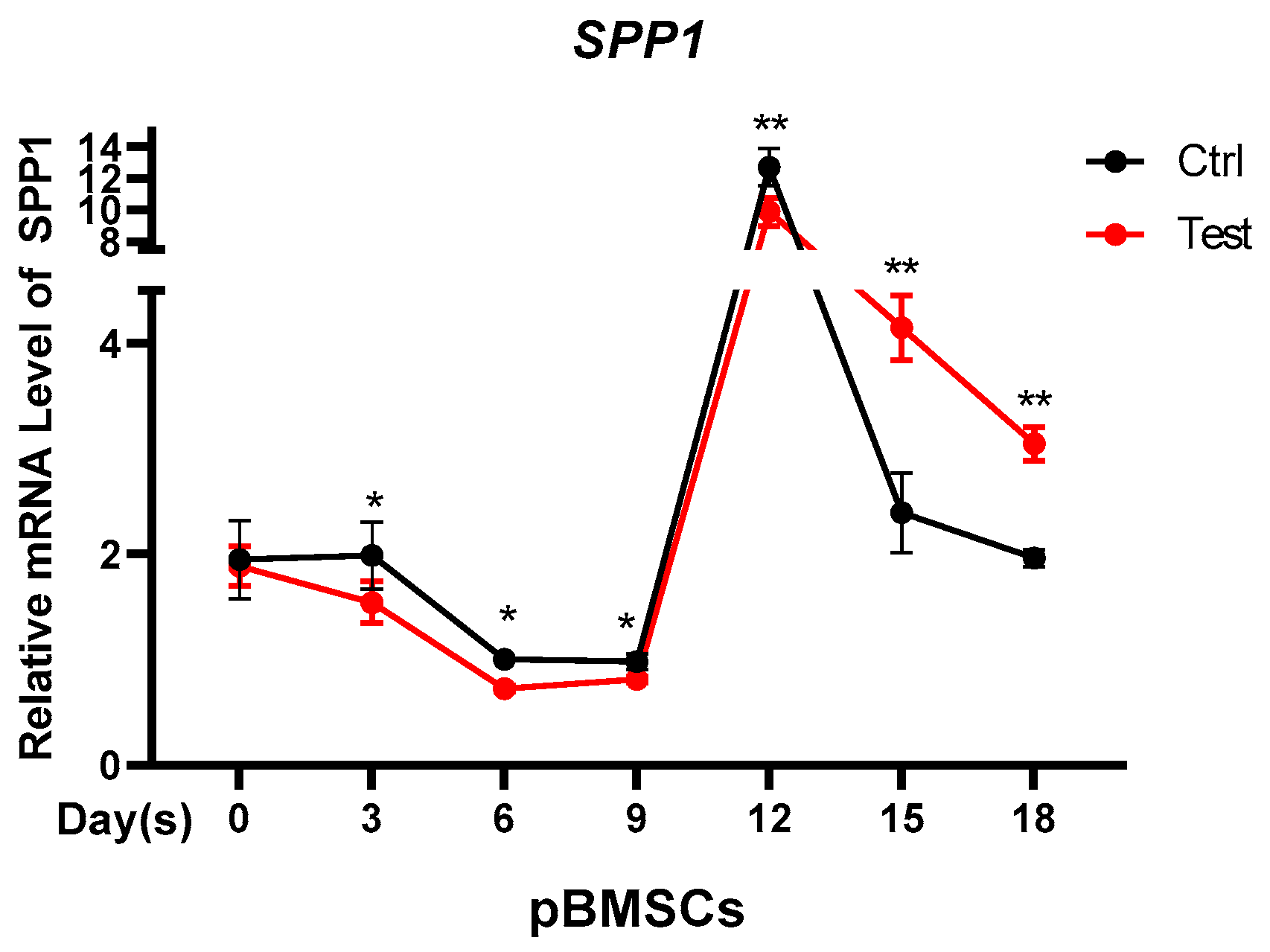
3.4. Testosterone Regulates the Expression of SPP1
3.5. Novel-miR-659 Promoted Adipogenesis of pBMSCs by Targeting SPP1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Zhang, F.C.; Qian, S.W.; Li, X.; Cui, Z.M.; Dang, Y.J.; Tang, Q.Q. Artemisinin derivatives prevent obesity by inducing browning of WAT and enhancing BAT function. Cell Res. 2016, 26, 1169–1172. [Google Scholar] [CrossRef]
- Berrington, D.G.A.; Hartge, P.; Cerhan, J.R.; Flint, A.J.; Hannan, L.; Macinnis, R.J.; Moore, S.C.; Tobias, G.S.; Anton-Culver, H.; Freeman, L.B.; et al. Body-mass index and mortality among 1.46 million white adults. N. Engl. J. Med. 2010, 363, 2211–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerradi, M.; Dereumetz, J.; Boulet, M.; Tchernof, A. Androgens, body fat Distribution and Adipogenesis. Curr. Obes. Rep. 2014, 3, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hua, L.; Chen, J.; Zhang, J.; Bai, X.; Gao, B.; Li, C.; Shi, Z.; Sheng, W.; Gao, Y.; et al. Identification and characterization of long non-coding RNAs in subcutaneous adipose tissue from castrated and intact full-sib pair Huainan male pigs. BMC Genom. 2017, 18, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossmann, M. Low Testosterone in Men with Type 2 Diabetes: Significance and Treatment. J. Clin. Endocrinol. Metab. 2011, 96, 2341–2353. [Google Scholar] [CrossRef] [PubMed]
- Christoffersen, B.O.; Gade, L.P.; Golozoubova, V.; Svendsen, O.; Raun, K. Influence of castration-induced testosterone and estradiol deficiency on obesity and glucose metabolism in male Göttingen minipigs. Steroids 2010, 75, 676–684. [Google Scholar] [CrossRef]
- Wang, K.; Li, W.; Bai, Y.; Yang, W.; Ling, Y.; Fang, M. ssc-miR-7134-3p regulates fat accumulation in castrated male pigs by targetingMARK4 gene. Int. J. Biol. Sci. 2017, 13, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Bjorntorp, P. Effects of testosterone on triglyceride uptake and mobilization in different adipose tissues in male rats in vivo. Obes. Res. 1995, 3, 113–119. [Google Scholar] [CrossRef]
- Lundstrom, K.; Matthews, K.R.; Haugen, J.E. Pig meat quality from entire males. Animal 2009, 3, 1497–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonneau, M. Use of entire males for pig meat in the European Union. Meat Sci. 1998, 49, S257–S272. [Google Scholar] [CrossRef]
- Poklukar, K.; Andek-Potokar, M.; Vrecl, M.; Batorek-Lukač, N.; Fazarinc, G.; Kress, K.; Weiler, U.; Stefanski, V.; Akrlep, M. The effect of immunocastration on adipose tissue deposition and composition in pigs. Animal 2021, 15, 100118. [Google Scholar] [CrossRef] [PubMed]
- Gispert, M.; Angels, O.M.; Velarde, A.; Suarez, P.; Perez, J.; Font, I.F.M. Carcass and meat quality characteristics of immunocastrated male, surgically castrated male, entire male and female pigs. Meat Sci. 2010, 85, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Saintilan, R.; Sellier, P.; Billon, Y.; Gilbert, H. Genetic correlations between males, females and castrates for residual feed intake, feed conversion ratio, growth rate and carcass composition traits in Large White growing pigs. J. Anim. Breed. Genet. 2012, 129, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Wolf, E.; Braun-Reichhart, C.; Streckel, E.; Renner, S. Genetically engineered pig models for diabetes research. Transgenic Res. 2014, 23, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar] [PubMed]
- Jürets, A.; Le Bras, M.; Staffler, G.; Stein, G.; Leitner, L.; Neuhofer, A.; Tardelli, M.; Turkof, E.; Zeyda, M.; Stulnig, T.M. Inhibition of Cellular Adhesion by Immunological Targeting of Osteopontin Neoepitopes Generated through Matrix Metalloproteinase and Thrombin Cleavage. PLoS ONE 2016, 11, e148333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, M.; Shinohara, M.L. Intracellular osteopontin (iOPN) and immunity. Immunol. Res. 2011, 49, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolak, T. Osteopontin–A multi-modal marker and mediator in atherosclerotic vascular disease. Atherosclerosis 2014, 236, 327–337. [Google Scholar] [CrossRef]
- Bandopadhyay, M.; Bulbule, A.; Butti, R.; Chakraborty, G.; Ghorpade, P.; Ghosh, P.; Gorain, M.; Kale, S.; Kumar, D.; Kumar, S.; et al. Osteopontin as a therapeutic target for cancer. Expert Opin. Ther. Tar. 2014, 18, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Staines, K.A.; Macrae, V.E.; Farquharson, C. The importance of the SIBLING family of proteins on skeletal mineralisation and bone remodelling. J. Endocrinol. 2012, 214, 241–255. [Google Scholar] [CrossRef] [Green Version]
- Lund, S.A.; Giachelli, C.M.; Scatena, M. The role of osteopontin in inflammatory processes. J. Cell Commun. Signal. 2009, 3, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, J.; Kim, J. Improved Cellular Response of Osteoblast Cells Using Recombinant Human Osteopontin Protein Produced by Escherichia coli. Biotechnol. Lett. 2005, 27, 1767–1770. [Google Scholar] [CrossRef] [PubMed]
- Denhardt, D.T.G.X. Osteopontin: A protein with diverse functions. FASEB J. 1993, 15, 1475–1482. [Google Scholar] [CrossRef]
- Kiefer, F.W.; Zeyda, M.; Todoric, J.; Huber, J.; Geyeregger, R.; Weichhart, T.; Aszmann, O.; Ludvik, B.; Silberhumer, G.R.; Prager, G.; et al. Osteopontin Expression in Human and Murine Obesity: Extensive Local Up-Regulation in Adipose Tissue but Minimal Systemic Alterations. Endocrinology 2008, 149, 1350–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Ambrosi, J.; Catalán, V.; Ramírez, B.; Rodríguez, A.; Colina, I.; Silva, C.; Rotellar, F.; Mugueta, C.; Gil, M.J.; Cienfuegos, J.A.; et al. Plasma Osteopontin Levels and Expression in Adipose Tissue Are Increased in Obesity. J. Clin. Endocrinol. Metab. 2007, 92, 3719–3727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahles, F.; Findeisen, H.M.; Bruemmer, D. Osteopontin: A novel regulator at the cross roads of inflammation, obesity and diabetes. Mol. Metab. 2014, 3, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Kiefer, F.W.; Neschen, S.; Pfau, B.; Legerer, B.; Neuhofer, A.; Kahle, M.; Hrabé De Angelis, M.; Schlederer, M.; Mair, M.; Kenner, L.; et al. Osteopontin deficiency protects against obesity-induced hepatic steatosis and attenuates glucose production in mice. Diabetologia 2011, 54, 2132–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, J.; Miles, P.D.; Ofrecio, J.M.; Neels, J.G.; Yu, J.G.; Resnik, J.L.; Wilkes, J.; Talukdar, S.; Thapar, D.; Johnson, K.; et al. Osteopontin is required for the early onset of high fat diet-induced insulin resistance in mice. PLoS ONE 2010, 5, e13959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomiyama, T.; Perez-Tilve, D.; Ogawa, D.; Gizard, F.; Zhao, Y.; Heywood, E.B.; Jones, K.L.; Kawamori, R.; Cassis, L.A.; Tschöp, M.H.; et al. Osteopontin mediates obesity-induced adipose tissue macrophage infiltration and insulin resistance in mice. J. Clin. Investig. 2007, 117, 2877–2888. [Google Scholar] [CrossRef]
- Wang, M.; Guo, Y.; Zhou, Y.; Yuan, W.; Li, H.; Xiong, S.; Wang, K.; Zeng, G.; Huang, Q. Secreted-Osteopontin Contributes to Brown Adipogenesis In Vitro via a CD44-Dependent Pathway. Horm. Metab. Res. 2019, 51, 741. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Xu, Y.; Yuan, W.; Wang, M.; Zhou, Y.; Chen, K.; Huang, Q. Downregulation of osteopontin inhibits browning of white adipose tissues through PI3K-AKT pathway in C57BL/6 mice. Eur. J. Pharmacol. 2020, 866, 172822. [Google Scholar] [CrossRef] [PubMed]
- Zhong, X.; Shen, X.; Wen, J.; Kong, Y.; Chu, J.; Yan, G.; Li, T.; Liu, D.; Wu, M.; Zeng, G.; et al. Osteopontin-induced brown adipogenesis from white preadipocytes through a PI3K-AKT dependent signaling. Biochem. Biophys. Res. Commun. 2015, 459, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Shou, P.; Zhang, L.; Xu, C.; Zheng, C.; Han, Y.; Li, W.; Huang, Y.; Zhang, X.; Shao, C.; et al. An Osteopontin-Integrin Interaction Plays a Critical Role in Directing Adipogenesis and Osteogenesis by Mesenchymal Stem Cells. Stem Cells 2014, 32, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Viedma, V.; Tardelli, M.; Zeyda, M.; Sibilia, M.; Burks, J.D.; Stulnig, T.M. Osteopontin-deficient progenitor cells display enhanced differentiation to adipocytes. Obes. Res. Clin. Pract. 2018, 12, 277–285. [Google Scholar] [CrossRef]
- Fitter, S.; Zannettino, A.C.W. Osteopontin in the pathophysiology of obesity: Is Opn a fat cell foe? Obes. Res. Clin. Pract. 2018, 12, 249–250. [Google Scholar] [CrossRef]
- Cai, Z.; Zhang, L.; Chen, M.; Jiang, X.; Xu, N. Castration-induced changes in microRNA expression profiles in subcutaneous adipose tissue of male pigs. J. Appl. Genet. 2014, 55, 259–266. [Google Scholar] [CrossRef]
- Bai, C.; Chen, S.; Gao, Y.; Shan, Z.; Guan, W.; Ma, Y. Multi-lineage potential research of bone marrow mesenchymal stem cells from Bama miniature pig. J. Exp. Zool. Part B Mol. Dev. Evol. 2015, 324, 671–685. [Google Scholar] [CrossRef]
- Shoelson, S.E. Inflammation and insulin resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human Adipose Tissue Is a Source of Multipotent Stem Cells. Mol. Biol. Cell 2002, 12, 4279–4295. [Google Scholar] [CrossRef]
- Nardelli, C.; Iaffaldano, L.; Ferrigno, M.; Labruna, G.; Maruotti, G.M.; Quaglia, F.; Capobianco, V.; Di Noto, R.; Del, V.L.; Martinelli, P.; et al. Characterization and predicted role of the microRNA expression profile in amnion from obese pregnant women. Int. J. Obes. 2014, 38, 466–469. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, L.; Xu, Q.; Liu, X.; Chan, S.; Luo, Y.; He, S.; Fang, M. The Novel-miR-659/SPP1 Interaction Regulates Fat Deposition in Castrated Male Pigs. Animals 2022, 12, 944. https://doi.org/10.3390/ani12080944
Xiao L, Xu Q, Liu X, Chan S, Luo Y, He S, Fang M. The Novel-miR-659/SPP1 Interaction Regulates Fat Deposition in Castrated Male Pigs. Animals. 2022; 12(8):944. https://doi.org/10.3390/ani12080944
Chicago/Turabian StyleXiao, Lianmei, Qiao Xu, Ximing Liu, Shuheng Chan, Yabiao Luo, Shuaihan He, and Meiying Fang. 2022. "The Novel-miR-659/SPP1 Interaction Regulates Fat Deposition in Castrated Male Pigs" Animals 12, no. 8: 944. https://doi.org/10.3390/ani12080944
APA StyleXiao, L., Xu, Q., Liu, X., Chan, S., Luo, Y., He, S., & Fang, M. (2022). The Novel-miR-659/SPP1 Interaction Regulates Fat Deposition in Castrated Male Pigs. Animals, 12(8), 944. https://doi.org/10.3390/ani12080944