
Unmanned Aircraft Systems as a Powerful Tool to Detect Fine-Scale Spatial Positioning and Interactions between Waterbirds at High-Tide Roosts


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
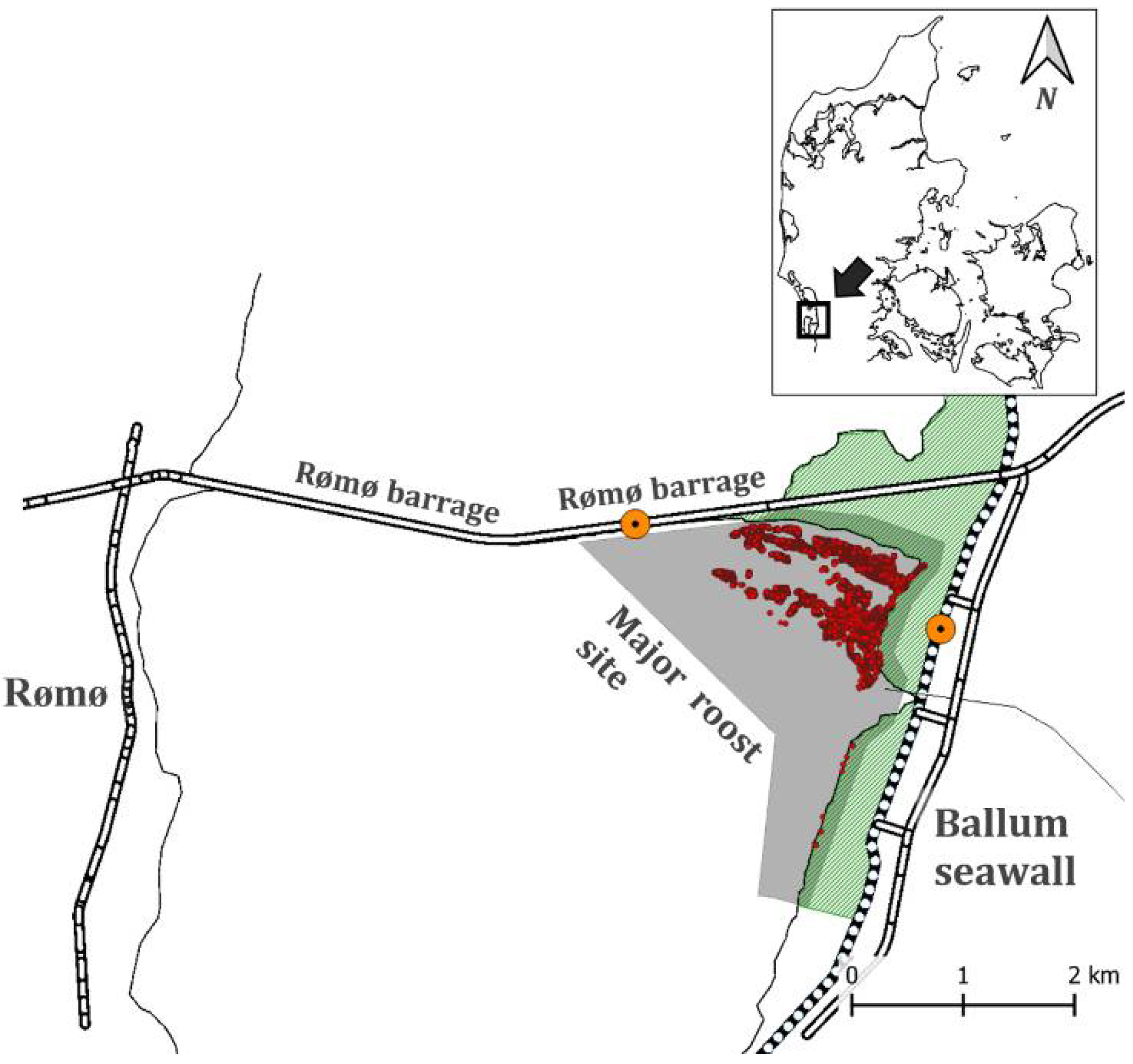
2.1. Study Area
2.2. Field Work and Data Collection
2.3. Data Preparation and Extraction
2.4. Data Analysis
2.4.1. Spatial Positioning from Different Viewing Perspectives
2.4.2. Density Patterns
2.4.3. Mixed and Non-Mixed Flocks
2.4.4. Inter-Species Interactions
2.4.5. Habitat Selection and Density
3. Results
3.1. Spatial Positioning from Different Viewing Perspectives
3.2. Density Patterns
3.3. Mixed and Non-Mixed Flocks
3.4. Inter-Species Interactions
3.4.1. Distance to Nearest Other Species
3.4.2. Individuals of Other Species within 10 m
3.5. Habitat Selection and Density
4. Discussion
4.1. A New Perspective on Roosting Dynamics and Species Interactions
4.2. Detailed Density Patterns
4.3. Flock Structure and Interactions
4.4. Habitat Selection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Anderson, K.; Gaston, K.J. Lightweight unmanned aerial vehicles will revolutionize spatial ecology. Front. Ecol. Environ. 2013, 11, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Dowding, J. Introduction to bird monitoring. In Inventory and Monitoring Toolbox: Birds; Greene, T., McNutt, K., Eds.; Department of Conservation, Te Papa Atawhai: Wellington, New Zeeland, 2012; pp. 1–33. ISBN DOCDM-578317. [Google Scholar]
- Bibby, C.; Burgess, N.; Hill, D.; Mustoe, S. Bird Census Techniques, 2nd ed.; Academic Press: London, UK, 2000; ISBN 0-12-095831-7. [Google Scholar]
- Gregory, R.; Noble, D.; Field, R.; Marchant, J.; Raven, M.; Gibbons, D. Using birds as indicators of biodiversity. Ornis Hungarica 2003, 12, 11–24. [Google Scholar]
- Recher, H.F. Some Aspects of the Ecology of Migrant Shorebirds. Ecology 1966, 47, 393–407. [Google Scholar] [CrossRef]
- Burton, N.H.K.; Evans, P.R. Aggressive behaviours and correlates of dominance in Purple Sandpipers Calidris maritima at a communal winter roost. Ibis 2001, 143, 248–254. [Google Scholar] [CrossRef]
- Laursen, K.; Frikke, J. Rastende Vandfugle i Vadehavet 1980–2010; Meltofte, H., Asbirk, S., Drachmann, J., Fjeldså, J., Jensen, S.B., Kampp, K., Petersen, B.S., Eds.; Dansk Ornitologisk Forening, BirdLife Denmark: Copenhagen, Denmark, 2013; Volume 107, ISBN 9788790310257. [Google Scholar]
- Manley, P.N.; Van Horne, B.; Roth, J.K.; Zielinski, W.J.; McKenzie, M.M.; Weller, T.J.; Weckerly, F.W.; Vojta, C. Multiple Species Inventory and Monitoring Technical Guide; United States Department of Agriculture, Forest Service: Washington, DC, USA, 2006; ISBN 10.2737/WO-GTR-73.
- Rogers, D.I.; Battley, P.F.; Piersma, T.; Van Gils, J.A.; Rogers, K.G. High-tide habitat choice: Insights from modelling roost selection by shorebirds around a tropical bay. Anim. Behav. 2006, 72, 563–575. [Google Scholar] [CrossRef] [Green Version]
- Koffijberg, K.; Blew, J.; Eskildsen, K.; Guenther, K.; Koks, B.; Laursen, K.; Rasmussen, L.-M.; Potel, P.; Suedbeck, P. High Tide Roosts in the Wadden Sea. A Review of Bird Distribution, Protection Regimes and Potential Sources of Anthropogenic Disturbance. A Report of the Wadden Sea Plan Project 34; Wadden Sea Ecosystem No. 16; Common Wadden Sea Secretariat, Trilateral Monitoring and Assessment Group, Joint Monitoring Group of Migratory Birds in the Wadden Sea: Wilhelmshaven, Germany, 2003; ISSN 0946-896X. [Google Scholar]
- Beauchamp, G. What is the magnitude of the group-size effect on vigilance? Behav. Ecol. 2008, 19, 1361–1368. [Google Scholar] [CrossRef] [Green Version]
- Peiman, K.S.; Robinson, B.W. Ecology and evolution of resource-related heterospecific aggression. Q. Rev. Biol. 2010, 85, 133–158. [Google Scholar] [CrossRef] [PubMed]
- Novcic, I. Aggression in Flocks of Foraging Shorebirds during Spring Stopover at Delaware Bay, USA. Waterbirds 2018, 41, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Melcalfe, N.B.; Furness, R.W. Aggression in shorebirds in relation to flock density and composition. Ibis 1986, 129, 553–563. [Google Scholar] [CrossRef]
- Peters, K.A.; Otis, D.L. Shorebird roost-site selection at two temporal scales: Is human disturbance a factor? J. Appl. Ecol. 2007, 44, 196–209. [Google Scholar] [CrossRef]
- Martin, J.; Edwards, H.H.; Burgess, M.A.; Percival, H.F.; Fagan, D.E.; Gardner, B.E.; Ortega-Ortiz, J.G.; Ifju, P.G.; Evers, B.S.; Rambo, T.J. Estimating distribution of hidden objects with drones: From tennis balls to manatees. PLoS ONE 2012, 7, e38882. [Google Scholar] [CrossRef] [PubMed]
- Burger, J.; Carlucci, S.A.; Jeitner, C.W.; Niles, L. Habitat choice, disturbance, and management of foraging shorebirds and gulls at a migratory stopover. J. Coast. Res. 2007, 23, 1159–1166. [Google Scholar] [CrossRef]
- Scheiffarth, G.; Nehls, G.; Austen, I. Modelling distribution of shorebirds on tidal flats in the Wadden Sea and visualisation of results with the GIS IDRISI. In IDRISI GIS 96; Lorup, E., Strobel, J., Eds.; Selbstverlag des Instituts für Geographie der Universität: Salzburg, Germany, 1996; Volume 25, ISBN 3-85283-010-9. [Google Scholar]
- Rasmussen, L.M.; Frikke, J.; Grell, M.B.; Vikstrøm, T. The Danish Wadden Sea as Breeding Ground and Crossroad for Birds; Dansk Ornitologisk Forening, BirdLife Denmark: Copenhagen, Denmark, 2008; ISBN 8790310519. [Google Scholar]
- Dulava, S.; Bean, W.T.; Richmond, O.M.W. Environmental Reviews and Case Studies: Applications of Unmanned Aircraft Systems (UAS) for Waterbird Surveys. Environ. Pract. 2015, 17, 201–210. [Google Scholar] [CrossRef]
- Sasse, D.B. Job-related mortality of wildlife workers in the United States, 1937–2000. Wildl. Soc. Bull. 2003, 31, 1015–1020. [Google Scholar] [CrossRef]
- Drever, M.C.; Chabot, D.; O’Hara, P.D.; Thomas, J.D.; Breault, A.; Millikin, R.L.; O’hara, P.D.; Thomas, J.D.; Breault, A.; Millikin, R.L. Evaluation of an unmanned rotorcraft to monitor wintering waterbirds and coastal habitats in british columbia, canada. J. Unmanned Veh. Syst. 2015, 3, 259–267. [Google Scholar] [CrossRef]
- Hodgson, J.C.; Mott, R.; Baylis, S.M.; Pham, T.T.; Wotherspoon, S.; Kilpatrick, A.D.; Raja Segaran, R.; Reid, I.; Terauds, A.; Koh, L.P. Drones count wildlife more accurately and precisely than humans. Methods Ecol. Evol. 2018, 9, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, J.C.; Baylis, S.M.; Mott, R.; Herrod, A.; Clarke, R.H. Precision wildlife monitoring using unmanned aerial vehicles. Sci. Rep. 2016, 6, 22574. [Google Scholar] [CrossRef] [Green Version]
- Hughes, A.; Teuten, E.; Starnes, T.; Cowie, N.; Swinfield, T.; Humpidge, R.; Williams, J.; Bridge, D.; Casey, C.; Asque, A.; et al. Drones for GIS—Best Practice; Version 2; Royal Society for the Protection of Birds Conservation: Sandy, UK, 2020. [Google Scholar]
- DJI PHANTOM 4 PRO Specs. Available online: https://www.dji.com/dk/phantom-4-pro/info#specs (accessed on 1 November 2021).
- Ministry of Transport. Bekendtgørelse om Flyvning Med Droner uden for Bymæssigt Område, j.nr. TS30503-03440; Retsinformation: Viborg, Denmark, 2021.
- Van Etten, J.; Sumner, M.; Cheng, J.; Baston, D.; Bevan, A.; Bivand, R.; Busetto, L.; Canty, M.; Fasoli, B.; Forrest, D.; et al. Package “Raster”, Geographic Data Analysis and Modeling. Available online: https://rspatial.org/raster (accessed on 28 January 2022).
- Kassambara, A. Package ‘Ggpubr’. Available online: https://rpkgs.datanovia.com/ggpubr/BugReports (accessed on 5 February 2022).
- Claus, A.; Wilke, O. Package ‘Cowplot’. Available online: https://wilkelab.org/cowplotBugReports (accessed on 2 February 2022).
- Silverman, B.W. Density Estimation for Statistics and Data Analysis. Monogr. Stat. Appl. Probab. 1986, 33, 9–13. [Google Scholar] [CrossRef]
- DiCiccio, T.J.; Efron, B. Bootstrap Confidence Intervals. Stat. Sci. 1996, 11, 189–228. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- D’Agostino, R.B.; Belanger, A.; D’Agostino(jr), R.B. A suggestion for using powerful and informative tests of normality. Am. Stat. 1990, 44, 316–321. [Google Scholar]
- Anscombe, F.J.; Glynn, W.J. Distribution of the kurtosis statistic b2 for normal statistics. Biometrika 1983, 70, 227–234. [Google Scholar] [CrossRef]
- Wilcoxon, F. Individual comparisons of grouped data by ranking methods. J. Econ. Entomol. 1946, 39, 269. [Google Scholar] [CrossRef] [PubMed]
- Levene, H. Robust tests for equality of variances. In Contributions to Probability and Statistics: Essays in Honor of Harold Hotelling; Olkin, I., Ghurye, S.G., Hoeffding, W., Madow, W.G., Mann, H.B., Eds.; Stanford University Press: Palo Alto, CA, USA, 1960; pp. 278–292. [Google Scholar]
- Spearman, C. The Proof and Measurement of Association between Two Things. Am. J. Psychol. 1904, 15, 72–101. [Google Scholar] [CrossRef]
- Buehler, D.M. Drone vs. human surveys of breeding shorebirds. Wader Study 2020, 127, 2018–2020. [Google Scholar] [CrossRef]
- Inoue, S.; Yamamoto, S.; Ringhofer, M.; Mendonça, R.S.; Pereira, C.; Hirata, S. Spatial positioning of individuals in a group of feral horses: A case study using drone technology. Mammal Res. 2019, 64, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, L.M. Optællinger af Kolonirugende Fugle på Fotos Optaget Med Drone ved Sneum Engsø, på Langli og holme i Limfjorden maj 2017; Report from Tidal Consult to Aarhus Universitet: Copenhagen, Denmark, 2017; pp. 4–98. [Google Scholar]
- Laursen, K.; Frikke, J.; Kahlert, J. Accuracy of ‘total counts’ of waterbirds from aircraft in coastal waters. Wildl. Biol. 2008, 14, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Scarton, F.; Valle, R.G.; Scarton, F. Drones Improve Effectiveness and Reduce Disturbance of Censusing Common Redshanks Tringa totanus Breeding on Salt Marshes. Ardea-Wageningen 2020, 107, 275–282. [Google Scholar] [CrossRef]
- Linchant, J.; Lisein, J.; Semeki, J.; Lejeune, P.; Vermeulen, C. Are unmanned aircraft systems (UASs) the future of wildlife monitoring? A review of accomplishments and challenges. Mamm. Rev. 2015, 45, 239–252. [Google Scholar] [CrossRef]
- Hong, S.J.; Han, Y.; Kim, S.Y.; Lee, A.Y.; Kim, G. Application of Deep-Learning Methods to Bird Detection Using Unmanned Aerial Vehicle Imagery. Sensors 2019, 19, 1651. [Google Scholar] [CrossRef] [Green Version]
- Juanes, F. Population Density and Body Size in Birds. Am. Soc. Nat. 1986, 128, 921–929. [Google Scholar] [CrossRef]
- Ryeland, J.; Weston, M.A.; Symonds, M.R.E. Leg length and temperature determine the use of unipedal roosting in birds. J. Avian Biol. 2019, 50, 1–9. [Google Scholar] [CrossRef]
- Dias, M.P.; Granadeiro, J.P.; Lecoq, M.; Santos, C.D.; Palmeirim, J.M. Distance to high-tide roosts constrains the use of foraging areas by dunlins: Implications for the management of estuarine wetlands. Biol. Conserv. 2006, 131, 446–452. [Google Scholar] [CrossRef]
- Laursen, K.; Kahlert, J.; Frikke, J. Factors affecting escape distances of staging waterbirds. Wildl. Biol. 2005, 11, 13–19. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 27 September | 23 October | |||||||
|---|---|---|---|---|---|---|---|---|
| Species | n | Median [CI] | Skewness (Sign.) | Kurtosis (Sign.) | n | Median [CI] | Skewness (Sign.) | Kurtosis (Sign.) |
| Dunlin | 49,068 | 4.05 [4.03; 4.06] | −0.60 (***) | 3.25 (***) | 21,375 | 7.95 [7.89; 8.01] | −0.72 (***) | 2.66 (***) |
| Golden Plover | - | - | - | 11,845 | 3.89 [3.85; 3.91] | 0.38 (***) | 2.39 (*) | |
| Oystercatcher | 1632 | 3.12 [3.05; 3.20] | 0.12 (*) | 1.81 (ns) | 1286 | 0.57 [0.55; 0.61] | 0.53 (***) | 2.78 (ns) |
| Bar-tailed Godwit | 2201 | 0.45 [0.43; 0.46] | 0.29 (***) | 1.82 (**) | 5466 | 1.28 [1.23; 1.32] | 0.09 (**) | 1.95 (***) |
| Avocet | 958 | 0.45 [0.42; 0.47] | 0.04 (ns) | 2.05 (***) | 1673 | 0.03 [0.02; 0.04] | 0.60 (***) | 2.99 (ns) |
| Wigeon | - | - | - | 25,773 | 0.33 [0.33; 0.33] | 0.42 (***) | 2.15 (***) | |
| Northern Pintail | 9491 | 0.17 [0.16; 0.17] | 1.01 (***) | 4.00 (***) | 1166 | 0.02 [0.02; 0.03] | −1.04 (ns) | 3.86 (*) |
| Shelduck | 8005 | 0.10 [0.10; 0.11] | 0.02 (ns) | 2.28 (***) | 472 | 0.01 [0.01; 0.01] | 0.76 (***) | 2.25 (**) |
| Barnacle Goose | 55 | 0.01 [0.01; 0.01] | 0.57 (ns) | 1.62 (***) | 204 | 0.10 [0.09; 0.10] | −0.41 (*) | 3.90 (*) |
| Greylag Goose | 508 | 0.03 [0.03; 0.04] | 1.25 (*) | 3.13 (ns) | 582 | 0.04 [0.03; 0.04] | 0.24 (*) | 2.60 (ns) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castenschiold, J.H.F.; Bregnballe, T.; Bruhn, D.; Pertoldi, C. Unmanned Aircraft Systems as a Powerful Tool to Detect Fine-Scale Spatial Positioning and Interactions between Waterbirds at High-Tide Roosts. Animals 2022, 12, 947. https://doi.org/10.3390/ani12080947
Castenschiold JHF, Bregnballe T, Bruhn D, Pertoldi C. Unmanned Aircraft Systems as a Powerful Tool to Detect Fine-Scale Spatial Positioning and Interactions between Waterbirds at High-Tide Roosts. Animals. 2022; 12(8):947. https://doi.org/10.3390/ani12080947
Chicago/Turabian StyleCastenschiold, Johan H. Funder, Thomas Bregnballe, Dan Bruhn, and Cino Pertoldi. 2022. "Unmanned Aircraft Systems as a Powerful Tool to Detect Fine-Scale Spatial Positioning and Interactions between Waterbirds at High-Tide Roosts" Animals 12, no. 8: 947. https://doi.org/10.3390/ani12080947
APA StyleCastenschiold, J. H. F., Bregnballe, T., Bruhn, D., & Pertoldi, C. (2022). Unmanned Aircraft Systems as a Powerful Tool to Detect Fine-Scale Spatial Positioning and Interactions between Waterbirds at High-Tide Roosts. Animals, 12(8), 947. https://doi.org/10.3390/ani12080947