
Symptomatic Infection with Vairimorpha spp. Decreases Diapause Survival in a Wild Bumble Bee Species (Bombus griseocollis)

{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Bees’ Maintenance
2.2. Diapause Simulation
2.3. Weight and Body Size Measurement
2.4. Dissection of Gut and Ovaries
2.5. Quantification of Vairimorpha
2.6. Statistical Analysis
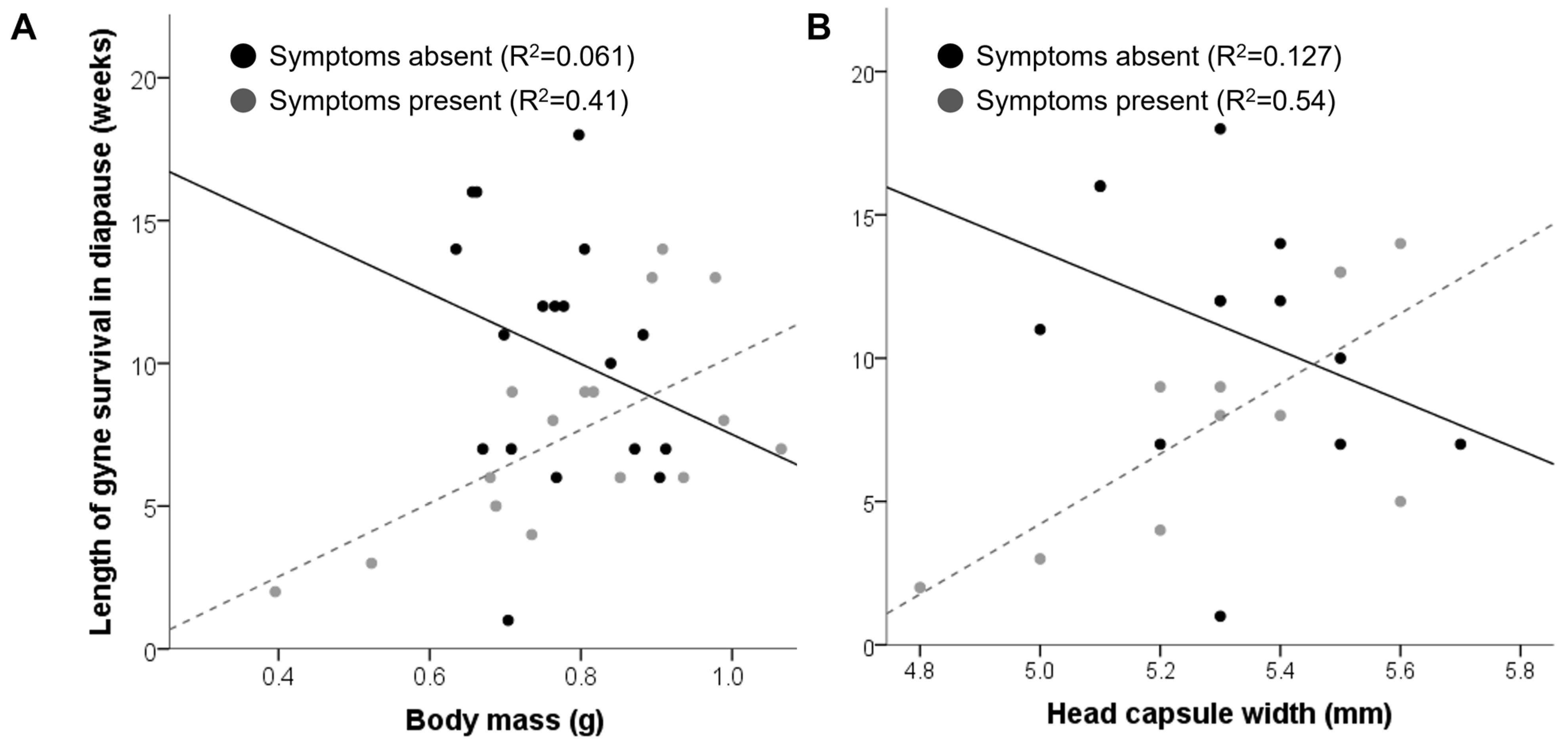
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leonhardt, S.D.; Gallai, N.; Garibaldi, L.A.; Kuhlmann, M.; Klein, A.M. Economic gain, stability of pollination and bee diversity decrease from southern to northern Europe. Basic Appl. Ecol. 2013, 14, 461–471. [Google Scholar] [CrossRef]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Meeus, I.; Pisman, M.; Smagghe, G.; Piot, N. Interaction effects of different drivers of wild bee decline and their influence on host-pathogen dynamics. Curr. Opin. Insect Sci. 2018, 26, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Bromenshenk, J.J.; Henderson, C.B.; Wick, C.H.; Stanford, M.F.; Zulich, A.W.; Jabbour, R.E.; Deshpande, S.V.; McCubbin, P.E.; Seccomb, R.A.; Welch, P.M.; et al. Iridovirus and microsporidian linked to honey bee colony decline. PLoS ONE 2010, 5, e13181. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.A.; Lim, H.C.; Lozier, J.D.; Duennes, M.A.; Thorp, R. Test of the invasive pathogen hypothesis of bumble bee decline in North America. Proc. Natl. Acad. Sci. USA 2016, 113, 4386–4391. [Google Scholar] [CrossRef]
- Cordes, N.; Huang, W.F.; Strange, J.P.; Cameron, S.A.; Griswold, T.L.; Lozier, J.D.; Solter, L.F. Interspecific geographic distribution and variation of the pathogens Nosema bombi and Crithidia species in United States bumble bee populations. J. Invertebr. Pathol. 2012, 109, 209–216. [Google Scholar] [CrossRef]
- Koch, J.B.; Strange, J.P. The Status of Bombus occidentalis and B. moderatus in Alaska with Special Focus on Nosema bombi Incidence. J. Northwest Sci. 2012, 86, 212–220. [Google Scholar]
- Tokarev, Y.S.; Huang, W.F.; Solter, L.F.; Malysh, J.M.; Becnel, J.J.; Vossbrinck, C.R. A formal redefinition of the genera Nosema and Vairimorpha (Microsporidia: Nosematidae) and reassignment of species based on molecular phylogenetics. J. Invertebr. Pathol. 2020, 169, 107279. [Google Scholar] [CrossRef]
- Baker, M.D.; Vossbrinck, C.R.; Maddox, J.V.; Undeen, A.H. Phylogenetic-relationships among Vairimorpha and Nosema species (microspora) based on ribosomal-RNA sequence data. J. Invertebr. Pathol. 1994, 64, 100–106. [Google Scholar] [CrossRef]
- Plischuk, S.; Martin-Hernandez, R.; Prieto, L.; Lucia, M.; Botias, C.; Meana, A.; Abrahamovich, A.H.; Lange, C.; Higes, M. South American native bumblebees (Hymenoptera: Apidae) infected by Nosema ceranae (Microsporidia), an emerging pathogen of honeybees (Apis mellifera). Environ. Microbiol. Rep. 2009, 1, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Whittington, R.; Winston, M.L. Effects of Nosema bombi and its treatment fumagillin on bumble bee (Bombus occidentalis) colonies. J. Invertebr Pathol 2003, 84, 54–58. [Google Scholar] [CrossRef]
- Imhoof, B.; Schmid-Hempel, P. Colony success of the bumble bee, Bombus terrestris, in relation to infections by two protozoan parasites, Crithidia bombi and Nosema bombi. Insectes Sociaux 1999, 46, 233–238. [Google Scholar] [CrossRef]
- Siviter, H.; Folly, A.J.; Brown, M.J.F.; Leadbeater, E. Individual and combined impacts of sulfoxaflor and Nosema bombi on bumblebee (Bombus terrestris) larval growth. Proc. Biol. Sci. 2020, 287, 20200935. [Google Scholar] [CrossRef] [PubMed]
- Otti, O.; Schmid-Hempel, P. A field experiment on the effect of Nosema bombi in colonies of the bumblebee Bombus terrestris. Ecol. Entomol. 2008, 33, 577–582. [Google Scholar] [CrossRef]
- Otti, O.; Schmid-Hempel, P. Nosema bombi: A pollinator parasite with detrimental fitness effects. J. Invertebr. Pathol. 2007, 96, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Rutrecht, S.T.; Klee, J.; Brown, M.J. Horizontal transmission success of Nosema bombi to its adult bumble bee hosts: Effects of dosage, spore source and host age. Parasitology 2007, 134, 1719–1726. [Google Scholar] [CrossRef]
- Rutrecht, S.T.; Brown, M.J.F. Within colony dynamics of Nosema bombi infections: Disease establishment, epidemiology and potential vertical transmission. Apidologie 2008, 39, 504–514. [Google Scholar] [CrossRef]
- Alford, D.V. A study of the hibernation of bumble bees (Hymenoptera: Bombidae) in Southern England. J. Anim. Ecol. 1969, 38, 149–170. [Google Scholar] [CrossRef]
- Michener, C.D. The Social Behavior of the Bees; Harvard University Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Amsalem, E.; Grozinger, C.M.; Padilla, M.; Hefetz, A. The physiological and genomic bases of bumble bee social behaviour. In Genomics, Physiology and Behaviour of Social Insects; Amro, Z., Clement, F.K., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 37–93. [Google Scholar]
- Beekman, M.; van Stratum, P.; Lingeman, R. Diapause survival and post-diapause performance in bumblebee queens (Bombus terrestris). Entomol. Exp. Appl. 1998, 89, 207–214. [Google Scholar] [CrossRef]
- Beekman, M.; Van Stratum, P. Does the diapause experience of bumblebee queens Bombus terrestris affect colony characteristics? Ecol. Entomol. 2000, 25, 1–6. [Google Scholar] [CrossRef]
- Bogo, G.; de Manincor, N.; Fisogni, A.; Galloni, M.; Bortolotti, L. Effects of queen mating status, pre-diapause weight and pupae’s sex on colony initiation in small-scale rearing of Bombus terrestris. Apidologie 2017, 48, 845–854. [Google Scholar] [CrossRef]
- Treanore, E.; Amsalem, E. The effect of intrinsic physiological traits on diapause survival and their underlying mechanisms in an annual bee species Bombus impatiens. Conserv. Physiol. 2020, 8, coaa103. [Google Scholar] [CrossRef] [PubMed]
- Wasielewski, O.; Wojciechowicz, T.; Giejdasz, K.; Krishnan, N. Overwintering strategies in the red mason solitary bee-physiological correlates of midgut metabolic activity and turnover of nutrient reserves in females of Osmia bicornis. Apidologie 2013, 44, 642–656. [Google Scholar] [CrossRef]
- Woodard, S.H.; Duennes, M.A.; Watrous, K.M.; Jha, S. Diet and nutritional status during early adult life have immediate and persistent effects on queen bumble bees. Conserv. Physiol. 2019, 7, coz048. [Google Scholar] [CrossRef]
- Sgolastra, F.; Arnan, X.; Pitts-Singer, T.L.; Maini, S.; Kemp, W.P.; Bosch, J. Pre-wintering conditions and post-winter performance in a solitary bee: Does diapause impose an energetic cost on reproductive success? Ecol. Entomol. 2016, 41, 201–210. [Google Scholar] [CrossRef]
- Bosch, J.; Sgolastra, F.; Kemp, W.P. Timing of eclosion affects diapause development, fat body consumption and longevity in Osmia lignaria, a univoltine, adult-wintering solitary bee. J. Insect Physiol. 2010, 56, 1949–1957. [Google Scholar] [CrossRef]
- Watrous, K.M.; Costa, C.P.; Diaz, Y.R.; Woodard, S.H. Flexibility in the critical period of nutrient sequestration in bumble bee queens. Integr. Org. Biol. 2021, 3, obab009. [Google Scholar] [CrossRef]
- Sømme, L. Effects of Glycerol on Cold-Hardiness in Insects. Can. J. Zool. 1964, 42, 87–101. [Google Scholar] [CrossRef]
- Dolezal, A.G.; Carrillo-Tripp, J.; Judd, T.M. Allen Miller, W.; Bonning, B.C.; Toth, A.L. Interacting stressors matter: Diet quality and virus infection in honeybee health. R Soc. Open Sci. 2019, 6, 181803. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Chen, Y.P.; Rivera, R.; Carroll, M.; Chambers, M.; Hidalgo, G.; de Jong, E.W. Honey bee colonies provided with natural forage have lower pathogen loads and higher overwinter survival than those fed protein supplements. Apidologie 2016, 47, 186–196. [Google Scholar] [CrossRef]
- Tritschler, M.; Vollmann, J.J.; Yanez, O.; Chejanovsky, N.; Crailsheim, K.; Neumann, P. Protein nutrition governs within-host race of honey bee pathogens. Sci. Rep. 2017, 7, 14988. [Google Scholar] [CrossRef] [PubMed]
- Mayack, C.; Naug, D. Energetic stress in the honeybee Apis mellifera from Nosema ceranae infection. J. Invertebr. Pathol. 2009, 100, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, L.L.; Grincavitch, C.; McArt, S.H. Crithidia bombi can infect two solitary bee species while host survivorship depends on diet. Parasitology 2021, 148, 435–442. [Google Scholar] [CrossRef]
- Goblirsch, M.; Huang, Z.Y.; Spivak, M. Physiological and behavioral changes in honey bees (Apis mellifera) induced by Nosema ceranae infection. PLoS ONE 2013, 8, e58165. [Google Scholar] [CrossRef]
- Hagen, M.; Dupont, Y.L. Inter-tegular span and head width as estimators of fresh and dry body mass in bumblebees (Bombus spp.). Insectes Sociaux 2013, 60, 251–257. [Google Scholar] [CrossRef]
- Amsalem, E.; Hefetz, A. The appeasement effect of sterility signaling in dominance contests among Bombus terrestris workers. Behav. Ecol. Sociobiol. 2010, 64, 1685–1694. [Google Scholar] [CrossRef]
- Mcivor, C.A.; Malone, L.A. Nosema bombi, a microsporidian pathogen of the bumble bee Bombus terrestris (L.). N. Z. J. Zool. 1995, 22, 25–31. [Google Scholar] [CrossRef]
- Bramke, K.; Muller, U.; McMahon, D.P.; Rolff, J. Exposure of larvae of the solitary bee Osmia bicornis to the honey bee pathogen Nosema ceranae affects life history. Insects 2019, 10, 380. [Google Scholar] [CrossRef]
- Schmid-Hempel, P.; Loosli, R. A contribution to the knowledge of Nosema infections in bumble bees, Bombus spp. Apidologie 1998, 29, 525–535. [Google Scholar] [CrossRef]
- Li, W.; Chen, Y.; Cook, S.C. Chronic Nosema ceranae infection inflicts comprehensive and persistent immunosuppression and accelerated lipid loss in host Apis mellifera honey bees. Int. J. Parasitol. 2018, 48, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Hahn, D.A.; Denlinger, D.L. Meeting the energetic demands of insect diapause: Nutrient storage and utilization. J. Insect Physiol. 2007, 53, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Chole, H.; Woodard, S.H.; Bloch, G. Body size variation in bees: Regulation, mechanisms, and relationship to social organization. Curr. Opin. Insect Sci. 2019, 35, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Treanore, E.D.; Kiner, J.M.; Kerner, M.E.; Amsalem, E. Shift in worker physiology and gene expression pattern from reproductive to diapause-like with colony age in the bumble bee Bombus impatiens. J. Exp. Biol. 2020, 223, jeb218768. [Google Scholar] [CrossRef]
- Oyen, K.J.; Jardine, L.E.; Parsons, Z.M.; Herndon, J.D.; Strange, J.P.; Lozier, J.D.; Dillon, M.E. Body mass and sex, not local climate, drive differences in chill coma recovery times in common garden reared bumble bees. J. Comp. Physiol. B 2021, 191, 843–854. [Google Scholar] [CrossRef]
- Owen, R.E. Body size variation and optimal body size of bumble bee queens (Hymenoptera, Apidae). Can. Entomol. 1988, 120, 19–27. [Google Scholar] [CrossRef]
- Roberts, S.P.; Harrison, J.F.; Dudley, R. Allometry of kinematics and energetics in carpenter bees (Xylocopa varipuncta) hovering in variable-density gases. J. Exp. Biol. 2004, 207, 993–1004. [Google Scholar] [CrossRef]
- Vesterlund, S.R.; Lilley, T.M.; van Ooik, T.; Sorvari, J. The effect of overwintering temperature on the body energy reserves and phenoloxidase activity of bumblebee Bombus lucorum queens. Insectes Sociaux 2014, 61, 265–272. [Google Scholar] [CrossRef]
- Tyler, E.R.; Adams, S.; Mallon, E.B. An immune response in the bumblebee, Bombus terrestris leads to increased food consumption. BMC Physiol. 2006, 6, 6. [Google Scholar] [CrossRef]
- Brunner, F.S.; Schmid-Hempel, P.; Barribeau, S.M. Protein-poor diet reduces host-specific immune gene expression in Bombus terrestris. Proc. Biol. Sci. 2014, 281, 20140128. [Google Scholar] [CrossRef]
- Jones, B.; Shipley, E.; Arnold, K.E. Social immunity in honeybees-Density dependence, diet, and body mass trade-offs. Ecol. Evol. 2018, 8, 4852–4859. [Google Scholar] [CrossRef] [PubMed]
- Holt, H.L.; Aronstein, K.A.; Grozinger, C.M. Chronic parasitization by Nosema microsporidia causes global expression changes in core nutritional, metabolic and behavioral pathways in honey bee workers (Apis mellifera). BMC Genom. 2013, 14, 799. [Google Scholar] [CrossRef] [PubMed]
- Mayack, C.; Natsopoulou, M.E.; McMahon, D.P. Nosema ceranae alters a highly conserved hormonal stress pathway in honeybees. Insect Mol. Biol. 2015, 24, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Fisher, F.M., Jr.; Sanborn, R.C. Production of insect juvenile hormone by the microsporidian parasite Nosema. Nature 1962, 194, 1193. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.J.; Singh, A.; Schilder, R.J.; López-Uribe, M.M. Squash bees host high diversity and prevalence of parasites in the northeastern United States. J. Invertebr. Pathol. 2022, 195, 107848. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlova, M.; Porter, M.; Hines, H.M.; Amsalem, E. Symptomatic Infection with Vairimorpha spp. Decreases Diapause Survival in a Wild Bumble Bee Species (Bombus griseocollis). Animals 2023, 13, 1656. https://doi.org/10.3390/ani13101656
Orlova M, Porter M, Hines HM, Amsalem E. Symptomatic Infection with Vairimorpha spp. Decreases Diapause Survival in a Wild Bumble Bee Species (Bombus griseocollis). Animals. 2023; 13(10):1656. https://doi.org/10.3390/ani13101656
Chicago/Turabian StyleOrlova, Margarita, Monique Porter, Heather M. Hines, and Etya Amsalem. 2023. "Symptomatic Infection with Vairimorpha spp. Decreases Diapause Survival in a Wild Bumble Bee Species (Bombus griseocollis)" Animals 13, no. 10: 1656. https://doi.org/10.3390/ani13101656
APA StyleOrlova, M., Porter, M., Hines, H. M., & Amsalem, E. (2023). Symptomatic Infection with Vairimorpha spp. Decreases Diapause Survival in a Wild Bumble Bee Species (Bombus griseocollis). Animals, 13(10), 1656. https://doi.org/10.3390/ani13101656