

Isolation and Characterization of Swinepox Virus from Outbreak in Russia
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Virus Isolation
2.3. DNA Isolation and Amplification
2.4. Phylogenetic Study
2.5. Animal Experiment
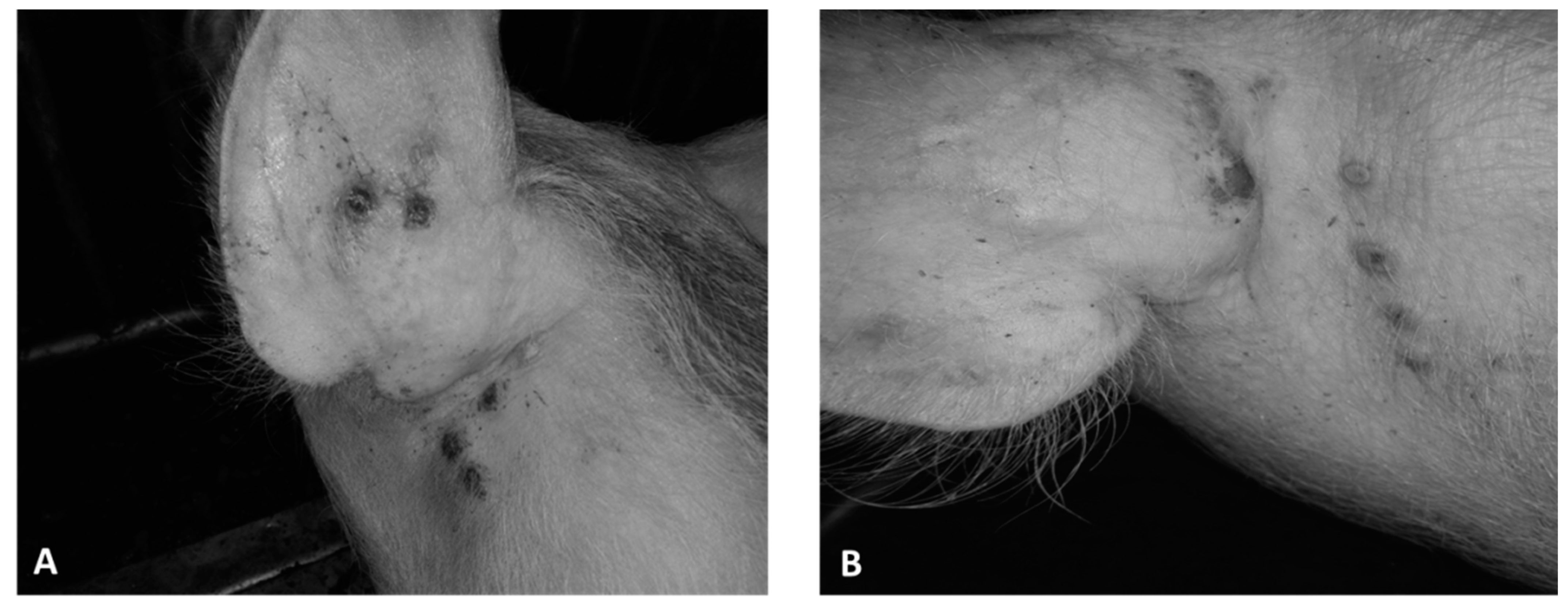
3. Results
3.1. Virus Isolation and Adaptation
3.2. SWPV Identification
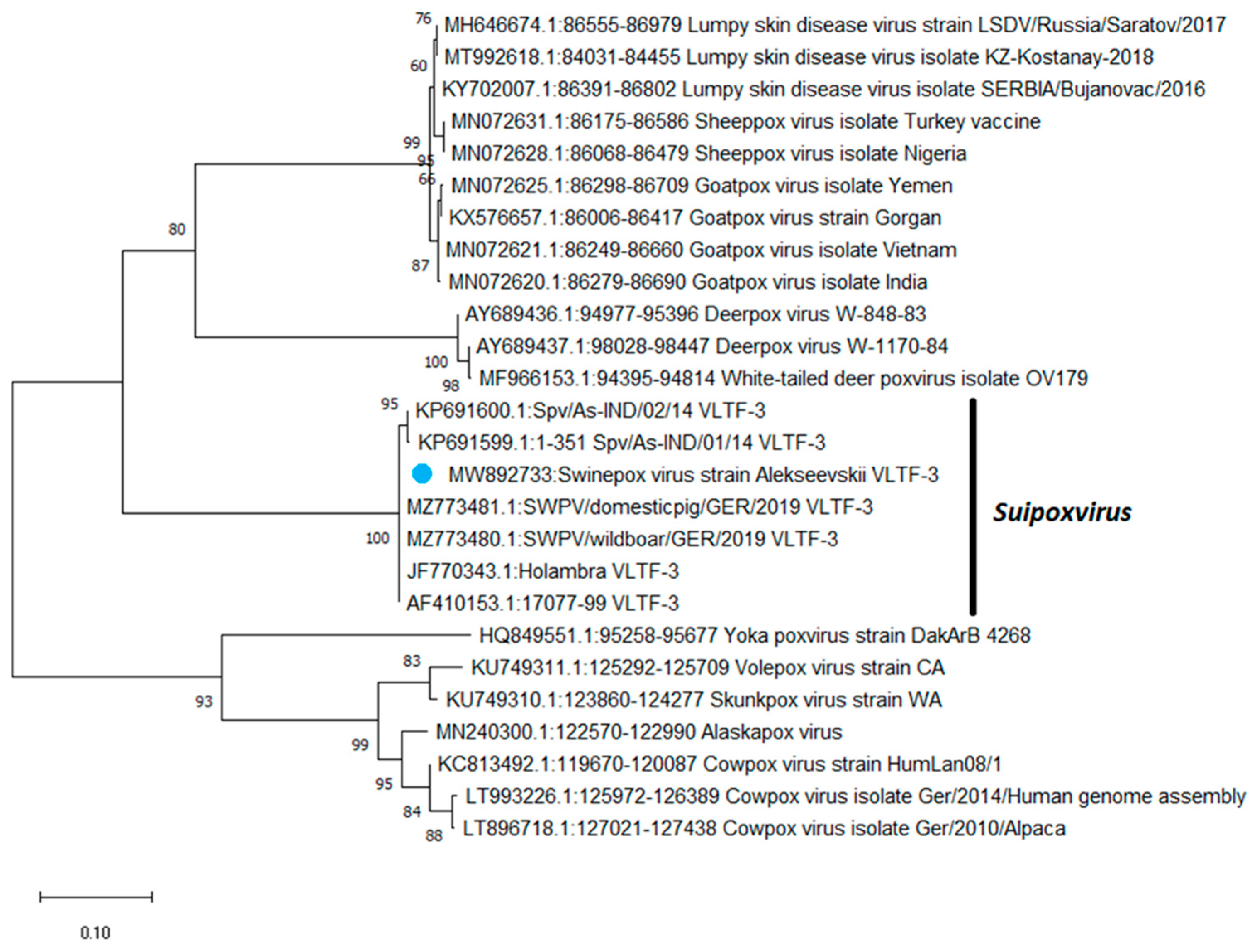
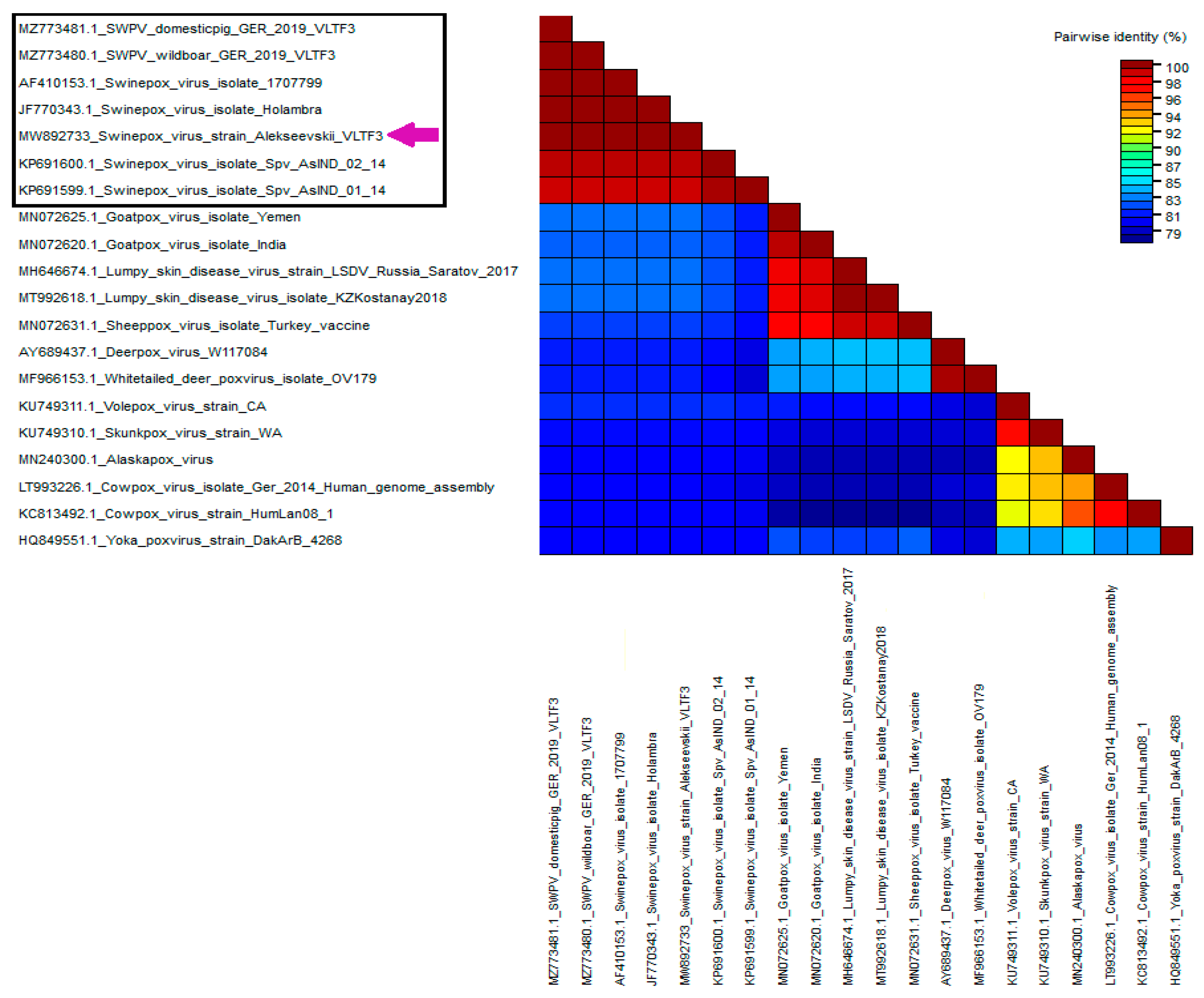
3.3. Phylogenetic Study
3.4. Animal Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jubb, T.F.; Ellis, T.M.; Peet, R.L.; Parkinson, J. Swinepox in pigs in northern Western Australia. Aust. Vet. J. 1992, 69, 99. [Google Scholar] [CrossRef]
- McNutt, S.H.; Murray, C.; Purwin, P. Swine pox. JAVMA 1929, 74, 752. [Google Scholar]
- De Boer, G.F. Swinepox, virus isolation, experimental infections and the differentiation from vaccinia virus infections. Arch. Virol. 1975, 49, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.B.; Olson, L.D. Epizootic of concurrent cutaneous streptococcal abscesses and swinepox in a herd of swine. Am. J. Vet. Med. Assoc. 1978, 172, 676–680. [Google Scholar]
- Manickam, R.; Mohan, M. A note on outbreak of swinepox in large White Yorkshire piglets. Indian J. Vet. Med. 1987, 7, 71–72. [Google Scholar]
- Ouchi, M.; Fujiwara, M.; Hatano, Y.; Yao, M.; Uno, F.; Yoshida, M.; Yamada, M.; Nii, S. An outbreak of swine pox in Okayama prefecture. J. Japan Vet. Med. Assoc. 1988, 41, 870–874. [Google Scholar] [CrossRef]
- Borst, G.H.; Kimman, T.G.; Gielkens, A.L.; van der Kamp, J.S. Four sporadic cases of congenital swinepox. Vet. Rec. 1990, 127, 61–63. [Google Scholar]
- Moorkamp, L.; Beineke, A.; Kaim, U.; Diesterbeck, U.; Urstadt, S.; Czerny, C.P.; Rüberg, H.; Beilage, E.G. Swinepox—Skin disease with sporadic occurrence. Dtsch. Tierarztl. Wochenschr. 2008, 115, 162–166. [Google Scholar]
- Medaglia, M.L.; Pereira, A.C.; Freitas, T.R.; Damaso, C.R. Swinepox virus oubreak, Brazil, 2011. Emerg. Infect. Dis. 2011, 17, 1976–1978. [Google Scholar] [CrossRef]
- Mariano, V.; Nardi, A.; Vergari, E.; Carletti, F.; Barbieri, L.; Cardeti, G. Poxvirus in a swine farm in Italy: A sporadic outbreak. Large Anim. Rev. 2015, 21, 219–220. [Google Scholar]
- Durlav, P.B.; Biswajyoti, B.; Mousumi, B.; Parikshit, K.; Samsun, N.; Lakshya, J.D.; Puja, M.; Das, S.K. Detection and characterization of Swinepox virus from pig population of Assam, a North Eastern state of India. Indian J. Anim. Res. 2017, 52, 1337–1342. [Google Scholar]
- Thachamvally, R.; Sanjay, B.; Naveen, K.; Naresh, J.; Bidhan, C.B.; Gulshan, N.; Nand, K.M.; Devan, A.; Taruna, A.; Rajesh, K.V.; et al. Isolation and genetic characterization of swinepox virus from pigs in India. Microbiol. Infect. Dis. 2016, 46, 60–65. [Google Scholar]
- Olinda, R.G.; Maia, L.A.; Cargnelutti, J.F.; Gois, R.; Batista, J.S.; Dantas, A.F.; Riet-Correa, F. Swinepox dermatitis in backyard pigs in Northeastern Brazil. Pesq. Vet. Bras. 2016, 36, 468–472. [Google Scholar] [CrossRef]
- Kaiser, F.K.; Wiedemann, A.; Kühl, B.; Menke, L.; Beineke, A.; Baumgärtner, W.; Wohlsein, P.; Rigbers, K.; Becher, P.; Peters, M.; et al. Swinepox Virus Strains Isolated from Domestic Pigs and Wild Boar in Germany Display Altered Coding Capacity in the Terminal Genome Region Encoding for Species-Specific Genes. Viruses 2021, 13, 2038. [Google Scholar] [CrossRef]
- House, J.A.; House, C.A. Swine pox. In Diseases of Swine; Leman, A.D., Straw, B.E., Mengeling, W.L., D’Allaire, S., Taylor, D.J., Eds.; Iowa State University: Ames, IA, USA, 1994; pp. 358–361. [Google Scholar]
- Munz, E.; Dumbell, K. Swinepox. In Infectious Diseases of Livestock; Coetzer, J.A.W., Thomson, G.R., Tustin, R.C., Eds.; Oxford University Press: New York, NY, USA, 1994; pp. 627–629. [Google Scholar]
- Kasza, L.; Griesemer, R.A. Experimental swine pox. Am. J. Vet. Res. 1962, 23, 443–450. [Google Scholar]
- Medaglia, M.L.G.; Sá, N.M.; Correa, I.A.; Costa, L.J.; Damaso, C.R. One-step duplex polymerase chain reaction for the detection of swinepox and vaccinia viruses in skin lesions of swine with poxvirus-related disease. J. Virol. Methods 2015, 219, 10–13. [Google Scholar] [CrossRef]
- Murphy, F.A.; Gibbs, E.P.J.; Horzinek, M.C.; Studdert, M.J. Herpesviridae. In Veterinary Virology; Murphy, F.A., Gibbs, E.P., Studdert, M.J., Horzinek, M.C., Eds.; Academic Press: San Diego, CA, USA, 1999; Volume 3, pp. 301–311. [Google Scholar]
- Timina, A.; Biryuchenkova, M. Gene diagnostics of swinepox. Proc. Fed. Cent. Anim. Health 2020, 17, 86–91. [Google Scholar]
- Freshney, R.I. Culture of Animal Cells: A Manual of Basic Techniques; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2010. [Google Scholar]
- Reed, L.J.; Muench, H.A. Simple method of estimating fifty per cent end points. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Mittal, D.; Mahajan, V.; Pathak, D.; Filia, G. Differential diagnosis of swine pox during an outbreak. Indian Vet. J. 2011, 88, 9–11. [Google Scholar]
- Singh, J.L.; Gupta, D.K.; Kumar, M.; Chandra, R.; Shukla, S.K.; Kumar, S.; Kumar, S. Clinico-pathological observation and therapeutic management of swine pox in a piggery farm. Indian J. Vet. Med. 2005, 25, 68–70. [Google Scholar]
- Leland, D.S.; Ginocchio, C.C. Role of Cell Culture for Virus Detection in the Age of Technology. Clin. Microbiol. Rev. 2007, 20, 49–78. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.K.; Meyer, R.C. Adaptation of swinepox virus to an established cell line. Appl. Microbiol. 1972, 23, 180–182. [Google Scholar] [CrossRef]
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus tissue tropism and shedding: A quantitative study in experimentally infected sheep and goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef]
- Kitching, R.P.; Mellor, P.S. Insect transmission of capripox virus. Res. Vet. Sci. 1988, 40, 255–258. [Google Scholar] [CrossRef]
- Möller, J.; Moritz, T.; Schlottau, K.; Krstevski, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Experimental lumpy skin disease virus infection of cattle: Comparison of a field strain and a vaccine strain. Arch. Virol. 2019, 164, 2931–2941. [Google Scholar] [CrossRef]
- Puja, M.; Bora, D.P.; Samsun, N.; Barman, N.N.; Probodh, B.; Shantanu, T.; Dutta, L.J.; Das, S.K. Identification of swinepox virus from natural outbreaks in pig population of Assam. Virus. Dis. 2018, 29, 395–399. [Google Scholar]
- Junghyun, H.; Se-Hoon, P.; Jae-Young, S.; Soo-Hwan, A.; Byung-Yoon, A. Construction of recombinant swinepox viruses and expression of the classical swine fever virus E2 protein. J. Virol. Methods 2011, 93, 49–56. [Google Scholar]
- Williams, P.P.; Hall, M.R.; McFarland, M.D. Immunological responses of cross-bred and in-bred miniature pigs to swine poxvirus. Vet. Immunol. Immunopathol. 1989, 23, 149–159. [Google Scholar] [CrossRef]
- Tripathy, D.N. Swinepox virus as a vaccine vector for swine pathogens. Adv. Vet. Med. 1999, 41, 463–480. [Google Scholar]
- Foley, P.L.; Paul, P.S.; Levings, R.L.; Hanson, S.K.; Middle, L.A. Swinepox virus as a vector for the delivery of immunogens. Ann. N. Y. Acad. Sci. 1991, 646, 220–222. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Lin, H.; Li, B.; He, K.; Fan, H. Swinepox virus vector-based vaccines: Attenuation and biosafety assessments following subcutaneous prick inoculation. Vet. Res. 2018, 49, 14. [Google Scholar] [CrossRef] [PubMed]
- Huixing, L.; Zhe, M.; Lei, C.; Hongjie, F. Recombinant Swinepox Virus Expressing Glycoprotein E2 of Classical Swine Fever Virus Confers Complete Protection in Pigs upon Viral Challenge image. Front. Vet. Sci. 2017, 4, 81. [Google Scholar]
- Xu, J.; Huang, D.; Liu, S.; Lin, H.; Zhu, H.; Liu, B.; Chen, W.; Lu, C. Immune responses and protective efficacy of a recombinant swinepox virus coexpressing HA1 genes of H3N2 and H1N1 swine influenza virus in mice and pigs. Vet. Microbiol. 2013, 162, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Huixing, L.; Zhe, M.; Xin, H.; Lei, C.; Hongjie, F. Construction and immunogenicity of a recombinant swinepox virus expressing a multi-epitope peptide for porcine reproductive and respiratory syndrome virus. Sci. Rep. 2017, 7, 43990. [Google Scholar]
- Bárcena, J.; Blasco, R. Recombinant swinepox virus expressing-galactosidase: Investigation of viral host range and gene expression levels in cell culture. Virology 1998, 243, 396–405. [Google Scholar] [CrossRef]
- Afonso, C.L.; Tulman, E.R.; Lu, Z.; Zsak, L.; Osorio, F.A.; Balinsky, C.; Kutish, G.F.; Rock, D.L. The Genome of Swinepox Virus. J. Virol. 2002, 76, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Assarsson, E.; Greenbaum, J.A.; Sundström, M.; Schaffer, L.; Hammond, J.A.; Pasquetto, V.; Oseroff, C.; Hendrickson, R.C.; Lefkowitz, E.J.; Tscharke, D.C.; et al. Kinetic analysis of a complete poxvirus transcriptome reveals an immediate-early class of genes. Proc. Natl. Acad. Sci. USA 2008, 105, 2140–2145. [Google Scholar] [CrossRef]
- Stack, J.; Haga, I.R.; Schröder, M.; Bartlett, N.W.; Maloney, G.; Reading, P.C.; Fitzgerald, K.A.; Smith, G.L.; Bowie, A.G. Vaccinia virus protein A46R targets multiple Toll-like-interleukin-1 receptor adaptors and contributes to virulence. J. Exp. Med. 2005, 201, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Bowie, A.; Kiss-Toth, E.; Symons, J.A.; Smith, G.L.; Dower, S.K.; O’Neill, L.A.J. A46R and A52R from vaccinia virus are antagonists of host IL-1 and toll-like receptor signaling. Proc. Natl. Acad. Sci. USA 2000, 97, 10162–10167. [Google Scholar] [CrossRef] [PubMed]
- Keating, S.E.; Maloney, G.M.; Moran, E.M.; Bowie, A.G. IRAK-2 participates in multiple toll-like receptor signaling pathways to NFkappaB via activation of TRAF6 ubiquitination. J. Biol. Chem. 2007, 282, 33435–33443. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | No. of Animals | Infection Material | Survival % | Fever | SPW Skin Lesion | Viremia | SWPV DNA in Oral and Nasal Swabs | SWPV in Oral and Nasal Swabs |
|---|---|---|---|---|---|---|---|---|
| 1 | 5 | Clarified 10% suspension of skin scab (not titrated) | 100 | No | Yes at 5–30 dpi | No | Yes at 8–30 dpi | Yes, isolated at 11, 17, and 25 dpi |
| 2 | 5 | Adapted to PK-15 cells SWPV isolate (1 × 106 TCID50) | 100 | No | Yes at 11–30 dpi | No | Yes at 11–30 dpi | Yes, isolated at 11, 17, and 25 dpi |
| 3 | 5 | PBS | 100 | No | No | No | No | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koltsov, A.; Sukher, M.; Kholod, N.; Namsrayn, S.; Tsybanov, S.; Koltsova, G. Isolation and Characterization of Swinepox Virus from Outbreak in Russia. Animals 2023, 13, 1786. https://doi.org/10.3390/ani13111786
Koltsov A, Sukher M, Kholod N, Namsrayn S, Tsybanov S, Koltsova G. Isolation and Characterization of Swinepox Virus from Outbreak in Russia. Animals. 2023; 13(11):1786. https://doi.org/10.3390/ani13111786
Chicago/Turabian StyleKoltsov, Andrey, Mikhail Sukher, Natalia Kholod, Sanzhi Namsrayn, Sodnom Tsybanov, and Galina Koltsova. 2023. "Isolation and Characterization of Swinepox Virus from Outbreak in Russia" Animals 13, no. 11: 1786. https://doi.org/10.3390/ani13111786
APA StyleKoltsov, A., Sukher, M., Kholod, N., Namsrayn, S., Tsybanov, S., & Koltsova, G. (2023). Isolation and Characterization of Swinepox Virus from Outbreak in Russia. Animals, 13(11), 1786. https://doi.org/10.3390/ani13111786