
Identification of Risk Areas for Gloydius Snakebites in South Korea



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
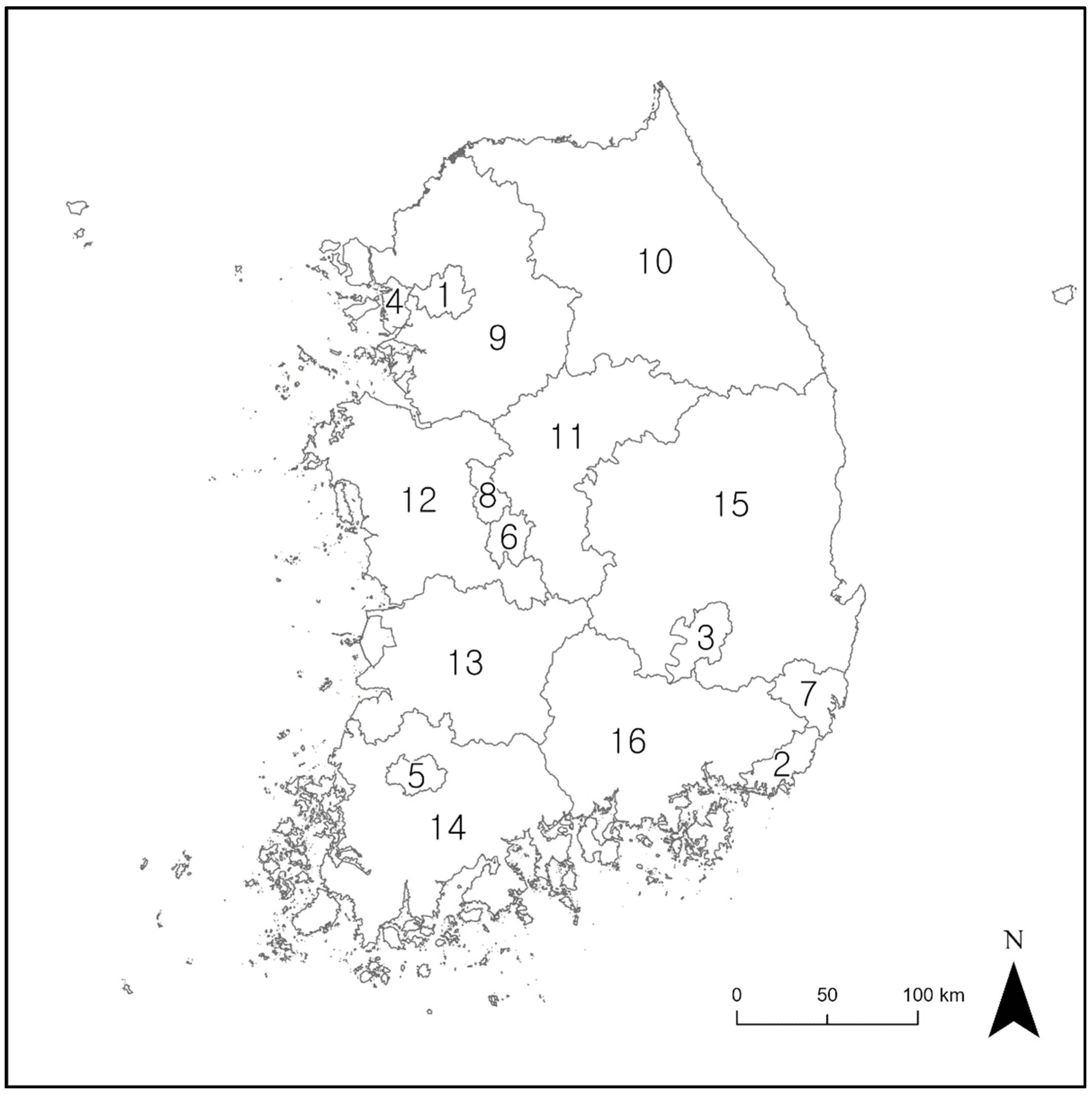
2.1. Study Area
2.2. Target Species
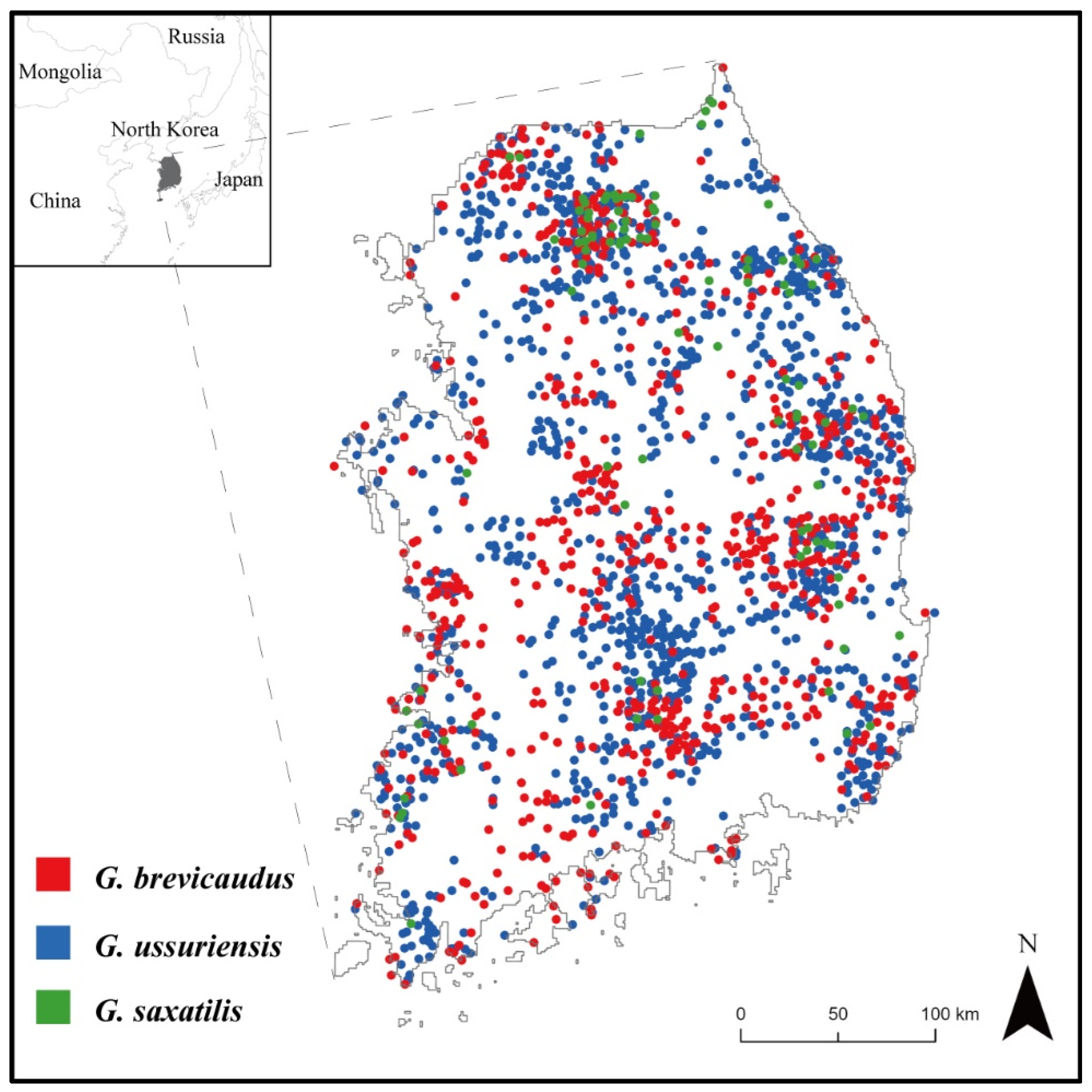
2.3. Data Collection
2.4. Species Distribution Modeling
3. Results
3.1. Model Performance Evaluation
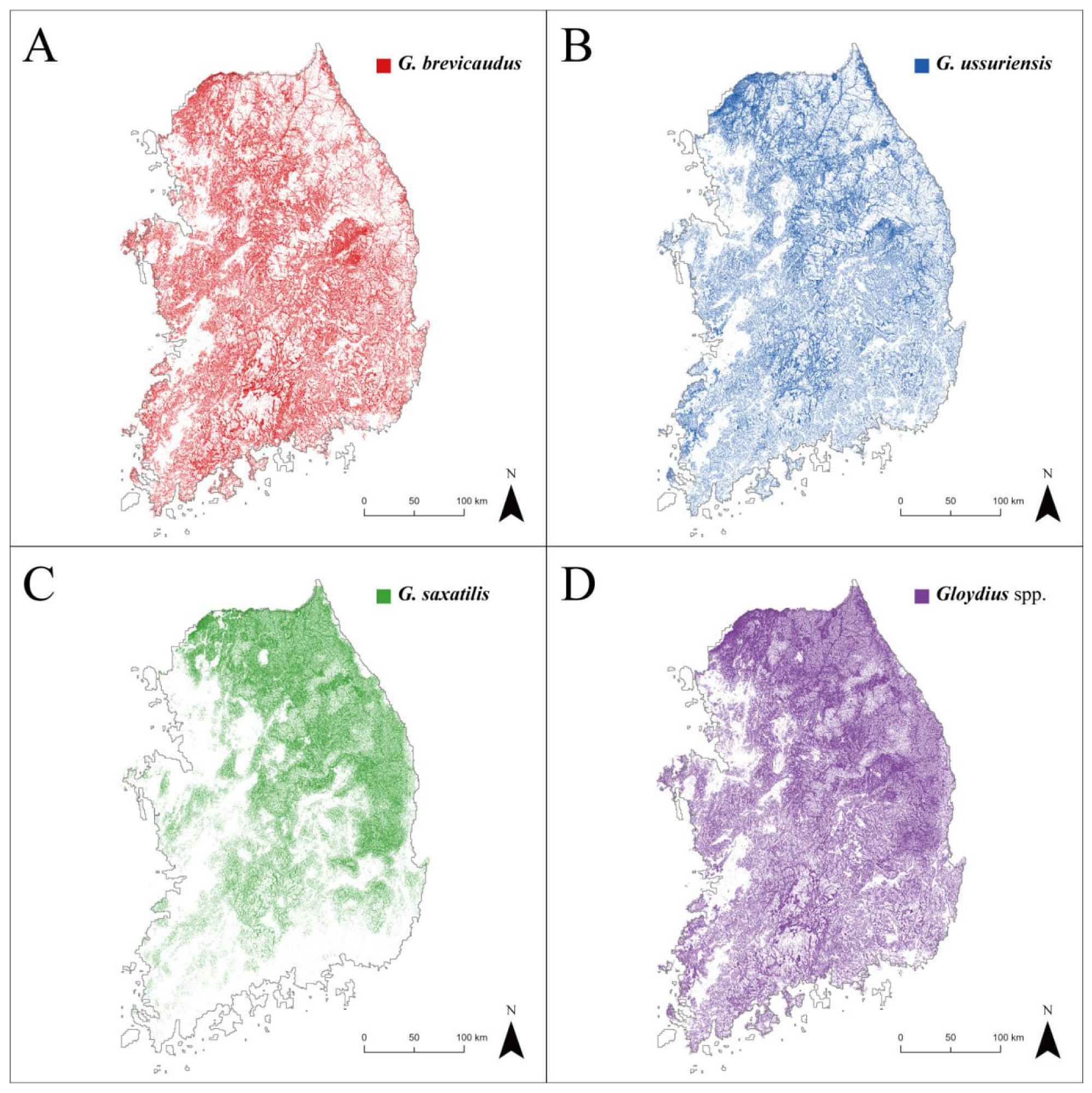
3.2. Habitat Distribution of Gloydius spp. and Its Relationship with Snakebite Accidents
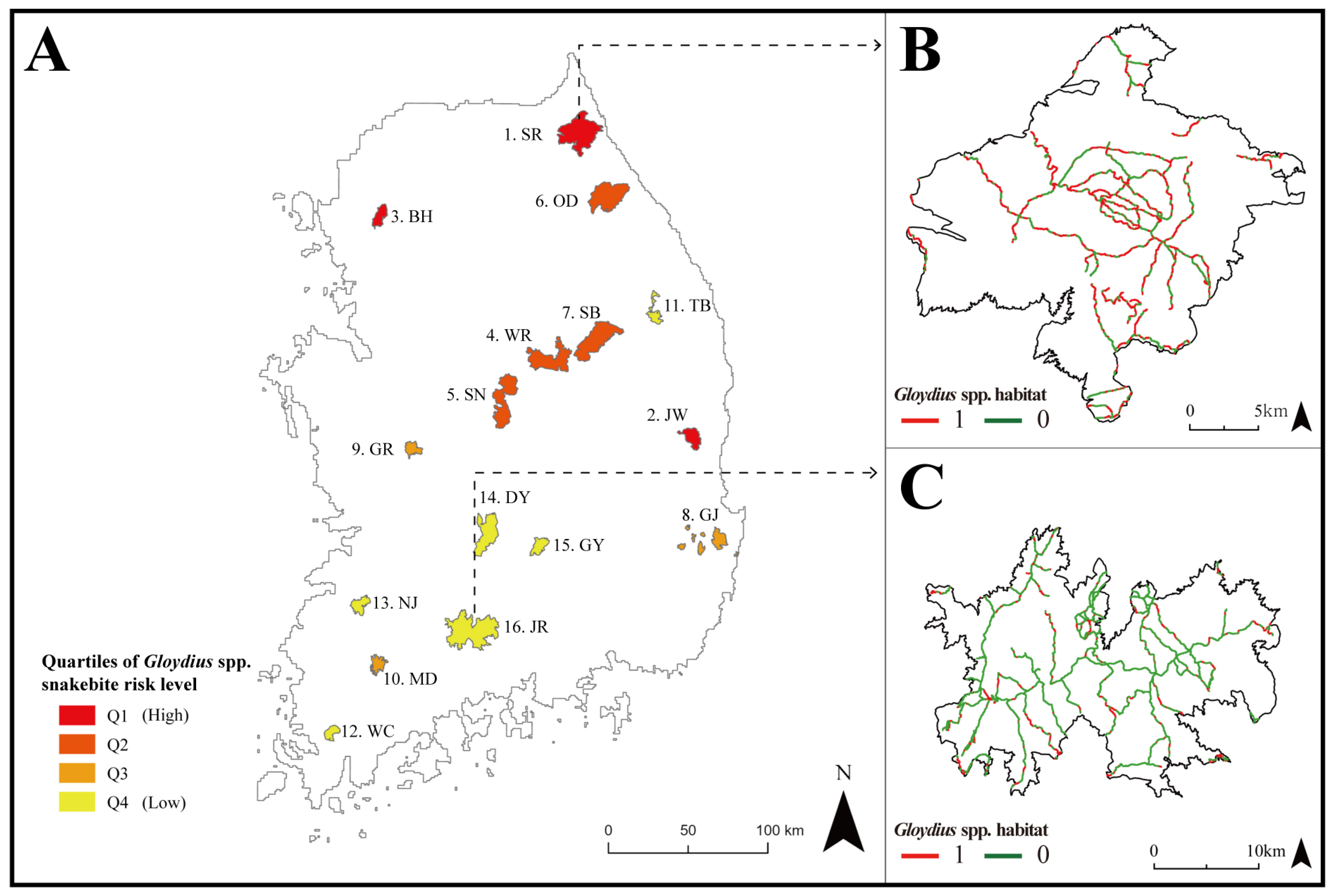
3.3. Snakebite Risk Assessment in National Parks
4. Discussion
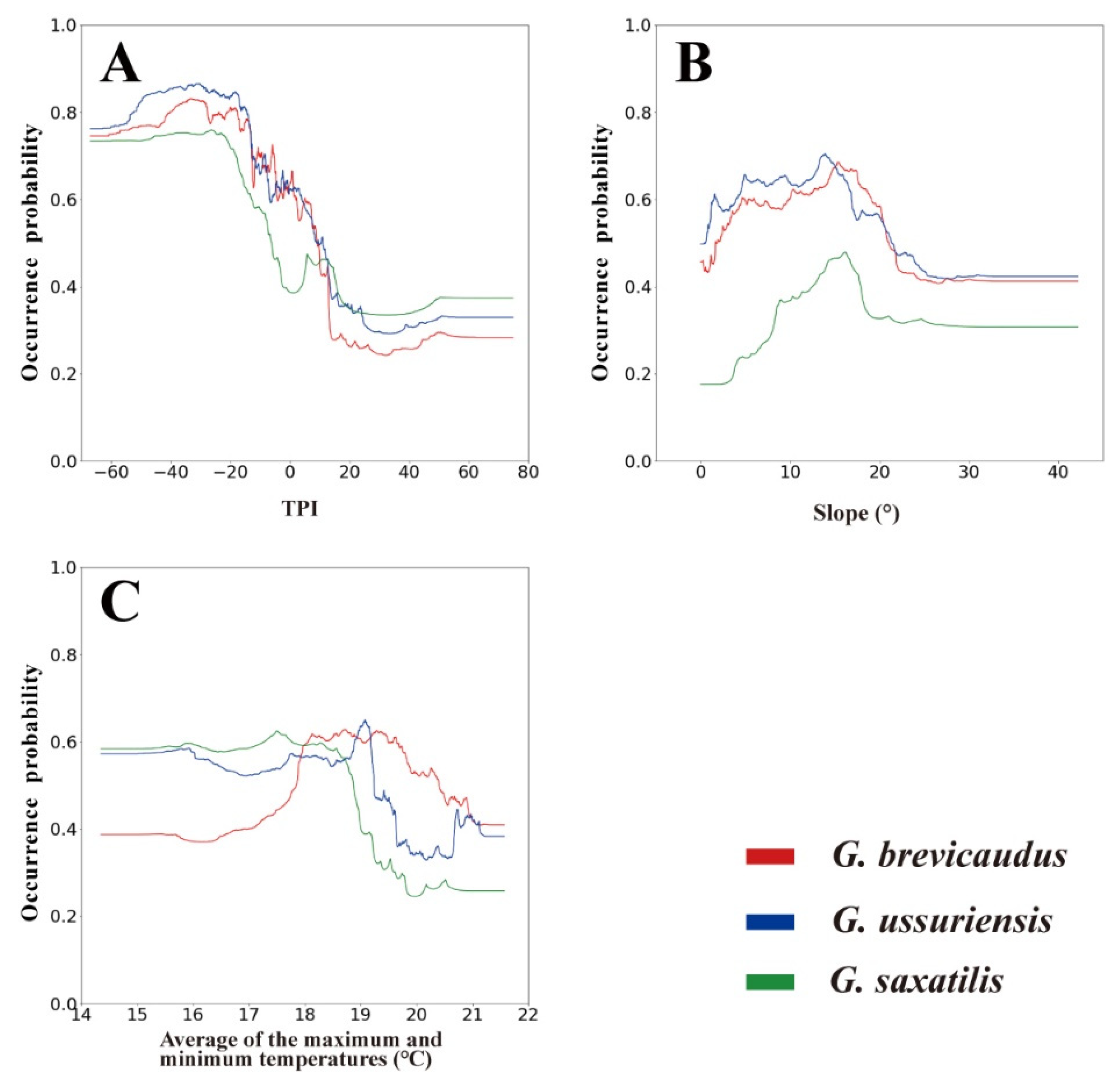
4.1. Evaluating the Importance of Environmental Variables
4.2. Snakebite Risk Assessment at a Regional Scale
4.3. Risk Assessment of Gloydius spp. in National Parks
4.4. Significance of the Research
4.5. Limitations of the Study and Future Research Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pop, M.I.; Gradinaru, S.R.; Popescu, V.D.; Haase, D.; Iojă, C.I. Emergency-line calls as an indicator to assess human–wildlife interaction in urban areas. Ecosphere 2023, 14, e4418. [Google Scholar] [CrossRef]
- Madden, F. Creating coexistence between humans and wildlife: Global perspectives on local efforts to address human–wildlife conflict. Hum. Dimens. Wildl. 2004, 9, 247–257. [Google Scholar] [CrossRef]
- Hamman, E.; Woolaston, K.; Lewis, B. Legal responses to human-wildlife conflict: The precautionary principle, risk analysis and the ‘lethal management’ of endangered species. IUCN Acad. Environ. Law ej 2016, 7, 57–83. [Google Scholar]
- Gouda, A.; Elnabarawy, N.; Badawy, S. A study of snakebite envenomation cases admitted to Egyptian national poisoning center. Acta Med. Int. 2017, 4, 34–40. [Google Scholar] [CrossRef]
- Boyd, J.J.; Agazzi, G.; Svajda, D.; Morgan, A.J.; Ferrandis, S.; Norris, R.L. Venomous snakebite in mountainous terrain: Prevention and management. Wilderness Environ. Med. 2007, 18, 190–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Regional Action Plan for Prevention and Control of Snakebite Envenoming in the South-East Asia 2022–2030. Available online: https://www.who.int/publications/i/item/9789290209829 (accessed on 19 April 2023).
- Kurniawan, N.; Fathoni, M.; Fatchiyah, F.; Septiadi, L.; Smith, E.N. Composition, distribution, and habitat type of snakes in Java, with discussion on the impact of human-snake interactions during 2013–2019. Herpetol. Notes 2021, 14, 691–711. [Google Scholar]
- Schneider, M.C.; Min, K.D.; Hamrick, P.N.; Montebello, L.R.; Ranieri, T.M.; Mardini, L.; Camara, V.M.; Raggio Luiz, R.; Liese, B.; Vuckovic, M.; et al. Overview of snakebite in Brazil: Possible drivers and a tool for risk mapping. PLoS Negl. Trop. Dis. 2021, 15, e0009044. [Google Scholar] [CrossRef] [PubMed]
- Warrell, D.A. Snake bite. Lancet 2010, 375, 77–88. [Google Scholar] [CrossRef]
- Kang, C.; Kim, D.H.; Kim, S.C.; Kim, D.S.; Jeong, C.Y. Atraumatic splenic rupture after coagulopathy owing to a snakebite. Wilderness Environ. Med. 2014, 25, 325–328. [Google Scholar] [CrossRef] [Green Version]
- Hifumi, T.; Sakai, A.; Kondo, Y.; Yamamoto, A.; Morine, N.; Ato, M.; Shibayama, K.; Umezawa, K.; Kiriu, N.; Kato, H.; et al. Venomous snake bites: Clinical diagnosis and treatment. J. Intensive Care 2015, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Ogura, Y.; Fukuchi, K.; Morimoto, H.; Yuki, T.; Otsuka, M.; Shimauchi, T.; Honda, T.; Tokura, Y. Elevation of circulating neutrophil extracellular traps, interleukin (IL)-8, IL-22, and vascular endothelial growth factor in patients with venomous snake mamushi (Gloydius blomhoffii) bites. J. Dermatol. 2022, 49, 124–132. [Google Scholar] [CrossRef]
- Orlov, N.L.; Sundukov, Y.N.; Kropachev, I.I. Distribution of pitvipers of “Gloydius blomhoffii” complex in Russia with the first records of Gloydius blomhoffii blomhoffii at Kunashir Island (Kuril Archipelago, Russian far east). Russ. J. Herpetol. 2014, 21, 169–178. Available online: https://www.researchgate.net/publication/279030602 (accessed on 19 April 2023).
- Kim, B.S.; Oh, H.S. Movement and home range of the red-tongued viper snake (Gloydius ussuriensis) inhabiting Gapado. Korean J. Environ. Ecol. 2015, 29, 192–199. [Google Scholar] [CrossRef]
- Goldingay, R.L.; Newell, D.A. Small-scale field experiments provide important insights to restore the rock habitat of Australia’s most endangered snake. Restor. Ecol. 2017, 25, 243–252. [Google Scholar] [CrossRef]
- Wasserberger, J.; Ordog, G.; Merkin, T.E. Southern Pacific rattlesnake bite: A unique clinical challenge. J. Emerg. Med. 2006, 31, 263–266. [Google Scholar] [CrossRef]
- Bogati, S.; Bhatt, N.; Nepal, S.; Nepali, P.; Sharma, S.K. Snakebite in the hills and mountains of Nepal. J. Nepal. Med. Assoc. 2022, 60, 1066–1069. [Google Scholar] [CrossRef] [PubMed]
- Merow, C.; Smith, M.J.; Edwards, T.C.; Guisan, A.; McMahon, S.M.; Normand, S.; Thuiller, W.; Wüest, R.O.; Zimmermann, N.E.; Elith, J. What do we gain from simplicity versus complexity in species distribution models? Ecography 2014, 37, 1267–1281. [Google Scholar] [CrossRef]
- Ananjeva, N.B.; Golynsky, E.E.; Lin, S.M.; Orlov, N.L.; Tseng, H.Y. Modeling habitat suitability to predict the potential distribution of the Kelung cat snake Boiga kraepelini Steineger. Russ. J. Herpetol. 2015, 22, 197–205. [Google Scholar]
- Archis, J.N.; Akcali, C.; Stuart, B.L.; Kikuchi, D.; Chunco, A.J. Is the future already here? The impact of climate change on the distribution of the eastern coral snake (Micrurus fulvius). PeerJ 2018, 6, e4647. [Google Scholar] [CrossRef] [Green Version]
- Kalboussi, M.; Achour, H. Modelling the spatial distribution of snake species in northwestern Tunisia using maximum entropy (Maxent) and Geographic Information System (GIS). J. For. Res. 2017, 29, 233–245. [Google Scholar] [CrossRef]
- Yanez-Arenas, C.; Peterson, A.T.; Mokondoko, P.; Rojas-Soto, O.; Martinez-Meyer, E. The use of ecological niche modeling to infer potential risk areas of snakebite in the Mexican state of Veracruz. PLoS ONE 2014, 9, e100957. [Google Scholar] [CrossRef] [Green Version]
- Melo Araujo, S.C.; Ceron, K.; Guedes, T.B. Use of geospatial analyses to address snakebite hotspots in mid-northern Brazil—A direction to health planning in shortfall biodiversity knowledge areas. Toxicon 2022, 213, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, M.; Kafash, A.; Khani, A.; Nabati, N. Applying species distribution models in public health research by predicting snakebite risk using venomous snakes’ habitat suitability as an indicating factor. Sci. Rep. 2020, 10, 18073. [Google Scholar] [CrossRef] [PubMed]
- Korea Meteorological Administration (KMA). Climate Statistics. Available online: https://data.kma.go.kr/ (accessed on 30 March 2023).
- Olaya-Marín, E.J.; Martínez-Capel, F.; Vezza, P. A comparison of artificial neural networks and random forests to predict native fish species richness in Mediterranean rivers. Knowl. Manag. Aquat. Ecosyst. 2013, 409, 07. [Google Scholar] [CrossRef] [Green Version]
- Do, M.S.; Lee, J.W.; Jang, H.J.; Kim, D.I.; Yoo, J.C. Interspecific competition and spatial ecology of three species of vipers in Korea: An application of ecological niche-based models and GIS. Korean J. Environ. Ecol. 2016, 30, 173–184. [Google Scholar] [CrossRef]
- Shakoori, S.; Todehdehghan, F.; Shiravi, A.; Hojati, V. The assessment of captive breeding in the Caucasian viper (Gloydius halys caucasicus) in Iran. J. Entomol. Zool. Stud. 2015, 3, 257–259. [Google Scholar]
- Fani, S.; Vaezi, Q.; Todehdehghan, F. Serum level of thyroid stimulating hormone, triiodothyronine, thyroxin and weight rate in Caucasian pit viper (Viperidae: Gloydius Halys Caucasicus) of Iran during different seasons. Annu. Res. Rev. Biol. 2017, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.; Jang, Y.; Borzee, A. Snakebite envenomings in the Republic of Korea from the 1970s to the 2020s: A review. Toxicon 2021, 196, 8–18. [Google Scholar] [CrossRef]
- Oh, H.S.; Banjade, M. Ecology of red-tongue viper (Gloydius ussuriensis) in Jeju Island, South Korea. In Snake Venom and Ecology, 1st ed.; Mohammad, M.S., Umar, S., Tijjani, R.B., Tijjani, S.I., Eds.; IntechOpen: London, UK, 2021; pp. 63–76. [Google Scholar]
- Lee, S.; Jeon, W. Epidemiology of patients with snake bite or envenomation in emergency department: NEDIS (national emergency department information system). J. Korean Soc. Clin. Toxicol. 2022, 20, 40–50. [Google Scholar] [CrossRef]
- Sawai, Y.; Honma, M.; Kawamura, Y.; Saki, A.; Hatsuse, M. Rhabdophis tigrinus in Japan: Pathogenesis of envenomation and production of antivenom. J. Toxicol. Toxin Rev. 2002, 21, 181–201. [Google Scholar] [CrossRef]
- Anita, S.; Hamidy, A.; Mulyadi; Mori, A. Effects of body size and condition on antipredator behavior related to nuchal glands in Rhabdophis subminiatus. J. Hepatol. 2022, 56, 454–460. [Google Scholar]
- Luiselli, L. Resource partitioning and interspecific competition in snakes: The search for general geographical and guild patterns. Oikos 2006, 114, 193–211. [Google Scholar] [CrossRef]
- Luiselli, L.; Filippi, E.; Di Lena, E. Ecological relationships between sympatric Vipera aspis and Vipera ursinii in high-altitude habitats of central Italy. J. Hepatol. 2007, 41, 378–384. [Google Scholar] [CrossRef]
- Do, M.S.; Yoo, J.C. Distribution pattern according to altitude and habitat type of the red-tongue viper snake (Gloydius ussuriensis) in the Cheon-ma mountain. J. Wet. Res. 2014, 16, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Do, M.S.; Shim, J.H.; Choi, Y.M.; Yoo, J.C. Effect of weight of radio-transmitters on survival of red-tongue viper snake (Gloydius ussuriensis) and short-tailed viper snake (Gloydius saxatilis) in the radio-transmitter implantation. J. Wet. Res. 2014, 16, 85–92. [Google Scholar] [CrossRef] [Green Version]
- De Reu, J.; Bourgeois, J.; Bats, M.; Zwertvaegher, A.; Gelorini, V.; De Smedt, P.; Chu, W.; Antrop, M.; De Maeyer, P.; Finke, P.; et al. Application of the topographic position index to heterogeneous landscapes. Geomorphology 2013, 186, 39–49. [Google Scholar] [CrossRef]
- Buchhorn, M.; Lesiv, M.; Tsendbazar, N.E.; Herold, M.; Bertels, L.; Smets, B. Copernicus global land cover layers—Collection 2. Remote Sens. 2020, 12, 1044. [Google Scholar] [CrossRef] [Green Version]
- Korea Forest Service (KFS). Hiking Trails in South Korea. Available online: https://www.forest.go.kr/ (accessed on 19 April 2023).
- Kim, H. Downloading the Latest South Korea’s Administrative Divisions (SHP). Available online: http://www.gisdeveloper.co.kr/ (accessed on 19 April 2023).
- Korean Statistical Information Service (KOSIS). Population by Administrative Region. Available online: https://kosis.kr/ (accessed on 19 April 2023).
- Health Insurance Review and Assessment Service (HIRA). The Top 100 of Disease Statistics in Life. Available online: https://www.hira.or.kr/ (accessed on 19 April 2023).
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Speiser, J.L.; Miller, M.E.; Tooze, J.; Ip, E. A comparison of random forest variable selection methods for classification prediction modeling. Expert. Syst. Appl. 2019, 134, 93–101. [Google Scholar] [CrossRef]
- Michel, C.L.; Pastore, J.H.; Bonnet, X. Impact of cool versus warm temperatures on gestation in the aspic viper (Vipera aspis). Comp. Biochem. Physiol. Part. A Mol. Integr. 2013, 165, 338–342. [Google Scholar] [CrossRef]
- Morisette, J.T.; Jarnevich, C.S.; Holcombe, T.R.; Talbert, C.B.; Ignizio, D.; Talbert, M.K.; Silva, C.; Koop, D.; Swanson, A.; Young, N.E. VisTrails SAHM: Visualization and workflow management for species habitat modeling. Ecography 2013, 36, 129–135. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Nicodemus, K.K. Letter to the editor: On the stability and ranking of predictors from random forest variable importance measures. Brief Bioinform. 2011, 12, 369–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, M.S.; Choi, S.; Jang, H.J.; Suh, J.H. Predicting the distribution of three Korean pit viper species (Gloydius brevicaudus, G. ussuriensis, and G. intermedius) under climate change. Russ. J. Herpetol. 2022, 29, 262–274. [Google Scholar] [CrossRef]
- Ding, L.; Gan, X.N.; He, S.P.; Zhao, E.M. A phylogeographic, demographic and historical analysis of the short-tailed pit viper (Gloydius brevicaudus): Evidence for early divergence and late expansion during the pleistocene. Mol. Ecol. 2011, 20, 1905–1922. [Google Scholar] [CrossRef] [PubMed]
- Kadota, Y. Is ovophis okinavensis active only in the cool season? Temporal foraging pattern of a subtropical pit viper in Okinawa, Japan. Zool. Stud. 2011, 50, 269–275. [Google Scholar]
- Korner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Lind, A.J.; Welsh, H.H., Jr. Ontogenetic changes in foraging behaviour and habitat use by the Oregon garter snake, Thamnophis atratus hydrophilus. Anim. Behav. 1994, 48, 1261–1273. [Google Scholar] [CrossRef] [Green Version]
- Heard, G.W.; Black, D.; Robertson, P. Habitat use by the inland carpet python (Morelia spilota metcalfei: Pythonidae): Seasonal relationships with habitat structure and prey distribution in a rural landscape. Austral Ecol. 2004, 29, 446–460. [Google Scholar] [CrossRef]
- Martínez-Freiría, F.; Sillero, N.; Lizana, M.; Brito, J.C. GIS-based niche models identify environmental correlates sustaining a contact zone between three species of European vipers. Divers. Distrib. 2008, 14, 452–461. [Google Scholar] [CrossRef]
- Schiebel, P.E.; Hubbard, A.M.; Goldman, D.I. Comparative study of snake lateral undulation kinematics in model heterogeneous terrain. Integr. Comp. Biol. 2020, icaa125. [Google Scholar] [CrossRef] [PubMed]
- Adavodi, R.; Khosravi, R.; Cushman, S.A.; Kaboli, M. Topographical features and forest cover influence landscape connectivity and gene flow of the Caucasian pit viper, Gloydius caucasicus (Nikolsky, 1916), in Iran. Landsc. Ecol. 2019, 34, 2615–2630. [Google Scholar] [CrossRef]
- Bravo-Vega, C.A.; Cordovez, J.M.; Renjifo-Ibanez, C.; Santos-Vega, M.; Sasa, M. Estimating snakebite incidence from mathematical models: A test in Costa Rica. PLoS Negl. Trop. Dis. 2019, 13, e0007914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korean Statistical Information Service (KOSIS). Number of Visitors to the National Park. Available online: https://kosis.kr (accessed on 19 April 2023).
- Obradović, S.; Tešin, A. Hiking in the COVID-19 era: Motivation and post-outbreak intentions. J. Sport Tour. 2022, 26, 147–164. [Google Scholar] [CrossRef]
- Martin, G.; Erinjery, J.J.; Ediriweera, D.; de Silva, H.J.; Lalloo, D.G.; Iwamura, T.; Murray, K.A. A mechanistic model of snakebite as a zoonosis: Envenoming incidence is driven by snake ecology, socioeconomics and its impacts on snakes. PLoS Negl. Trop. Dis. 2022, 16, e0009867. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Unit | Data Source (Reference) | |
|---|---|---|---|
| Topological | Digital elevation model (DEM) | m | USGS’s EarthExplorer (https://earthexplorer.usgs.gov) |
| Topographic position index (TPI) | - | Obtained from DEM | |
| Slope | ° (degree) | ||
| Land cover | Bare | % | Land Cover Viewer (https://lcviewer.vito.be/2015) |
| Herbaceous | |||
| Shrubland | |||
| Forest | |||
| Climatic | Average of the maximum and minimum temperatures | °C | WorldClim (https://www.worldclim.org) |
| Precipitation | mm | ||
| ID | Region | Human Population (Million) | Area (km2) | Number of Snakebites | Number of Snakebites per 10,000 People |
|---|---|---|---|---|---|
| 1 | Seoul | 9.93 | 605.24 | 78 | 0.08 |
| 2 | Busan | 3.50 | 732.46 | 28 | 0.08 |
| 3 | Daegu | 2.48 | 880.62 | 97 | 0.39 |
| 4 | Incheon | 2.94 | 350.81 | 45 | 0.15 |
| 5 | Gwangju | 1.47 | 498.01 | 73 | 0.50 |
| 6 | Daejeon | 1.51 | 539.16 | 85 | 0.56 |
| 7 | Ulsan | 1.17 | 1044.90 | 51 | 0.44 |
| 8 | Sejong-si | 0.24 | 464.86 | 14 | 0.58 |
| 9 | Gyeonggi-do | 12.71 | 10,045.13 | 528 | 0.42 |
| 10 | Gangwon-do | 1.55 | 16,590.84 | 316 | 2.04 |
| 11 | Chungcheongbuk-do | 1.59 | 7408.86 | 288 | 1.81 |
| 12 | Chungcheongnam-do | 2.10 | 8023.70 | 392 | 1.87 |
| 13 | Jeollabuk-do | 1.86 | 8024.87 | 267 | 1.43 |
| 14 | Jeollanam-do | 1.90 | 10,374.78 | 526 | 2.76 |
| 15 | Gyeongsangbuk-do | 2.70 | 18,901.51 | 569 | 2.11 |
| 16 | Gyeongsangnam-do | 3.37 | 9567.07 | 297 | 0.88 |
| Environmental Variable | G. brevicaudus | G. ussuriensis | G. saxatilis | Median |
|---|---|---|---|---|
| TPI | 76.25 | 184.75 | 10.90 | 76.25 |
| Slope | 53.17 | 115.58 | 7.30 | 53.17 |
| Average of the maximum and minimum temperatures | 48.12 | 116.11 | 10.46 | 48.12 |
| Precipitation | 43.93 | 88.76 | 6.04 | 43.93 |
| Forest cover | 43.80 | 87.36 | 4.39 | 43.80 |
| Herbaceous cover | 44.75 | 76.49 | 3.94 | 44.75 |
| Bare cover | 11.96 | 24.51 | 1.09 | 11.96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, Y.; Kim, C.; Yoon, S.; Kang, W. Identification of Risk Areas for Gloydius Snakebites in South Korea. Animals 2023, 13, 1959. https://doi.org/10.3390/ani13121959
Moon Y, Kim C, Yoon S, Kang W. Identification of Risk Areas for Gloydius Snakebites in South Korea. Animals. 2023; 13(12):1959. https://doi.org/10.3390/ani13121959
Chicago/Turabian StyleMoon, Youngjoo, Chaewan Kim, Sungsoo Yoon, and Wanmo Kang. 2023. "Identification of Risk Areas for Gloydius Snakebites in South Korea" Animals 13, no. 12: 1959. https://doi.org/10.3390/ani13121959
APA StyleMoon, Y., Kim, C., Yoon, S., & Kang, W. (2023). Identification of Risk Areas for Gloydius Snakebites in South Korea. Animals, 13(12), 1959. https://doi.org/10.3390/ani13121959