
Modeling the Wintering Habitat Distribution of the Black Stork in Shaanxi, China: A Hierarchical Integration of Climate and Land Use/Land Cover Data

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
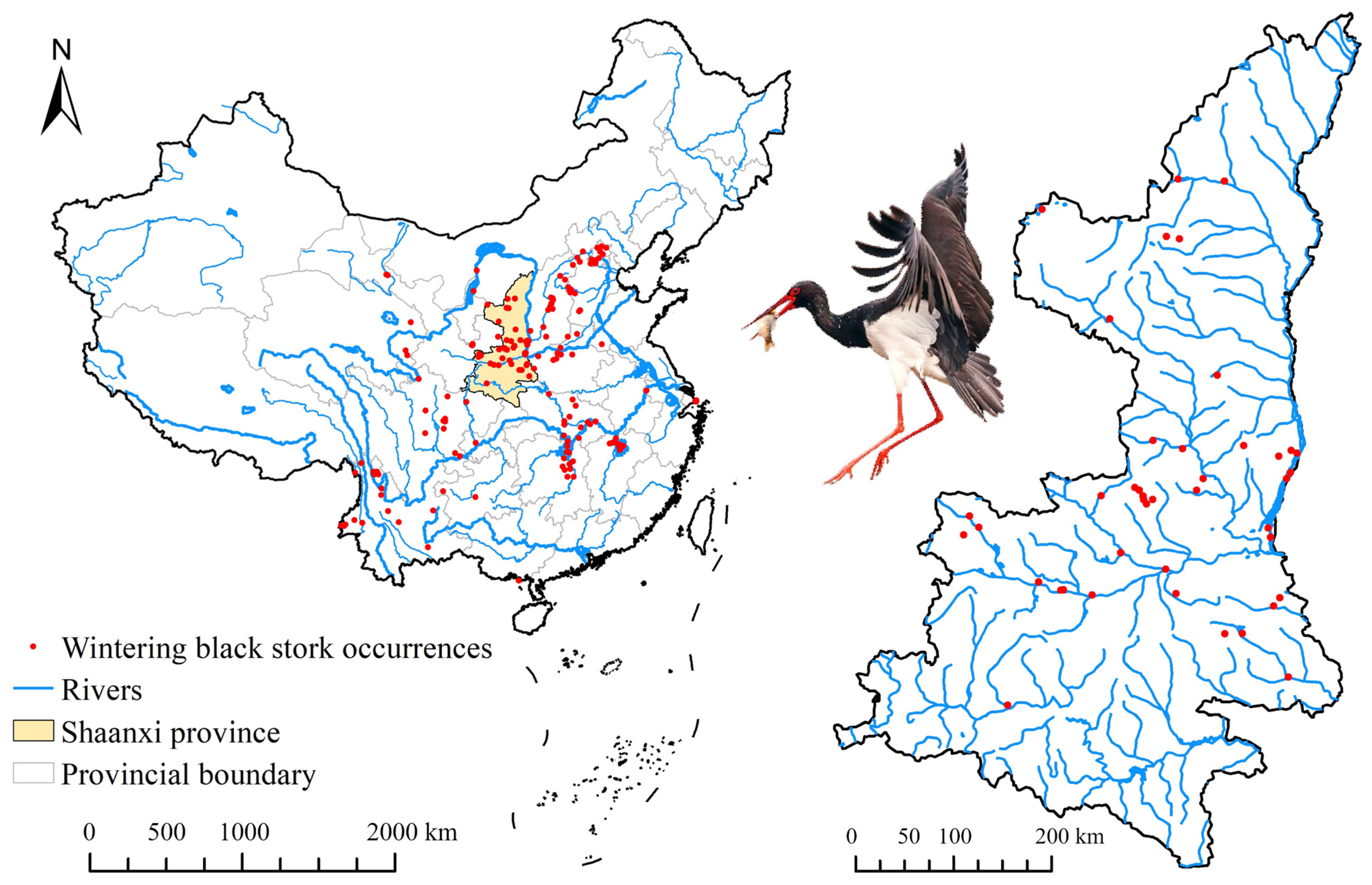
2.1. Species Occurrence Data and Scales
2.1.1. National Scale Database
2.1.2. Provincial Scale Database
2.2. Environmental Predictor Variables
2.3. LULC Simulation
2.4. Species Distribution Modelling
3. Results
3.1. Predicted LULC Changes
3.2. Major Environmental Factors That Affected Wintering Habitat Distribution
3.3. National and Provincial Scale SDMs for the Current Scenario
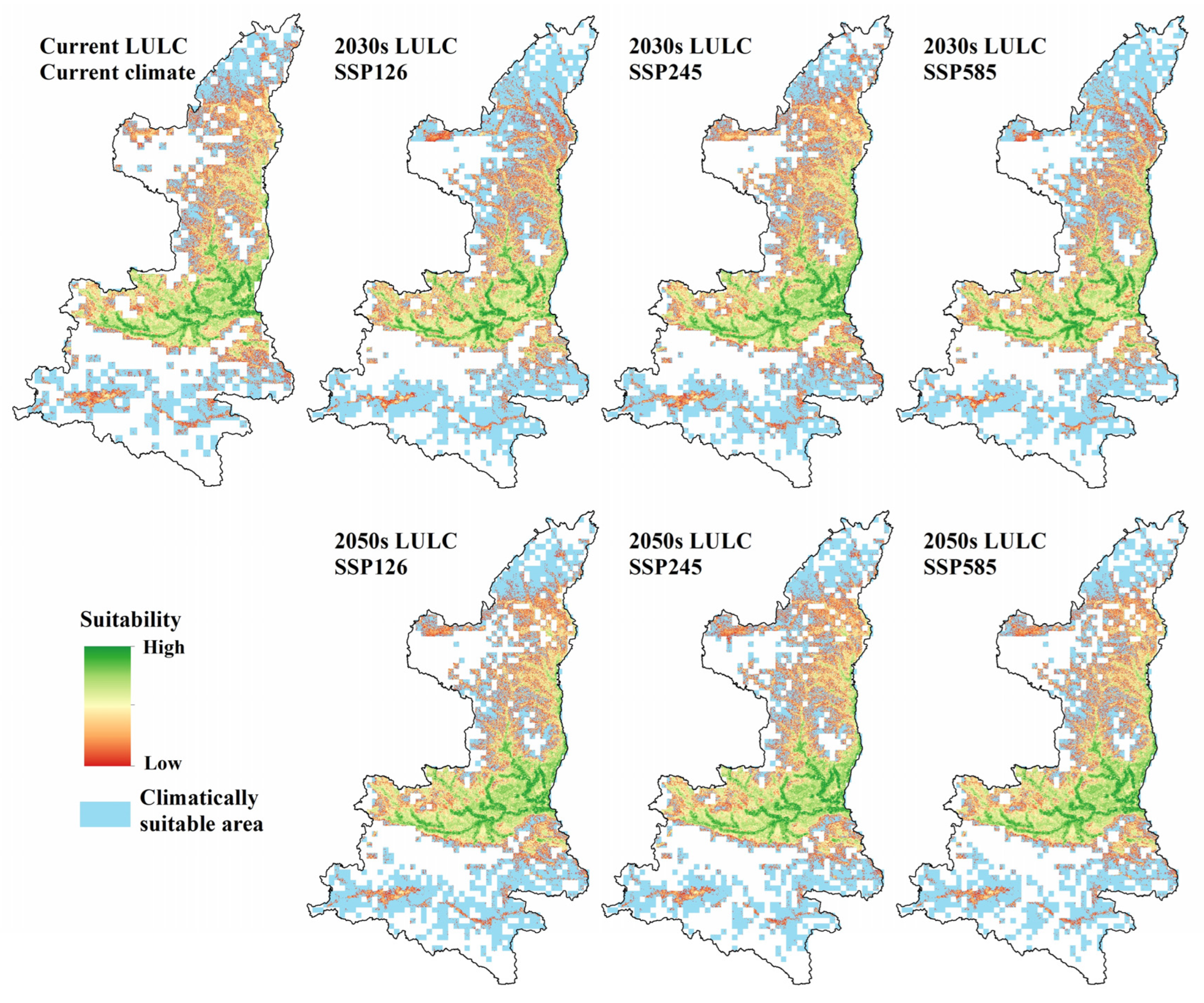
3.4. Projections of Future Scenarios
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J.; Li, D.; Xue, Y.; Wu, B.; He, X.; Liu, F. Identifying potential refugia and corridors under climate change: A case study of endangered Sichuan golden monkey (Rhinopithecus roxellana) in Qinling Mountains, China. Am. J. Primatol. 2018, 80, e22929. [Google Scholar] [CrossRef]
- Li, J.; Liu, F.; Xue, Y.; Zhang, Y.; Li, D. Assessing vulnerability of giant pandas to climate change in the Qinling Mountains of China. Ecol. Evol. 2017, 7, 4003–4015. [Google Scholar] [CrossRef]
- Mateo, R.G.; Gastón, A.; Aroca-Fernández, M.J.; Broennimann, O.; Guisan, A.; Saura, S.; García-Viñas, J.I. Hierarchical species distribution models in support of vegetation conservation at the landscape scale. J. Veg. Sci. 2019, 30, 386–396. [Google Scholar] [CrossRef]
- Pang, S.E.H.; De Alban, J.D.T.; Webb, E.L. Effects of climate change and land cover on the distributions of a critical tree family in the Philippines. Sci. Rep. 2021, 11, 276. [Google Scholar] [CrossRef] [PubMed]
- del Río, S.; Canas, R.; Cano, E.; Cano-Ortiz, A.; Musarella, C.; Pinto-Gomes, C.; Penas, A. Modelling the impacts of climate change on habitat suitability and vulnerability in deciduous forests in Spain. Ecol. Indic. 2021, 131, 108202. [Google Scholar] [CrossRef]
- Elsen, P.R.; Monahan, W.B.; Merenlender, A.M. Topography and human pressure in mountain ranges alter expected species responses to climate change. Nat. Commun. 2020, 11, 1974. [Google Scholar] [CrossRef]
- Liu, L.; Liao, J.; Wu, Y.; Zhang, Y. Breeding range shift of the red-crowned crane (Grus japonensis) under climate change. PLoS ONE 2020, 15, e0229984. [Google Scholar] [CrossRef] [PubMed]
- Virkkala, R.; Heikkinen, R.K.; Fronzek, S.; Leikola, N. Climate change, northern birds of conservation concern and matching the hotspots of habitat suitability with the reserve network. PLoS ONE 2013, 8, e63376. [Google Scholar] [CrossRef]
- Zhao, Q.; Boomer, G.S.; Royle, J.A. Integrated modeling predicts shifts in waterbird population dynamics under climate change. Ecography 2019, 42, 1470–1481. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P.; Liu, C. Modelling species distributions in Britain: A hierarchical integration of climate and land-cover data. Ecography 2004, 27, 285–298. [Google Scholar] [CrossRef]
- Wang, C.; Wang, G.; Guo, Z.; Dai, L.; Liu, H.; Li, Y.; Chen, H.; Zhao, Y.; Zhang, Y.; Cheng, H. Effects of land-use change on the distribution of the wintering red-crowned crane (Grus japonensis) in the coastal area of northern Jiangsu Province, China. Land Use Policy 2020, 90, 104269. [Google Scholar] [CrossRef]
- Ye, X.; Yu, X.; Yu, C.; Tayibazhaer, A.; Xu, F.; Skidmore, A.K.; Wang, T. Impacts of future climate and land cover changes on threatened mammals in the semi-arid Chinese Altai Mountains. Sci. Total Environ. 2018, 612, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, M.; Zarzo-Arias, A.; Guélat, J.; Mateo, R.G.; Guisan, A. Accounting for niche truncation to improve spatial and temporal predictions of species distributions. Front. Ecol. Evol. 2022, 10, 944116. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Bellamy, C.; Boughey, K.; Hawkins, C.; Reveley, S.; Spake, R.; Williams, C.; Altringham, J. A sequential multi-level framework to improve habitat suitability modelling. Landsc. Ecol. 2020, 35, 1001–1020. [Google Scholar] [CrossRef]
- Carrillo-Garcia, C.; Girola-Iglesias, L.; Guijarro, M.; Hernando, C.; Madrigal, J.; Mateo, R.G. Ecological niche models applied to post-megafire vegetation restoration in the context of climate change. Sci. Total Environ. 2022, 855, 158858. [Google Scholar] [CrossRef]
- Mateo, R.G.; Aroca-Fernández, M.J.; Gastón, A.; Gómez-Rubio, V.; Saura, S.; García-Viñas, J.I. Looking for an optimal hierarchical approach for ecologically meaningful niche modelling. Ecol. Model. 2019, 409, 108735. [Google Scholar] [CrossRef]
- Petitpierre, B.; McDougall, K.; Seipel, T.; Broennimann, O.; Guisan, A.; Kueffer, C. Will climate change increase the risk of plant invasions into mountains? Ecol. Appl. 2016, 26, 530–544. [Google Scholar] [CrossRef]
- Chevallier, D.; Le Maho, Y.; Brossault, P.; Baillon, F.; Massemin, S. The use of stopover sites by Black Storks (Ciconia nigra) migrating between West Europe and West Africa as revealed by satellite telemetry. J. Ornithol. 2010, 152, 1–13. [Google Scholar] [CrossRef]
- Liang, W.-L.; Zhu, J.; Ren, Z.-M. Complete mitochondrial genome of Ciconia nigra (Ciconiiformes: Ciconiidae), a threatened stork in China. Mitochondrial DNA Part B 2019, 4, 2509–2510. [Google Scholar] [CrossRef]
- Knudsen, E.; Linden, A.; Both, C.; Jonzen, N.; Pulido, F.; Saino, N.; Sutherland, W.J.; Bach, L.A.; Coppack, T.; Ergon, T.; et al. Challenging claims in the study of migratory birds and climate change. Biol. Rev. Camb. Philos. Soc. 2011, 86, 928–946. [Google Scholar] [CrossRef] [PubMed]
- Telleria, J.L.; Fernandez-Lopez, J.; Fandos, G. Effect of Climate Change on Mediterranean Winter Ranges of Two Migratory Passerines. PLoS ONE 2016, 11, e0146958. [Google Scholar] [CrossRef]
- Sanderson, F.J.; Donald, P.F.; Pain, D.J.; Burfield, I.J.; van Bommel, F.P.J. Long-term population declines in Afro-Palearctic migrant birds. Biol. Conserv. 2006, 131, 93–105. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Walther, B.A.; Thuiller, W.; Rahbek, C.; Jiguet, F. Potential impacts of climate change on the winter distribution of Afro-Palaearctic migrant passerines. Biol. Lett. 2009, 5, 248–251. [Google Scholar] [CrossRef]
- Mateo, R.G.; Gastón, A.; Aroca-Fernández, M.J.; Saura, S.; García-Viñas, J.I. Optimization of forest sampling strategies for woody plant species distribution modelling at the landscape scale. For. Ecol. Manag. 2018, 410, 104–113. [Google Scholar] [CrossRef]
- El-Gabbas, A.; Dormann, C.F. Wrong, but useful: Regional species distribution models may not be improved by range-wide data under biased sampling. Ecol. Evol. 2018, 8, 2196–2206. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P.; Araújo, M. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Aghainajafi-Zadeh, S.; Hemami, M.-R.; Heydari, F. Relationship between Food Resources and Brooding Site by Asiatic Houbara (Chlamydotis macqueenii) in Central Steppe of Iran S. World Acad. Sci. Eng. Technol. 2011, 53, 558–560. [Google Scholar]
- Yousefi, M.; Ahmadi, M.; Nourani, E.; Rezaei, A.L.I.; Kafash, A.; Khani, A.L.I.; Sehhatisabet, M.E.; Adibi, M.A.; Goudarzi, F.; Kaboli, M. Habitat suitability and impacts of climate change on the distribution of wintering population of Asian Houbara Bustard Chlamydotis macqueenii in Iran. Bird Conserv. Int. 2016, 27, 294–304. [Google Scholar] [CrossRef]
- Eyring, V.; Bony, S.; Meehl, G.A.; Senior, C.A.; Stevens, B.; Stouffer, R.J.; Taylor, K.E. Overview of the Coupled Model Intercomparison Project Phase 6 (CMIP6) experimental design and organization. Geosci. Model Dev. 2016, 9, 1937–1958. [Google Scholar] [CrossRef]
- Meinshausen, M.; Nicholls, Z.R.J.; Lewis, J.; Gidden, M.J.; Vogel, E.; Freund, M.; Beyerle, U.; Gessner, C.; Nauels, A.; Bauer, N.; et al. The shared socio-economic pathway (SSP) greenhouse gas concentrations and their extensions to 2500. Geosci. Model Dev. 2020, 13, 3571–3605. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Computer Software. 2019. Available online: https://www.r-project.org/ (accessed on 5 April 2021).
- Liang, X.; Guan, Q.; Clarke, K.C.; Liu, S.; Wang, B.; Yao, Y. Understanding the drivers of sustainable land expansion using a patch-generating land use simulation (PLUS) model: A case study in Wuhan, China. Comput. Environ. Urban Syst. 2021, 85, 101569. [Google Scholar] [CrossRef]
- Zhai, H.; Lv, C.; Liu, W.; Yang, C.; Fan, D.; Wang, Z.; Guan, Q. Understanding Spatio-Temporal Patterns of Land Use/Land Cover Change under Urbanization in Wuhan, China, 2000–2019. Remote Sens. 2021, 13, 3331. [Google Scholar] [CrossRef]
- Kvalseth, T.O. A Coefficient of Agreement for Nominal Scales: An Asymmetric Version of Kappa. Educ. Psychol. Meas. 1991, 51, 95–101. [Google Scholar] [CrossRef]
- Andrade, A.F.A.d.; Velazco, S.J.E.; De Marco Júnior, P. ENMTML: An R package for a straightforward construction of complex ecological niche models. Environ. Model. Softw. 2020, 125, 104615. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Cohen, J. A Coefficient of Agreement for Nominal Scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Leroy, B.; Delsol, R.; Hugueny, B.; Meynard, C.N.; Barhoumi, C.; Barbet-Massin, M.; Bellard, C. Without quality presence–absence data, discrimination metrics such as TSS can be misleading measures of model performance. J. Biogeogr. 2018, 45, 1994–2002. [Google Scholar] [CrossRef]
- Li, W.; Guo, Q. How to assess the prediction accuracy of species presence–absence models without absence data? Ecography 2013, 36, 788–799. [Google Scholar] [CrossRef]
- Boyce, M.S.; Vernier, P.R.; Nielsen, S.E.; Schmiegelow, F.K.A. Evaluating resource selection functions. Ecol. Model. 2002, 157, 281–300. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Dai, Y.; Hacker, C.E.; Zhang, Y.; Li, W.; Zhang, Y.; Liu, H.; Zhang, J.; Ji, Y.; Xue, Y.; Li, D. Identifying climate refugia and its potential impact on Tibetan brown bear (Ursus arctos pruinosus) in Sanjiangyuan National Park, China. Ecol. Evol. 2019, 9, 13278–13293. [Google Scholar] [CrossRef] [PubMed]
- Cano, L.S.; Pacheco, C.; Refoyo, P.; Tellería, J.L. Geographical and environmental factors affecting the distribution of wintering black storks Ciconia nigra in the Iberian Peninsula. J. Avian Biol. 2014, 45, 514–521. [Google Scholar] [CrossRef]
- Ramachandran, R.M.; Roy, P.S.; Chakravarthi, V.; Joshi, P.K.; Sanjay, J. Land use and climate change impacts on distribution of plant species of conservation value in Eastern Ghats, India: A simulation study. Environ. Monit. Assess. 2020, 192, 86. [Google Scholar] [CrossRef]
- Thuiller, W.; Brotons, L.; Araújo, M.B.; Lavorel, S. Effects of restricting environmental range of data to project current and future species distributions. Ecography 2004, 27, 165–172. [Google Scholar] [CrossRef]
- Sánchez-Fernández, D.; Lobo, J.M.; Hernández-Manrique, O.L. Species distribution models that do not incorporate global data misrepresent potential distributions: A case study using Iberian diving beetles. Divers. Distrib. 2011, 17, 163–171. [Google Scholar] [CrossRef]
- Fan, L.; Wei, Z.; Tong, W. The Characteristic of Temporal and Spatial Distribution of the Differences between Ground and Air Temperature in the Arid Region of Northwest China. Plateau Weather. 2004, 23, 360–367. (In Chinese) [Google Scholar]
- Qiao, l.; Du, J.; Li, Q.; Wu, L. Study on the applicability of drought indicators in Shaanxi Province. Agric. Res. Arid. Zones 2010, 28, 002. (In Chinese) [Google Scholar]
- Amaral, A.G.; Munhoz, C.B.R.; Walter, B.M.T.; Aguirre-Gutiérrez, J.; Raes, N. Richness pattern and phytogeography of the Cerrado herb–shrub flora and implications for conservation. J. Veg. Sci. 2017, 28, 848–858. [Google Scholar] [CrossRef]
- Deb, J.C.; Phinn, S.; Butt, N.; McAlpine, C.A. The impact of climate change on the distribution of two threatened Dipterocarp trees. Ecol. Evol. 2017, 7, 2238–2248. [Google Scholar] [CrossRef]
- Kanagaraj, R.; Araujo, M.B.; Barman, R.; Davidar, P.; De, R.; Digal, D.K.; Gopi, G.V.; Johnsingh, A.J.T.; Kakati, K.; Kramer-Schadt, S.; et al. Predicting range shifts of Asian elephants under global change. Divers. Distrib. 2019, 25, 822–838. [Google Scholar] [CrossRef]
- Peterson, A.T.; Cobos, M.E.; Jiménez-García, D. Major challenges for correlational ecological niche model projections to future climate conditions. Ann. N. Y. Acad. Sci. 2018, 1429, 66–77. [Google Scholar] [CrossRef]
- Scheele, B.C.; Foster, C.N.; Banks, S.C.; Lindenmayer, D.B. Niche Contractions in Declining Species: Mechanisms and Consequences. Trends Ecol. Evol. 2017, 32, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Mair, L.; Jönsson, M.; Räty, M.; Bärring, L.; Strandberg, G.; Lämås, T.; Snäll, T. Land use changes could modify future negative effects of climate change on old-growth forest indicator species. Divers. Distrib. 2018, 24, 1416–1425. [Google Scholar] [CrossRef]
- Methorst, J.; Böhning-Gaese, K.; Khaliq, I.; Hof, C. A framework integrating physiology, dispersal and land-use to project species ranges under climate change. J. Avian Biol. 2017, 48, 1532–1548. [Google Scholar] [CrossRef]
- Titeux, N.; Henle, K.; Mihoub, J.-B.; Regos, A.; Geijzendorffer, I.R.; Cramer, W.; Verburg, P.H.; Brotons, L. Biodiversity scenarios neglect future land-use changes. Glob. Chang. Biol. 2016, 22, 2505–2515. [Google Scholar] [CrossRef]
- Chen, X.; Yu, L.; Cao, Y.; Xu, Y.; Zhao, Z.; Zhuang, Y.; Liu, X.; Du, Z.; Liu, T.; Yang, B.; et al. Habitat quality dynamics in China’s first group of national parks in recent four decades: Evidence from land use and land cover changes. J. Environ. Manag. 2023, 325, 116505. [Google Scholar] [CrossRef]
- Coppée, T.; Paquet, J.-Y.; Titeux, N.; Dufrêne, M. Temporal transferability of species abundance models to study the changes of breeding bird species based on land cover changes. Ecol. Model. 2022, 473, 110136. [Google Scholar] [CrossRef]
- Faleiro, F.V.; Machado, R.B.; Loyola, R.D. Defining spatial conservation priorities in the face of land-use and climate change. Biol. Conserv. 2013, 158, 248–257. [Google Scholar] [CrossRef]
- Oswald, J.S.; Joshua, J.L.; Paul, B.; Craig, G.; Gary, K.; Douglas, A.B.; Jason, B.; Kevin, M.J.; Mary, L.K.; Kit, M.; et al. Conserving Biodiversity: Practical Guidance about Climate Change Adaptation Approaches in Support of Land-use Planning. Nat. Areas J. 2015, 35, 190–203. [Google Scholar] [CrossRef]
- Mantyka-Pringle, C.S.; Visconti, P.; Di Marco, M.; Martin, T.G.; Rondinini, C.; Rhodes, J.R. Climate change modifies risk of global biodiversity loss due to land-cover change. Biol. Conserv. 2015, 187, 103–111. [Google Scholar] [CrossRef]
- Ma, Z.; Chen, Y.; Melville, D.S.; Fan, J.; Liu, J.; Dong, J.; Tan, K.; Cheng, X.; Fuller, R.A.; Xiao, X.; et al. Changes in area and number of nature reserves in China. Conserv. Biol. 2019, 33, 1066–1075. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year 2020 | Year 2030 | Change in 2020–2030 (%) | Year 2050 | Change in 2020–2050 (%) | |
|---|---|---|---|---|---|
| Cropland | 59,858.55 | 58,964.04 | −1.53 | 62,569.8 | 4.53 |
| Forest | 83,945.79 | 84,545.82 | 0.71 | 92,049.57 | 9.65 |
| Grassland | 42,187.5 | 41,398.74 | −1.87 | 42,259.5 | 0.17 |
| Wetland | 1018.08 | 1025.28 | 0.71 | 1160.19 | 13.95 |
| Impervious | 3074.76 | 4141.26 | 34.69 | 6588.27 | 114.26 |
| Bare land | 1436.04 | 1448.19 | 0.85 | 1400.67 | −2.46 |
| Scale | Metrics | |||||
|---|---|---|---|---|---|---|
| AUC | Kappa | TSS | Jaccard | Sorensen | Boyce | |
| National | 0.91 | 0.69 | 0.69 | 0.75 | 0.85 | 0.96 |
| Provincial | 0.93 | 0.78 | 0.78 | 0.82 | 0.90 | 0.73 |
| Scale | Variable | Algorithms | μ (Mean) | |||||
|---|---|---|---|---|---|---|---|---|
| MXD | GLM | GAM | RDF | SVM | BRT | |||
| National | LULC | 0.13 | 0.30 | 0.14 | 0.15 | 0.04 | 0.11 | 0.14 |
| Bio15 | 0.17 | 0.16 | 0.18 | 0.18 | 0.16 | 0.12 | 0.16 | |
| Bio3 | 0.03 | 0.07 | 0.09 | 0.18 | 0.04 | 0.14 | 0.09 | |
| Bio14 | 0.27 | 0.15 | 0.22 | 0.20 | 0.31 | 0.14 | 0.21 | |
| Bio9 | 0.40 | 0.33 | 0.37 | 0.29 | 0.46 | 0.49 | 0.39 | |
| Provincial | Aspect | 0.01 | 0.04 | 0.01 | 0.07 | 0.02 | 0.03 | 0.03 |
| Elevation | 0.07 | 0.07 | 0.13 | 0.14 | 0.09 | 0.06 | 0.09 | |
| LULC | 0.14 | 0.26 | 0.18 | 0.15 | 0.14 | 0.14 | 0.17 | |
| Nightlight | 0.29 | 0.04 | 0.04 | 0.13 | 0.16 | 0.08 | 0.13 | |
| Dist_road | 0.11 | 0.15 | 0.14 | 0.13 | 0.12 | 0.05 | 0.12 | |
| Slope | 0.19 | 0.14 | 0.05 | 0.18 | 0.20 | 0.14 | 0.15 | |
| Dist_water | 0.19 | 0.30 | 0.47 | 0.21 | 0.27 | 0.50 | 0.32 | |
| National Model | Provincial Model | |||||
|---|---|---|---|---|---|---|
| Current | 2030s | 2050s | Current | 2030s | 2050s | |
| LULC change only | 96,333 | 85,006 (−11.7%) | 79,698 (−17.3%) | |||
| Baseline condition | 127,056 | 77,939 | ||||
| SSP126 | 136,800 (7.6%) | 135,800 (6.9%) | 69,944 (−10.25%) | 74,645 (−4.23%) | ||
| SSP245 | 138,300 (8.8%) | 135,400 (6.6%) | 75,749 (−2.80%) | 78,588 (0.83) | ||
| SSP585 | 133,200 (4.8%) | 133,800 (5.3%) | 68,983 (−11.49%) | 74,709 (−4.14%) | ||
| 2030s | 2050s | |||||
|---|---|---|---|---|---|---|
| Stable | Gain | Lost | Stable | Gain | Lost | |
| SSP126 | 61,189 | 8754 | 16,749 | 66,432 | 8213 | 11,506 |
| SSP245 | 73,322 | 10,265 | 4615 | 67,030 | 8718 | 10,908 |
| SSP585 | 60,619 | 8363 | 17,319 | 66,408 | 8300 | 11,529 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuohetahong, Y.; Lu, R.; Gan, F.; Li, M.; Ye, X.; Yu, X. Modeling the Wintering Habitat Distribution of the Black Stork in Shaanxi, China: A Hierarchical Integration of Climate and Land Use/Land Cover Data. Animals 2023, 13, 2726. https://doi.org/10.3390/ani13172726
Tuohetahong Y, Lu R, Gan F, Li M, Ye X, Yu X. Modeling the Wintering Habitat Distribution of the Black Stork in Shaanxi, China: A Hierarchical Integration of Climate and Land Use/Land Cover Data. Animals. 2023; 13(17):2726. https://doi.org/10.3390/ani13172726
Chicago/Turabian StyleTuohetahong, Yilamujiang, Ruyue Lu, Feng Gan, Min Li, Xinping Ye, and Xiaoping Yu. 2023. "Modeling the Wintering Habitat Distribution of the Black Stork in Shaanxi, China: A Hierarchical Integration of Climate and Land Use/Land Cover Data" Animals 13, no. 17: 2726. https://doi.org/10.3390/ani13172726
APA StyleTuohetahong, Y., Lu, R., Gan, F., Li, M., Ye, X., & Yu, X. (2023). Modeling the Wintering Habitat Distribution of the Black Stork in Shaanxi, China: A Hierarchical Integration of Climate and Land Use/Land Cover Data. Animals, 13(17), 2726. https://doi.org/10.3390/ani13172726