
Two Years after Coxiella burnetii Detection: Pathogen Shedding and Phase-Specific Antibody Response in Three Dairy Goat Herds
 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Herd History
2.1.1. Dairy Goat Herd A
2.1.2. Dairy Goat Herd B
2.1.3. Dairy Goat Herd C
2.2. Collection of Blood Samples and Vaginal Swabs
2.3. Collection of Environmental Dust and Bulk Tank Milk
2.4. Laboratory Analysis of Sera, Vaginal Swabs, Dust Swabs, and Bulk Tank Milk
2.5. Collection and Analysis of Abortion Material
2.6. Statistical Analysis
3. Results
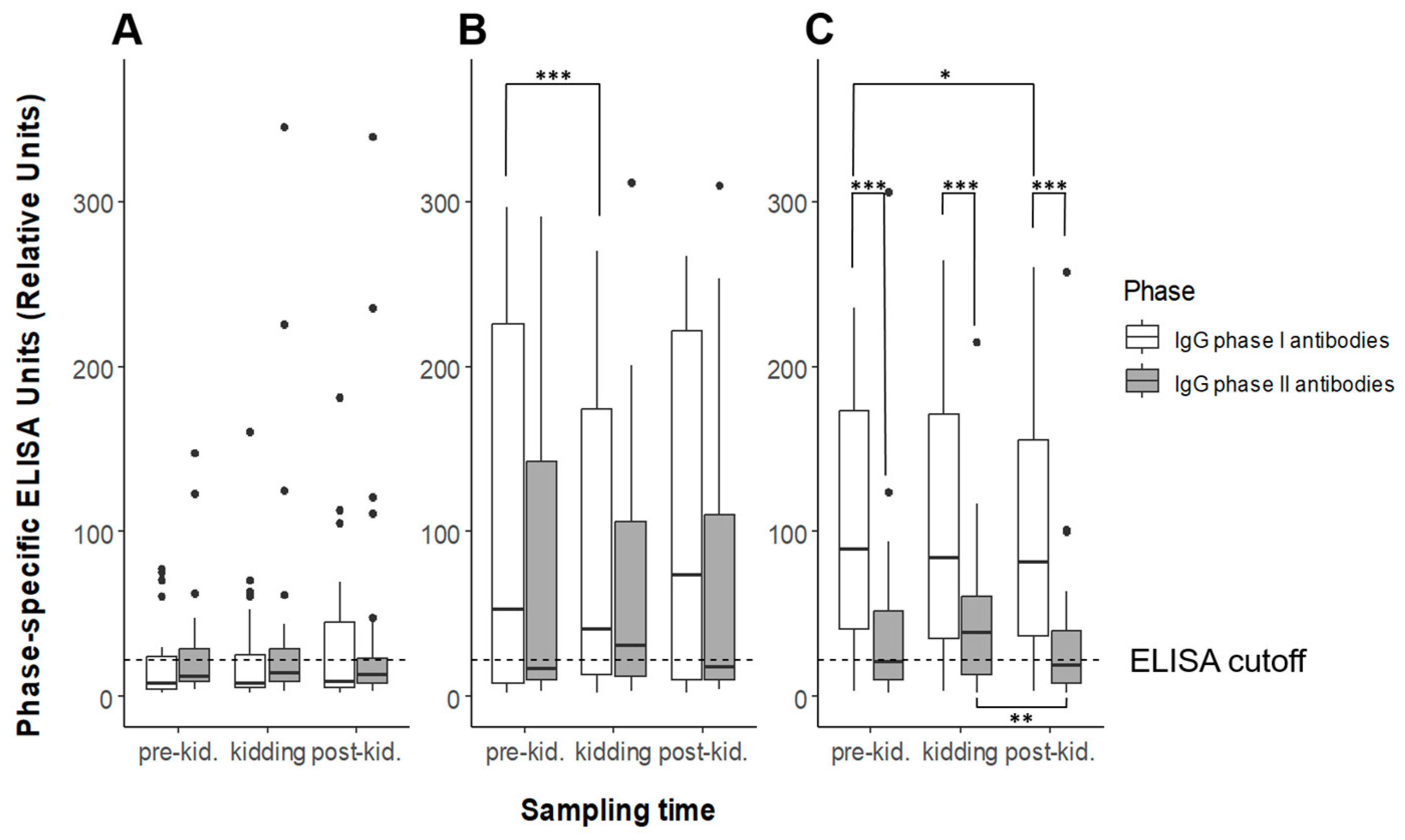
3.1. Serology
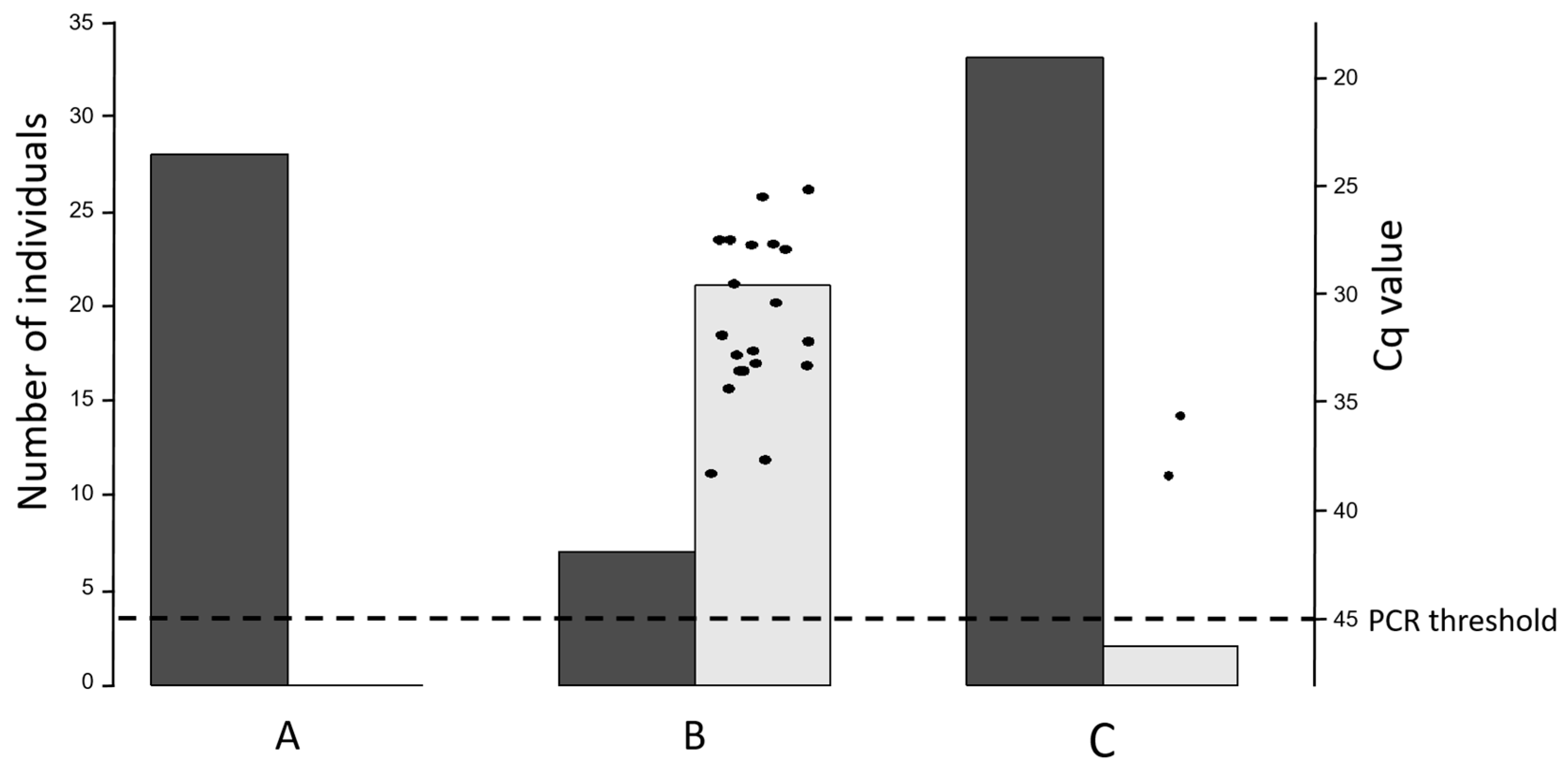
3.2. Vaginal Swabs
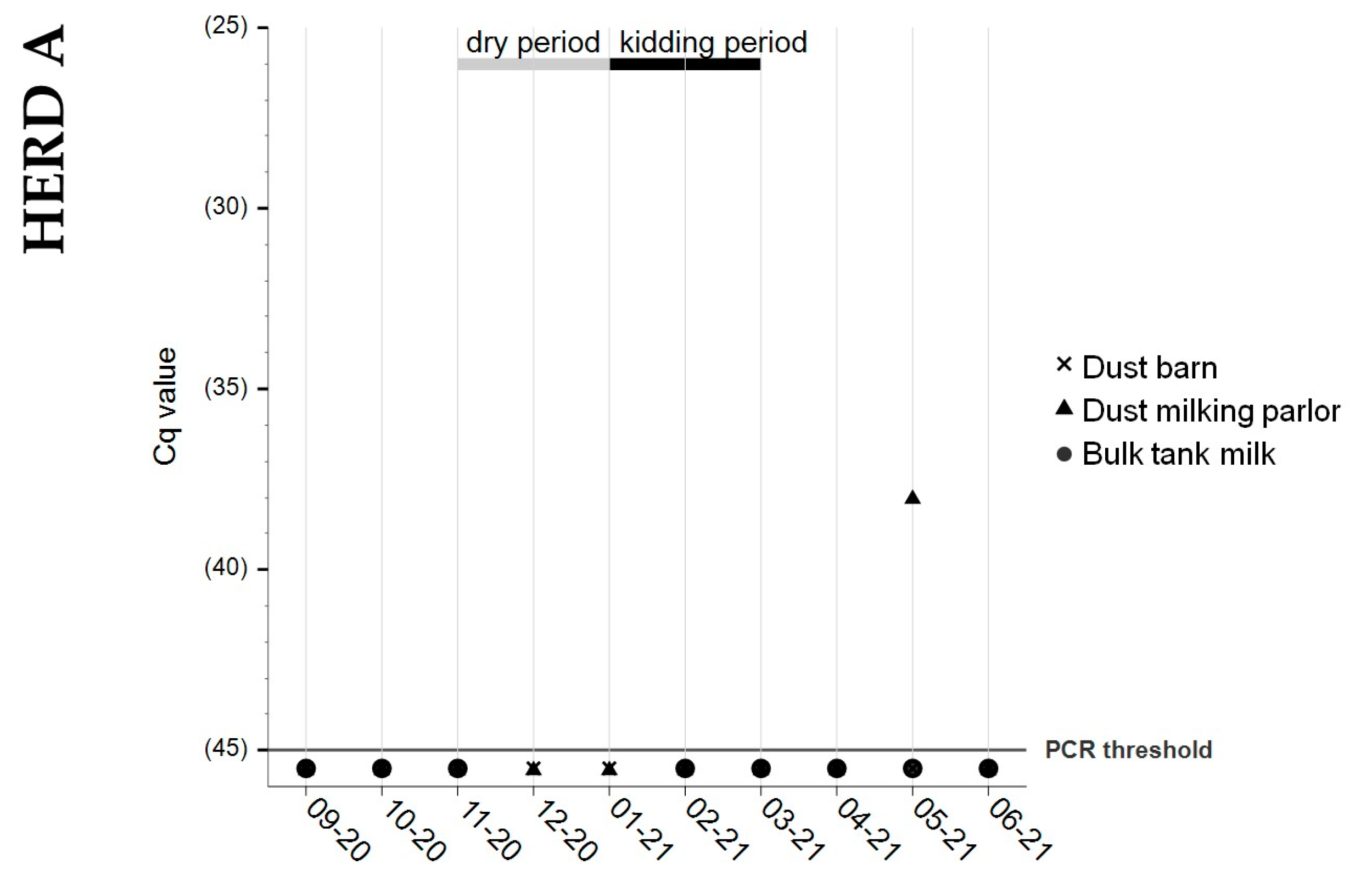
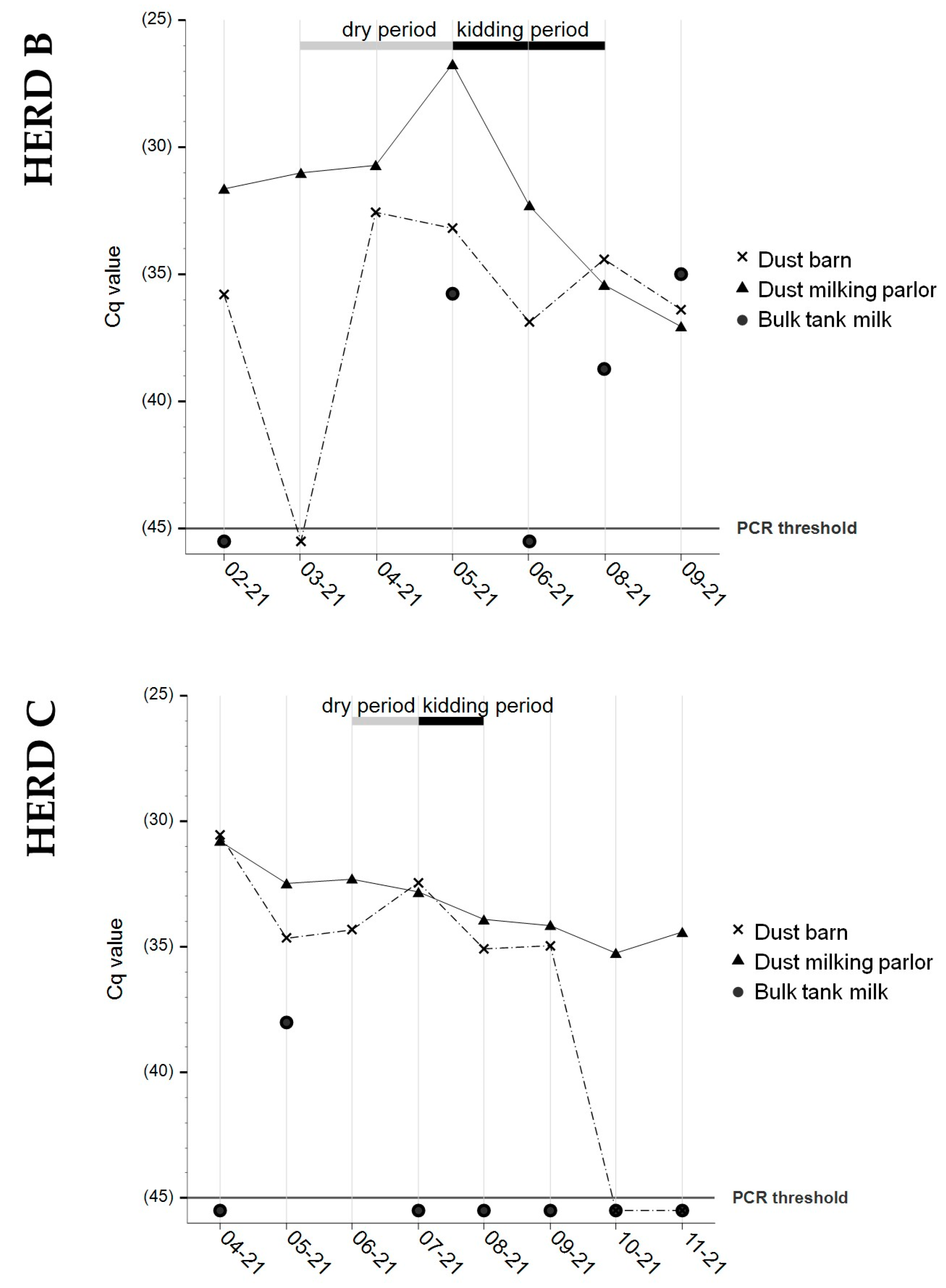
3.3. Bulk Tank Milk
3.4. Dust Samples from Barn and Milking Parlor
3.5. Abortion Material
3.6. Vaginal Swab and Phase-Specific Serology from Goat #D1 in Herd C
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BTM | Bulk tank milk |
| Cq | Cycle quantification value |
| ELISA | Enzyme-linked immunosorbent assay |
| FSVO | Swiss Federal Food Safety and Veterinary Office |
| ID50 | Median infectious dose |
| IgG PhI | Immunoglobulin G phase I |
| IgG PhII | Immunoglobulin G phase II |
| RU | Relative units |
| TSV | Swiss Ordinance on epizootic diseases |
References
- Arricau-Bouvery, N.; Rodolakis, A. Is Q fever an emerging or re-emerging zoonosis? Vet. Res. 2005, 36, 327–349. [Google Scholar] [CrossRef] [PubMed]
- Stoker, M.G.P.; Fiset, P. Phase variation of the Nine Mile and other strains of Rickettsia burneti. Can. J. Microbiol. 1956, 2, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Kreizinger, Z.; Szeredi, L.; Bacsadi, Á.; Nemes, C.; Sugár, L.; Varga, T.; Sulyok, K.M.; Szigeti, A.; Ács, K.; Tóbiás, E.; et al. Occurrence of Coxiella burnetii and Chlamydiales species in abortions of domestic ruminants and in wild ruminants in Hungary, Central Europe. J. Vet. Diagn. Investig. 2015, 27, 206–210. [Google Scholar] [CrossRef]
- Ferrara, G.; Colitti, B.; Pagnini, U.; D’Angelo, D.; Iovane, G.; Rosati, S.; Montagnaro, S. Serological evidence of Q fever among dairy cattle and buffalo populations in the Campania region, Italy. Pathogens 2022, 11, 901. [Google Scholar] [CrossRef]
- Georgiev, M.; Afonso, A.; Neubauer, H.; Needham, H.; Thiéry, R.; Rodolakis, A.; Roest, H.J.; Stärk, K.D.; Stegeman, J.A.; Vellema, P.; et al. Q fever in humans and farm animals in four European countries. Euro. Surveill. 2013, 18, 20407. [Google Scholar] [CrossRef]
- Palmer, N.C.; Kierstead, M.; Key, D.W.; Williams, J.C.; Peacock, M.G.; Vellend, H. Placentitis and abortion in goats and sheep in Ontario caused by Coxiella burnetii. Can. Vet. J. 1983, 24, 60–61. [Google Scholar] [PubMed]
- Agerholm, J.S. Coxiella burnetii associated reproductive disorders in domestic animals—A critical review. Acta Vet. Scand. 2013, 55, 13. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.; Prüfer, L.; Walter, M.; Ganter, I.; Frangoulidis, D.; Runge, M.; Ganter, M. Comparison of Coxiella burnetii excretion between sheep and goats naturally infected with one cattle-associated genotype. Pathogens 2020, 9, 652. [Google Scholar] [CrossRef]
- Arricau Bouvery, N.; Souriau, A.; Lechopier, P.; Rodolakis, A. Experimental Coxiella burnetii infection in pregnant goats: Excretion routes. Vet. Res. 2003, 34, 423–433. [Google Scholar] [CrossRef]
- Berri, M.; Rousset, E.; Champion, J.L.; Russo, P.; Rodolakis, A. Goats may experience reproductive failures and shed Coxiella burnetii at two successive parturitions after a Q fever infection. Res. Vet. Sci. 2007, 83, 47–52. [Google Scholar] [CrossRef]
- Sting, R.; Molz, K.; Philipp, W.; Bothe, F.; Runge, M.; Ganter, M. Quantitative real-time PCR and phase specific serology are mutually supportive in Q fever diagnostics in goats. Vet. Microbiol. 2013, 167, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Magouras, I.; Hunninghaus, J.; Scherrer, S.; Wittenbrink, M.M.; Hamburger, A.; Stärk, K.D.C.; Schüpbach-Regula, G. Coxiella burnetii infections in small ruminants and humans in Switzerland. Transbound. Emerg. Dis. 2017, 64, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Canevari, J.T.; Firestone, S.M.; Vincent, G.; Campbell, A.; Tan, T.; Muleme, M.; Cameron, A.W.N.; Stevenson, M.A. The prevalence of Coxiella burnetii shedding in dairy goats at the time of parturition in an endemically infected enterprise and associated milk yield losses. BMC Vet. Res. 2018, 14, 353. [Google Scholar] [CrossRef] [PubMed]
- Bontje, D.M.; Backer, J.A.; Hogerwerf, L.; Roest, H.I.J.; van Roermund, H.J.W. Analysis of Q fever in Dutch dairy goat herds and assessment of control measures by means of a transmission model. Prev. Vet. Med. 2016, 123, 71–89. [Google Scholar] [CrossRef]
- de Cremoux, R.; Rousset, E.; Touratier, A.; Audusseau, G.; Nicollet, P.; Ribaud, D.; David, V.; Le Pape, M. Coxiella burnetii vaginal shedding and antibody responses in dairy goat herds in a context of clinical Q fever outbreaks. FEMS Immunol. Med. Microbiol. 2012, 64, 120–122. [Google Scholar] [CrossRef]
- Mertens, K.; Gerlach, C.; Neubauer, H.; Henning, K. Q fever—An update. Curr. Clin. Microbiol. Rep. 2017, 4, 61–70. [Google Scholar] [CrossRef]
- Bauer, B.U.; Runge, M.; Campe, A.; Henning, K.; Mertens-Scholz, K.; Boden, K.; Sobotta, K.; Frangoulidis, D.; Knittler, M.R.; Matthiesen, S.; et al. Coxiella burnetii: A review focusing on infections in German sheep and goat flocks. Berl. Munch. Tierarztl. Wochenschr. 2020, 133, 184–200. [Google Scholar] [CrossRef]
- Schneeberger, P.M.; Wintenberger, C.; van der Hoek, W.; Stahl, J.P. Q fever in the Netherlands—2007–2010: What we learned from the largest outbreak ever. Med. Mal. Infect. 2014, 44, 339–353. [Google Scholar] [CrossRef]
- Brooke, R.J.; Kretzschmar, M.E.E.; Mutters, N.T.; Teunis, P.F. Human dose response relation for airborne exposure to Coxiella burnetii. BMC Infect. Dis. 2013, 13, 488. [Google Scholar] [CrossRef]
- Maurin, M.; Raoult, D. Q fever. Clin. Microbiol. Rev. 1999, 12, 518–553. [Google Scholar] [CrossRef]
- Dupuis, G.; Petite, J.; Péter, O.; Vouilloz, M. An important outbreak of human Q fever in a Swiss alpine valley. Int. J. Epidemiol. 1987, 16, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Kampschreur, L.M.; Delsing, C.E.; Groenwold, R.H.H.; Wegdam-Blans, M.C.A.; Bleeker-Rovers, C.P.; De Jager-Leclercq, M.G.L.; Hoepelman, A.I.M.; Van Kasteren, M.E.; Buijs, J.; Renders, N.H.M.; et al. Chronic Q fever in the Netherlands 5 years after the start of the Q fever epidemic: Results from the Dutch chronic Q fever database. J. Clin. Microbiol. 2014, 52, 1637–1643. [Google Scholar] [CrossRef] [PubMed]
- Landais, C.; Fenollar, F.; Thuny, F.; Raoult, D. From acute Q fever to endocarditis: Serological follow-up strategy. Clin. Infect. Dis. 2007, 44, 1337–1340. [Google Scholar] [CrossRef]
- Melenotte, C.; Million, M.; Raoult, D. New insights in Coxiella burnetii infection: Diagnosis and therapeutic update. Expert Rev. Anti Infect. Ther. 2020, 18, 75–86. [Google Scholar] [CrossRef]
- Signs, K.A.; Stobierski, M.G.; Gandhi, T.N. Q fever cluster among raw milk drinkers in Michigan, 2011. Clin. Infect. Dis. 2012, 55, 1387–1389. [Google Scholar] [CrossRef]
- Benson, W.W.; Brock, D.W.; Mather, J. Serologic analysis of a penitentiary group using raw milk from a Q fever infected herd. Public Health Rep. 1963, 78, 707–710. [Google Scholar] [CrossRef]
- Bellini, C.; Magouras, I.; Chapuis-Taillard, C.; Clerc, O.; Masserey, E.; Peduto, G.; Péter, O.; Schaerrer, S.; Schuepbach, G.; Greub, G. Q fever outbreak in the terraced vineyards of Lavaux, Switzerland. New Microbes New Infect. 2014, 2, 93–99. [Google Scholar] [CrossRef]
- Bundesamt für Lebensmittelsicherheit und Veterinärwesen BLV Q Fieber (Coxiellose) [Q fever (coxiellosis)]. Bericht zur Überwachung von Zoonosen und Lebensmittelbedingten Krankheitsausbrüchen—Daten 2021 [Report on Zoonotic and Food-Borne Diseases Surveillance—Data 2021]; Bundesamt für Lebensmittelsicherheit und Veterinärwesen BLV: Bern, Switzerland, 2022; pp. 30–32. [Google Scholar]
- Chanton-Greutmann, H.; Thoma, R.; Corboz, L.; Borel, N.; Pospischil, A. Aborte beim kleinen Wiederkäuer in der Schweiz: Untersuchungen während zwei Ablammperioden (1996–1998) unter besonderer Beachtung des Chlamydienabortes [Abortion in small ruminants in Switzerland: Investigations in two lambing seasons with special regard to Chlamydiae]. Schweiz. Arch. Tierheilkd. 2002, 144, 483–492. [Google Scholar] [CrossRef]
- Heinzelmann, M.; Rodriguez-Campos, S.; Kittl, S.; Zanolari, P.; Hirsbrunner, G. Aborte und Totgeburten bei Ziegen unter besonderer Berücksichtigung von Coxiella burnetii [Abortions and stillbirths caused by Coxiella burnetii in goats]. Schweiz. Arch. Tierheilkd. 2020, 162, 625–633. [Google Scholar] [CrossRef]
- Schnydrig, P.; Vidal, S.; Brodard, I.; Frey, C.F.; Posthaus, H.; Perreten, V.; Rodriguez-Campos, S. Bacterial, fungal, parasitological and pathological analyses of abortions in small ruminants from 2012–2016. Schweiz. Arch. Tierheilkd. 2017, 159, 647–656. [Google Scholar] [CrossRef]
- Klee, S.R.; Tyczka, J.; Ellerbrok, H.; Franz, T.; Linke, S.; Baljer, G.; Appel, B. Highly sensitive real-time PCR for specific detection and quantification of Coxiella burnetii. BMC Microbiol. 2006, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Körner, S.; Makert, G.R.; Ulbert, S.; Pfeffer, M.; Mertens-Scholz, K. The prevalence of Coxiella burnetii in hard ticks in Europe and their role in Q fever transmission revisited—A systematic review. Front. Vet. Sci. 2021, 8, 655715. [Google Scholar] [CrossRef] [PubMed]
- Jodełko, A.; Szymańska-Czerwińska, M.; Rola, J.G.; Niemczuk, K. Molecular detection of Coxiella burnetii in small ruminants and genotyping of specimens collected from goats in Poland. BMC Vet. Res. 2021, 17, 341. [Google Scholar] [CrossRef] [PubMed]
- WOAH. Q fever. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health (WOAH): Paris, France, 2023; pp. 560–577. [Google Scholar]
- Ferrara, G.; Colitti, B.; Pagnini, U.; Iovane, G.; Rosati, S.; Montagnaro, S. Characterization of recombinant Ybgf protein for the detection of Coxiella antibodies in ruminants. J. Vet. Diagn. Investig. 2022, 34, 646–653. [Google Scholar] [CrossRef]
- Bauer, B.U.; Schoneberg, C.; Herms, T.L.; Runge, M.; Ganter, M. Surveillance of Coxiella burnetii shedding in three naturally infected dairy goat herds after vaccination, focusing on bulk tank milk and dust swabs. Vet. Sci. 2022, 9, 102. [Google Scholar] [CrossRef]
- Roest, H.I.; Post, J.; Van Gelderen, B.; Van Zijderveld, F.G.; Rebel, J.M. Q fever in pregnant goats: Humoral and cellular immune responses. Vet. Res. 2013, 44, 67. [Google Scholar] [CrossRef]
- Hatchette, T.; Campbell, N.; Hudson, R.; Raoult, D.; Marrie, T.J. Natural history of Q fever in goats. Vector-Borne Zoonotic Dis. 2003, 3, 11–15. [Google Scholar] [CrossRef]
- Muleme, M.; Stenos, J.; Vincent, G.; Wilks, C.R.; Devlin, J.M.; Campbell, A.; Cameron, A.; Stevenson, M.A.; Graves, S.; Firestone, S.M. Peripartum dynamics of Coxiella burnetii infections in intensively managed dairy goats associated with a Q fever outbreak in Australia. Prev. Vet. Med. 2017, 139, 58–66. [Google Scholar] [CrossRef]
- Van Den Brom, R.; Van Engelen, E.; Luttikholt, S.; Moll, L.; Van Maanen, K.; Vellema, P. Coxiella burnetii in bulk tank milk samples from dairy goat and dairy sheep farms in the Netherlands in 2008. Vet. Rec. 2012, 170, 310. [Google Scholar] [CrossRef]
- Van den Brom, R.; Santman-Berends, I.; Luttikholt, S.; Moll, L.; Van Engelen, E.; Vellema, P. Bulk tank milk surveillance as a measure to detect Coxiella burnetii shedding dairy goat herds in the Netherlands between 2009 and 2014. J. Dairy Sci. 2015, 98, 3814–3825. [Google Scholar] [CrossRef]
- Jansen, W.; Cargnel, M.; Boarbi, S.; Mertens, I.; Van Esbroeck, M.; Fretin, D.; Mori, M. Belgian bulk tank milk surveillance program reveals the impact of a continuous vaccination protocol for small ruminants against Coxiella burnetii. Transbound. Emerg. Dis. 2022, 69, e141–e152. [Google Scholar] [CrossRef] [PubMed]
- van den Brom, R.; van Engelen, E.; Vos, J.; Luttikholt, S.J.M.; Moll, L.; Roest, H.I.J.; van der Heijden, H.M.J.F.; Vellema, P. Detection of Coxiella burnetii in the bulk tank milk from a farm with vaccinated goats, by using a specific PCR technique. Small Rumin. Res. 2013, 110, 150–154. [Google Scholar] [CrossRef]
- de Bruin, A.; de Groot, A.; de Heer, L.; Bok, J.; Wielinga, P.R.; Hamans, M.; van Rotterdam, B.J.; Janse, I. Detection of Coxiella burnetii in complex matrices by using multiplex quantitative PCR during a major Q fever outbreak in the Netherlands. Appl. Environ. Microbiol. 2011, 77, 6516–6523. [Google Scholar] [CrossRef]
- Carrié, P.; Barry, S.; Rousset, E.; de Crémoux, R.; Sala, C.; Calavas, D.; Perrin, J.B.; Bronner, A.; Gasqui, P.; Gilot-Fromont, E.; et al. Swab cloths as a tool for revealing environmental contamination by Q fever in ruminant farms. Transbound. Emerg. Dis. 2019, 66, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Kersh, G.J.; Fitzpatrick, K.A.; Self, J.S.; Priestley, R.A.; Kelly, A.J.; Ryan Lash, R.; Marsden-Haug, N.; Nett, R.J.; Bjork, A.; Massung, R.F.; et al. Presence and persistence of Coxiella burnetii in the environments of goat farms associated with a Q fever outbreak. Appl. Environ. Microbiol. 2013, 79, 1697–1703. [Google Scholar] [CrossRef]
- Álvarez-Alonso, R.; Basterretxea, M.; Barandika, J.F.; Hurtado, A.; Idiazabal, J.; Jado, I.; Beraza, X.; Montes, M.; Liendo, P.; García-Pérez, A.L. A Q fever outbreak with a high rate of abortions at a dairy goat farm: Coxiella burnetii shedding, environmental contamination, and viability. Appl. Environ. Microbiol. 2018, 84, e01650-18. [Google Scholar] [CrossRef]
- Vellema, P.; Santman-Berends, I.; Dijkstra, F.; van Engelen, E.; Aalberts, M.; Ter Bogt-Kappert, C.; van den Brom, R. Dairy sheep played a minor role in the 2005–2010 human Q fever outbreak in the Netherlands compared to dairy goats. Pathogens 2021, 10, 1579. [Google Scholar] [CrossRef]
- de Cremoux, R.; Rousset, E.; Touratier, A.; Audusseau, G.; Nicollet, P.; le Ribaud, D.; David, V.; Le Pape, M. Assessment of vaccination by a phase I Coxiella burnetii-inactivated vaccine in goat herds in clinical Q fever situation. FEMS Immunol. Med. Microbiol. 2012, 64, 104–106. [Google Scholar] [CrossRef]
- Bauer, B.U.; Knittler, M.R.; Prüfer, T.L.; Wolf, A.; Matthiesen, S.; Runge, M.; Ganter, M. Humoral immune response to Q fever vaccination of three sheep flocks naturally pre-infected with Coxiella burnetii. Vaccine 2021, 39, 1499–1507. [Google Scholar] [CrossRef]
- WOAH. Enzootic abortion of ewes (ovine chlamydiosis) (infection with Chlamydia abortus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health (WOAH): Paris, France, 2023; pp. 1456–1465. [Google Scholar]
- Pantchev, A.; Sting, R.; Bauerfeind, R.; Tyczka, J.; Sachse, K. New real-time PCR tests for species-specific detection of Chlamydophila psittaci and Chlamydophila abortus from tissue samples. Vet. J. 2009, 181, 145–150. [Google Scholar] [CrossRef]
- Rousset, E.; Prigent, M.; Brugidou, R.; Martel, I.; Grob, A.; Le Gall, G.; Kerninon, S.; Delaval, J.; Chassin, A.; Vassiloglou, B.; et al. Adoption by a network’s laboratories of a validated quantitative real-time PCR method for monitoring Q fever abortions in ruminant livestock. Euroreference 2012, 8, 21–28. [Google Scholar]
- Gay, L.; Melenotte, C.; Lopez, A.; Desnues, B.; Raoult, D.; Leone, M.; Mezouar, S.; Mege, J.L. Impact of sex hormones on macrophage responses to Coxiella burnetii. Front. Immunol. 2021, 12, 705088. [Google Scholar] [CrossRef] [PubMed]
- Howard, Z.P.; Omsland, A. Selective inhibition of Coxiella burnetii replication by the steroid hormone progesterone. Infect. Immun. 2020, 88, e00894-19. [Google Scholar] [CrossRef] [PubMed]
- Bauer, B.U.; Knittler, M.R.; Herms, T.L.; Frangoulidis, D.; Matthiesen, S.; Tappe, D.; Runge, M.; Ganter, M. Multispecies Q fever outbreak in a mixed dairy goat and cattle farm based on a new bovine-associated genotype of Coxiella burnetii. Vet. Sci. 2021, 8, 252. [Google Scholar] [CrossRef]
- Wolf, A.; Prüfer, T.L.; Schoneberg, C.; Campe, A.; Runge, M.; Ganter, M.; Bauer, B.U. Risk factors for an infection with Coxiella burnetii in German sheep flocks. Epidemiol. Infect. 2020, 148, e260. [Google Scholar] [CrossRef]
- Blumer, S.; Moestl, K.; Krametter-Froetscher, R.; Hässig, M.; Pospischil, A.; Borel, N. Untersuchung der Serokonversion auf Chlamydia abortus von Schafen aus der Region Vorarlberg vor und nach der Alpung [The seroconversion of Chlamydia abortus in sheep from the region of Vorarlberg before and after alpine pasturing]. Schweiz. Arch. Tierheilkd. 2012, 154, 13–17. [Google Scholar] [CrossRef]
- Álvarez-Alonso, R.; Zendoia, I.I.; Barandika, J.F.; Jado, I.; Hurtado, A.; López, C.M.; García-Pérez, A.L. Monitoring Coxiella burnetii infection in naturally infected dairy sheep flocks throughout four lambing seasons and investigation of viable bacteria. Front. Vet. Sci. 2020, 7, 352. [Google Scholar] [CrossRef]
- Schimmer, B.; Lenferink, A.; Schneeberger, P.; Aangenend, H.; Vellema, P.; Hautvast, J.; van Duynhoven, Y. Seroprevalence and risk factors for Coxiella burnetii (Q fever) seropositivity in dairy goat farmers’ households in the Netherlands, 2009–2010. PLoS ONE 2012, 7, e42364. [Google Scholar] [CrossRef]
- Samuel, J.E.; Hendrix, L.R. Laboratory maintenance of Coxiella burnetii. Curr. Protoc. Microbiol. 2009, Chapter 6. Unit 6C.1. [Google Scholar] [CrossRef]
- Omsland, A.; Beare, P.A.; Hill, J.; Cockrell, D.C.; Howe, D.; Hansen, B.; Samuel, J.E.; Heinzen, R.A. Isolation from animal tissue and genetic transformation of Coxiella burnetii are facilitated by an improved axenic growth medium. Appl. Environ. Microbiol. 2011, 77, 3720–3725. [Google Scholar] [CrossRef]
- Astobiza, I.; Barandika, J.F.; Ruiz-Fons, F.; Hurtado, A.; Povedano, I.; Juste, R.A.; García-Pérez, A.L. Coxiella burnetii shedding and environmental contamination at lambing in two highly naturally-infected dairy sheep flocks after vaccination. Res. Vet. Sci. 2011, 91, e58–e63. [Google Scholar] [CrossRef] [PubMed]
- Hogerwerf, L.; Borlée, F.; Still, K.; Heederik, D.; van Rotterdam, B.; de Bruin, A.; Nielen, M.; Wouters, I.M. Detection of Coxiella burnetii DNA in inhalable airborne dust samples from goat farms after mandatory culling. Appl. Environ. Microbiol. 2012, 78, 5410–5412. [Google Scholar] [CrossRef] [PubMed]
- Joulié, A.; Laroucau, K.; Bailly, X.; Prigent, M.; Gasqui, P.; Lepetitcolin, E.; Blanchard, B.; Rousset, E.; Sidi-Boumedine, K.; Jourdain, E. Circulation of Coxiella burnetii in a naturally infected flock of dairy sheep: Shedding dynamics, environmental contamination, and genotype diversity. Appl. Environ. Microbiol. 2015, 81, 7253–7260. [Google Scholar] [CrossRef]
- de los Ramo, M.A.; Benito, A.A.; Quílez, J.; Monteagudo, L.V.; Baselga, C.; Tejedor, M.T. Coxiella burnetii and co-infections with other major pathogens causing abortion in small ruminant flocks in the Iberian Peninsula. Animals 2022, 12, 3454. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.; Azenha, D.; Oliveira, C.; Almeida, A. Coxiella burnetii and Chlamydia spp. coinfection in small ruminant abortion in Portugal. Small Rumin. Res. 2022, 207, 106616. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal ID | PCR Result: C. burnetii (Material, Cq Value) | PCR Result: Chl. abortus (Material, Cq Value) |
|---|---|---|
| Herd A | ||
| Goat #152 (primiparous, purchased) | negative (fetus and placenta) | negative (fetus and placenta) |
| Goat #129 (primiparous) | negative (fetus) | positive (fetus, Cq 36.9) |
| Herd B | ||
| Goat #565 (purchased) | positive (placenta, Cq 39.3) negative (fetus) | positive (fetus, Cq 36.9; placenta, Cq 27.6) |
| Goat #252 (purchased) | positive (placenta, Cq 38.4) negative (fetus) | negative (fetus and placenta) |
| Herd C | ||
| Goat #20 (multiparous) | negative (vaginal swab *) | negative (vaginal swab *) |
| Goat #24 (multiparous) | negative (vaginal swab *) | negative (vaginal swab *) |
| Goat #909 | negative (fetus) | negative (fetus) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trachsel, C.; Hirsbrunner, G.; Herms, T.L.; Runge, M.; Kiene, F.; Ganter, M.; Zanolari, P.; Bauer, B.U. Two Years after Coxiella burnetii Detection: Pathogen Shedding and Phase-Specific Antibody Response in Three Dairy Goat Herds. Animals 2023, 13, 3048. https://doi.org/10.3390/ani13193048
Trachsel C, Hirsbrunner G, Herms TL, Runge M, Kiene F, Ganter M, Zanolari P, Bauer BU. Two Years after Coxiella burnetii Detection: Pathogen Shedding and Phase-Specific Antibody Response in Three Dairy Goat Herds. Animals. 2023; 13(19):3048. https://doi.org/10.3390/ani13193048
Chicago/Turabian StyleTrachsel, Christa, Gaby Hirsbrunner, T. Louise Herms, Martin Runge, Frederik Kiene, Martin Ganter, Patrik Zanolari, and Benjamin U. Bauer. 2023. "Two Years after Coxiella burnetii Detection: Pathogen Shedding and Phase-Specific Antibody Response in Three Dairy Goat Herds" Animals 13, no. 19: 3048. https://doi.org/10.3390/ani13193048
APA StyleTrachsel, C., Hirsbrunner, G., Herms, T. L., Runge, M., Kiene, F., Ganter, M., Zanolari, P., & Bauer, B. U. (2023). Two Years after Coxiella burnetii Detection: Pathogen Shedding and Phase-Specific Antibody Response in Three Dairy Goat Herds. Animals, 13(19), 3048. https://doi.org/10.3390/ani13193048