
Yeast Culture Supplementation Effects on Systemic and Polymorphonuclear Leukocytes’ mRNA Biomarkers of Inflammation and Liver Function in Peripartal Dairy Cows †

 , , ,
, , ,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design, and Dietary Treatments
2.2. Blood Collection and Analyses
2.3. PMNL Isolation
2.4. RNA Isolation, cDNA Synthesis, and Real-Time Quantitative PCR
2.5. Statistical Analysis
3. Results
3.1. Blood Biomarkers
3.2. Gene Expression
3.2.1. Inflammation
3.2.2. Cell Receptors and Signaling
3.2.3. Oxidative Stress
4. Discussion
4.1. Inflammation
4.2. Liver Function
4.3. Gene Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drackley, J.K. ADSA Foundation Scholar Award. Biology of dairy cows during the transition period: The final frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef] [PubMed]
- Goff, J.P.; Horst, R.L. Physiological changes at parturition and their relationship to metabolic disorders. J. Dairy Sci. 1997, 80, 1260–1268. [Google Scholar] [CrossRef]
- Mezzetti, M.; Bionaz, M.; Trevisi, E. Interaction between inflammation and metabolism in periparturient dairy cows. J. Anim. Sci. 2020, 98, S155–S174. [Google Scholar] [CrossRef] [PubMed]
- Dann, H.M.; Drackley, J.K.; McCoy, G.C.; Hutjens, M.F.; Garrett, J.E. Effects of yeast culture (Saccharomyces cerevisiae) on prepartum intake and postpartum intake and milk production of Jersey cows. J. Dairy Sci. 2000, 83, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Knoblock, C.E.; Yoon, I.; Oba, M. Effects of supplementing a Saccharomyces cerevisiae fermentation product during the transition period on rumen fermentation of dairy cows fed fresh diets differing in starch content. J. Dairy Sci. 2019, 102, 9943–9955. [Google Scholar] [CrossRef] [Green Version]
- Zaworski, E.M.; Shriver-Munsch, C.M.; Fadden, N.A.; Sanchez, W.K.; Yoon, I.; Bobe, G. Effects of feeding various dosages of Saccharomyces cerevisiae fermentation product in transition dairy cows. J. Dairy Sci. 2014, 97, 3081–3098. [Google Scholar] [CrossRef] [Green Version]
- Nocek, J.E.; Holt, M.G.; Oppy, J. Effects of supplementation with yeast culture and enzymatically hydrolyzed yeast on performance of early lactation dairy cattle. J. Dairy Sci. 2011, 94, 4046–4056. [Google Scholar] [CrossRef] [Green Version]
- Stefenoni, H.; Harrison, J.H.; Adams-Progar, A.; Block, E. Effect of enzymatically hydrolyzed yeast on health and performance of transition dairy cattle. J. Dairy Sci. 2020, 103, 1541–1552. [Google Scholar] [CrossRef]
- Lopreiato, V.; Mezzetti, M.; Cattaneo, L.; Ferronato, G.; Minuti, A.; Trevisi, E. Role of nutraceuticals during the transition period of dairy cows: A review. J. Anim. Sci. Biotechnol. 2020, 11, 96. [Google Scholar] [CrossRef] [PubMed]
- Jensen, G.S.; Patterson, K.M.; Yoon, I. Yeast culture has anti-inflammatory effects and specifically activates NK cells. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 487–500. [Google Scholar] [CrossRef]
- Yuan, K.; Mendonca, L.G.; Hulbert, L.E.; Mamedova, L.K.; Muckey, M.B.; Shen, Y.; Elrod, C.C.; Bradford, B.J. Yeast product supplementation modulated humoral and mucosal immunity and uterine inflammatory signals in transition dairy cows. J. Dairy Sci. 2015, 98, 3236–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knoblock, C.E.; Shi, W.; Yoon, I.; Oba, M. Effects of supplementing a Saccharomyces cerevisiae fermentation product during the periparturient period on the immune response of dairy cows fed fresh diets differing in starch content. J. Dairy Sci. 2019, 102, 6199–6209. [Google Scholar] [CrossRef] [PubMed]
- Aung, M.; Ohtsuka, H.; Izumi, K. Short communication: Effect of yeast cell wall supplementation on peripheral leukocyte populations and mRNA expression of cytokines in lactating dairy cows. J. Dairy Sci. 2020, 103, 5634–5640. [Google Scholar] [CrossRef]
- Carpinelli, N.A.; Halfen, J.; Trevisi, E.; Chapman, J.D.; Sharman, E.D.; Anderson, J.L.; Osorio, J.S. Effects of peripartal yeast culture supplementation on lactation performance, blood biomarkers, rumen fermentation, and rumen bacteria species in dairy cows. J. Dairy Sci. 2021, 104, 10727–10743. [Google Scholar] [CrossRef] [PubMed]
- Van Amburgh, M.E.; Collao-Saenz, E.A.; Higgs, R.J.; Ross, D.A.; Recktenwald, E.B.; Raffrenato, E.; Chase, L.E.; Overton, T.R.; Mills, J.K.; Foskolos, A. The Cornell Net Carbohydrate and Protein System: Updates to the model and evaluation of version 6.5. J. Dairy Sci. 2015, 98, 6361–6380. [Google Scholar] [CrossRef]
- Batistel, F.; Osorio, J.S.; Ferrari, A.; Trevisi, E.; Socha, M.T.; Loor, J.J. Immunometabolic Status during the Peripartum Period Is Enhanced with Supplemental Zn, Mn, and Cu from Amino Acid Complexes and Co from Co Glucoheptonate. PloS ONE 2016, 11, e0155804. [Google Scholar] [CrossRef] [Green Version]
- Jacometo, C.B.; Zhou, Z.; Luchini, D.; Trevisi, E.; Correa, M.N.; Loor, J.J. Maternal rumen-protected methionine supplementation and its effect on blood and liver biomarkers of energy metabolism, inflammation, and oxidative stress in neonatal Holstein calves. J. Dairy Sci. 2016, 99, 6753–6763. [Google Scholar] [CrossRef]
- Trevisi, E.; Amadori, M.; Cogrossi, S.; Razzuoli, E.; Bertoni, G. Metabolic stress and inflammatory response in high-yielding, periparturient dairy cows. Res. Vet. Sci. 2012, 93, 695–704. [Google Scholar] [CrossRef]
- Osorio, J.S.; Trevisi, E.; Ballou, M.A.; Bertoni, G.; Drackley, J.K.; Loor, J.J. Effect of the level of maternal energy intake prepartum on immunometabolic markers, polymorphonuclear leukocyte function, and neutrophil gene network expression in neonatal Holstein heifer calves. J. Dairy Sci. 2013, 96, 3573–3587. [Google Scholar] [CrossRef] [Green Version]
- Osorio, J.S.; Ji, P.; Drackley, J.K.; Luchini, D.; Loor, J.J. Smartamine M and MetaSmart supplementation during the peripartal period alter hepatic expression of gene networks in 1-carbon metabolism, inflammation, oxidative stress, and the growth hormone-insulin-like growth factor 1 axis pathways. J. Dairy Sci. 2014, 97, 7451–7464. [Google Scholar] [CrossRef]
- Rosa, F.; Michelotti, T.C.; St-Pierre, B.; Trevisi, E.; Osorio, J.S. Early Life Fecal Microbiota Transplantation in Neonatal Dairy Calves Promotes Growth Performance and Alleviates Inflammation and Oxidative Stress during Weaning. Animals 2021, 11, 2704. [Google Scholar] [CrossRef] [PubMed]
- Moyes, K.M.; Drackley, J.K.; Morin, D.E.; Loor, J.J. Greater expression of TLR2, TLR4, and IL6 due to negative energy balance is associated with lower expression of HLA-DRA and HLA-A in bovine blood neutrophils after intramammary mastitis challenge with Streptococcus uberis. Funct. Integr. Genom. 2010, 10, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [Green Version]
- Sordillo, L.M. Nutritional strategies to optimize dairy cattle immunity. J. Dairy Sci. 2016, 99, 4967–4982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannerman, D.D.; Rinaldi, M.; Vinyard, B.T.; Laihia, J.; Leino, L. Effects of intramammary infusion of cis-urocanic acid on mastitis-associated inflammation and tissue injury in dairy cows. Am. J. Vet. Res. 2009, 70, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Trevisi, E.; Zecconi, A.; Bertoni, G.; Piccinini, R. Blood and milk immune and inflammatory profiles in periparturient dairy cows showing a different liver activity index. The Journal of dairy research 2010, 77, 310–317. [Google Scholar] [CrossRef]
- Ceciliani, F.; Ceron, J.J.; Eckersall, P.D.; Sauerwein, H. Acute phase proteins in ruminants. J. Proteom. 2012, 75, 4207–4231. [Google Scholar] [CrossRef]
- Mumbach, T.; Demarco, C.F.; Freitas, V.O.; Grazziotin, R.C.B.; Maf, A.S.; Gonçalves, F.M.; Brauner, C.C.; Jacometo, C.B.; Corrêa, M.N. Metabolic and ruminal fluid markers of dairy cows supplemented with a combination of yeast culture and hydrolyzed yeast. Acta Sci. Vet. 2017, 45, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Bertoni, G.; Trevisi, E.; Han, X.; Bionaz, M. Effects of inflammatory conditions on liver activity in puerperium period and consequences for performance in dairy cows. J. Dairy Sci. 2008, 91, 3300–3310. [Google Scholar] [CrossRef] [Green Version]
- Faccio-Demarco, C.; Mumbach, T.; Oliveira-de-Freitas, V.; Fraga, E.S.-R.R.; Medeiros-Goncalves, F.; Nunes-Correa, M.; Burkert-Del Pino, F.A.; Mendonca-Nunes-Ribeiro Filho, H.; Cassal-Brauner, C. Effect of yeast products supplementation during transition period on metabolic profile and milk production in dairy cows. Trop. Anim. Health Prod. 2019, 51, 2193–2201. [Google Scholar] [CrossRef]
- Bertoni, G.; Trevisi, E. Use of the liver activity index and other metabolic variables in the assessment of metabolic health in dairy herds. Vet. Clin. N. Am. Food Anim. Pract. 2013, 29, 413–431. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Jimenez, S.; Haerr, K.J.; Trevisi, E.; Loor, J.J.; Cardoso, F.C.; Osorio, J.S. Prepartal standing behavior as a parameter for early detection of postpartal subclinical ketosis associated with inflammation and liver function biomarkers in peripartal dairy cows. J. Dairy Sci. 2018, 101, 8224–8235. [Google Scholar] [CrossRef]
- Mezzetti, M.; Minuti, A.; Piccioli-Cappelli, F.; Amadori, M.; Bionaz, M.; Trevisi, E. The role of altered immune function during the dry period in promoting the development of subclinical ketosis in early lactation. J. Dairy Sci. 2019, 102, 9241–9258. [Google Scholar] [CrossRef]
- Al Ibrahim, R.M.; Kelly, A.K.; O’Grady, L.; Gath, V.P.; McCarney, C.; Mulligan, F.J. The effect of body condition score at calving and supplementation with Saccharomyces cerevisiae on milk production, metabolic status, and rumen fermentation of dairy cows in early lactation. J. Dairy Sci. 2010, 93, 5318–5328. [Google Scholar] [CrossRef] [Green Version]
- Aung, M.; Ohtsuka, H.; Izumi, K. Effect of yeast cell wall supplementation on production performances and blood biochemical indices of dairy cows in different lactation periods. Vet. World 2019, 12, 796–801. [Google Scholar] [CrossRef] [Green Version]
- Adeyemi, J.A.; Harmon, D.L.; Compart, D.M.P.; Ogunade, I.M. Effects of a blend of Saccharomyces cerevisiae-based direct-fed microbial and fermentation products in the diet of newly weaned beef steers: Growth performance, whole-blood immune gene expression, serum biochemistry, and plasma metabolome1. J. Anim. Sci. 2019, 97, 4657–4667. [Google Scholar] [CrossRef] [PubMed]
- Zanello, G.; Berri, M.; Dupont, J.; Sizaret, P.Y.; D’Inca, R.; Salmon, H.; Meurens, F. Saccharomyces cerevisiae modulates immune gene expressions and inhibits ETEC-mediated ERK1/2 and p38 signaling pathways in intestinal epithelial cells. PloS ONE 2011, 6, e18573. [Google Scholar] [CrossRef] [PubMed]
- Weedman, S.M.; Rostagno, M.H.; Patterson, J.A.; Yoon, I.; Fitzner, G.; Eicher, S.D. Yeast culture supplement during nursing and transport affects immunity and intestinal microbial ecology of weanling pigs. J. Anim. Sci. 2011, 89, 1908–1921. [Google Scholar] [CrossRef] [Green Version]
- Erminio, T.; Massimo, A.; Ivonne, A.; Nicola, L.; Giuseppe, B. Inflammatory Response and Acute Phase Proteins in the Transition Period of High-Yielding Dairy Cows. In Acute Phase Proteins as Early Non-Specific Biomarkers of Human and Veterinary Diseases; Francisco, V., Ed.; IntechOpen: Rijeka, Croatia, 2011. [Google Scholar]
- Moyes, K.M.; Graugnard, D.E.; Khan, M.J.; Mukesh, M.; Loor, J.J. Postpartal immunometabolic gene network expression and function in blood neutrophils are altered in response to prepartal energy intake and postpartal intramammary inflammatory challenge. J. Dairy Sci. 2014, 97, 2165–2177. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Bazzoni, F.; Tamassia, N.; Rossato, M.; Cassatella, M.A. Understanding the molecular mechanisms of the multifaceted IL-10-mediated anti-inflammatory response: Lessons from neutrophils. Eur. J. Immunol. 2010, 40, 2360–2368. [Google Scholar] [CrossRef]
- Zhou, Z.; Bu, D.P.; Vailati Riboni, M.; Khan, M.J.; Graugnard, D.E.; Luo, J.; Cardoso, F.C.; Loor, J.J. Prepartal dietary energy level affects peripartal bovine blood neutrophil metabolic, antioxidant, and inflammatory gene expression. J. Dairy Sci. 2015, 98, 5492–5505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batistel, F.; Osorio, J.S.; Tariq, M.R.; Li, C.; Caputo, J.; Socha, M.T.; Loor, J.J. Peripheral leukocyte and endometrium molecular biomarkers of inflammation and oxidative stress are altered in peripartal dairy cows supplemented with Zn, Mn, and Cu from amino acid complexes and Co from Co glucoheptonate. J. Anim. Sci. Biotechnol. 2017, 8, 33. [Google Scholar] [CrossRef] [Green Version]
- Paape, M.J.; Bannerman, D.D.; Zhao, X.; Lee, J.W. The bovine neutrophil: Structure and function in blood and milk. Vet. Res. 2003, 34, 597–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raffler, N.A.; Rivera-Nieves, J.; Ley, K. L-selectin in inflammation, infection and immunity. Drug Discov. Today Ther. Strateg. 2005, 2, 213–220. [Google Scholar] [CrossRef]
- Kuriakose, T.; Kanneganti, T.D. ZBP1: Innate Sensor Regulating Cell Death and Inflammation. Trends Immunol. 2018, 39, 123–134. [Google Scholar] [CrossRef]
- Izard, T.; Brown, D.T. Mechanisms and Functions of Vinculin Interactions with Phospholipids at Cell Adhesion Sites. J. Biol. Chem. 2016, 291, 2548–2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bays, J.L.; DeMali, K.A. Vinculin in cell-cell and cell-matrix adhesions. Cell. Mol. Life Sci. 2017, 74, 2999–3009. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.W.; Zhu, H.L.; Qi, Y.X.; Wu, T.; Huang, D.W.; Ding, H.S.; Chen, S.; Li, M.; Cheng, G.L.; Zhao, H.L.; et al. Quantitative comparative phosphoproteomic analysis of the effects of colostrum and milk feeding on liver tissue of neonatal calves. J. Dairy Sci. 2021, 104, 8265–8275. [Google Scholar] [CrossRef]
- Brown, C.; Morham, S.G.; Walsh, D.; Naghavi, M.H. Focal adhesion proteins talin-1 and vinculin negatively affect paxillin phosphorylation and limit retroviral infection. J. Mol. Biol. 2011, 410, 761–777. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Raphael, W. Significance of metabolic stress, lipid mobilization, and inflammation on transition cow disorders. Vet. Clin. N. Am. Food Anim. Pract. 2013, 29, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Abuelo, A.; Hernandez, J.; Benedito, J.L.; Castillo, C. The importance of the oxidative status of dairy cattle in the periparturient period: Revisiting antioxidant supplementation. J. Anim. Physiol. Anim. Nutr. 2015, 99, 1003–1016. [Google Scholar] [CrossRef] [Green Version]
- Ford, H.R.; Busato, S.; Bionaz, M. In vitro–In vivo Hybrid Approach for Studying Modulation of NRF2 in Immortalized Bovine Mammary Cells. Front. Anim. Sci. 2021, 2, 674355. [Google Scholar] [CrossRef]
- Vivarini, A.C.; Lopes, U.G. The Potential Role of Nrf2 Signaling in Leishmania Infection Outcomes. Front. Cell Infect. Microbiol. 2019, 9, 453. [Google Scholar] [CrossRef]
- Scholte, C.M.; Rezamand, P.; Tsai, C.Y.; Amiri, Z.M.; Ramsey, K.C.; McGuire, M.A. The effects of elevated subcutaneous fat stores on fatty acid composition and gene expression of proinflammatory markers in periparturient dairy cows. J. Dairy Sci. 2017, 100, 2104–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celi, P.; Gabai, G. Oxidant/Antioxidant Balance in Animal Nutrition and Health: The Role of Protein Oxidation. Front. Vet. Sci. 2015, 2, 48. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Guo, R.F.; Newstead, M.W.; Standiford, T.J.; Macariola, D.R.; Shanley, T.P. Effect of IL-10 on neutrophil recruitment and survival after Pseudomonas aeruginosa challenge. Am. J. Respir. Cell. Mol. Biol. 2009, 41, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Yoshidome, H.; Kato, A.; Edwards, M.J.; Lentsch, A.B. Interleukin-10 inhibits pulmonary NF-kappaB activation and lung injury induced by hepatic ischemia-reperfusion. Am. J. Physiol. 1999, 277, L919–L923. [Google Scholar] [CrossRef]


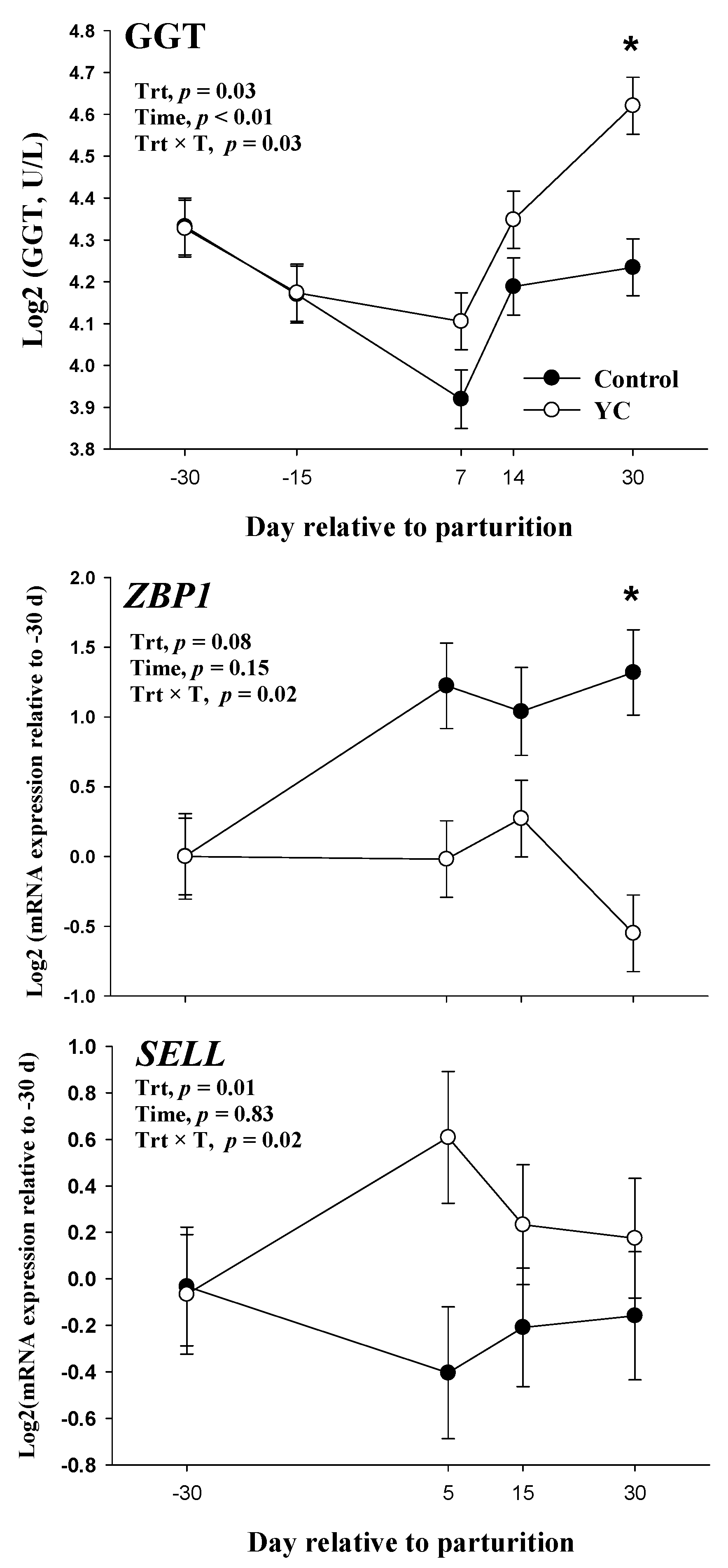{kind=link}
| Parameter 1 | Treatment | p-Value3 | |||||
|---|---|---|---|---|---|---|---|
| Control | YC | SEM 2 | Trt | Parity | Time | Trt × T | |
| Liver function | |||||||
| Total bilirubin, µmol/L 4 | 1.65 | 1.88 | 0.17 | 0.42 | --- | <0.01 | 0.77 |
| GOT, U/L | 95.0 | 95.0 | 3.18 | 0.99 | --- | <0.01 | 0.90 |
| GGT, U/L 4 | 18.0 | 19.9 | 0.04 | 0.03 | 0.03 | <0.01 | 0.03 |
| Cholesterol, mmol/L | 3.70 | 3.49 | 0.10 | 0.20 | --- | <0.01 | 0.96 |
| PON, U/mL | 106.2 | 97.9 | 3.50 | 0.12 | 0.03 | <0.01 | 0.16 |
| Inflammation | |||||||
| Albumin, g/L | 36.0 | 34.1 | 0.31 | <0.01 | 0.15 | <0.01 | 0.53 |
| Ceruloplasmin, µmol/L | 2.43 | 2.46 | 0.06 | 0.72 | --- | 0.03 | 0.15 |
| Haptoglobin, g/L 4 | 0.27 | 0.33 | 0.15 | 0.25 | --- | 0.09 | 0.59 |
| IL-1β, pg/mL 4 | 33.8 | 31.5 | 0.18 | 0.70 | 0.09 | <0.01 | 0.23 |
| Gene 1 | Treatment 2 | SEM 3 | p-Value4 | ||||
|---|---|---|---|---|---|---|---|
| Control | YC | Trt | Parity | Time | Trt × T | ||
| Inflammation | |||||||
| TNFA | 0.40 | 1.23 | 0.25 | 0.04 | --- | <0.01 | 0.21 |
| NFKB1 | −0.02 | 0.23 | 0.07 | 0.04 | --- | 0.33 | 0.42 |
| IL1B | 0.13 | −0.04 | 0.28 | 0.68 | --- | 0.94 | 0.72 |
| IL10 | 0.001 | 1.36 | 0.27 | 0.04 | --- | <0.01 | 0.21 |
| TRAF6 | 0.04 | 0.45 | 0.15 | 0.07 | --- | 0.08 | 0.16 |
| MYD88 | −0.10 | 0.12 | 0.14 | 0.30 | --- | 0.75 | 0.23 |
| Cell receptors and signaling | |||||||
| TLR2 | 0.14 | 0.41 | 0.12 | 0.11 | 0.05 | 0.04 | 0.32 |
| TLR4 | −0.08 | 0.15 | 0.17 | 0.33 | --- | 0.83 | 0.45 |
| SELL | −0.20 | 0.24 | 0.10 | 0.01 | 0.12 | 0.83 | 0.02 |
| IRAK1 | 0.10 | 0.26 | 0.12 | 0.37 | --- | 0.24 | 0.73 |
| ZBP1 | −0.07 | 0.89 | 0.17 | <0.01 | 0.02 | 0.15 | 0.02 |
| ITGAM | −0.83 | −0.68 | 0.19 | 0.56 | --- | <0.01 | 0.91 |
| TLN1 | −0.12 | 0.009 | 0.07 | 0.22 | --- | 0.15 | 0.52 |
| VCL | −0.63 | −0.32 | 0.12 | 0.08 | --- | <0.01 | 0.41 |
| Oxidative stress | |||||||
| NRF2 | 0.12 | 0.67 | 0.23 | 0.09 | 0.06 | 0.06 | 0.27 |
| SOD1 | −0.19 | 0.52 | 0.27 | 0.07 | --- | 0.67 | 0.67 |
| SOD2 | 0.11 | 0.17 | 0.29 | 0.88 | 0.07 | 0.27 | 0.81 |
| MPO | −2.00 | −3.27 | 0.52 | 0.10 | --- | <0.01 | 0.23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carpinelli, N.A.; Halfen, J.; Michelotti, T.C.; Rosa, F.; Trevisi, E.; Chapman, J.D.; Sharman, E.S.; Osorio, J.S. Yeast Culture Supplementation Effects on Systemic and Polymorphonuclear Leukocytes’ mRNA Biomarkers of Inflammation and Liver Function in Peripartal Dairy Cows. Animals 2023, 13, 301. https://doi.org/10.3390/ani13020301
Carpinelli NA, Halfen J, Michelotti TC, Rosa F, Trevisi E, Chapman JD, Sharman ES, Osorio JS. Yeast Culture Supplementation Effects on Systemic and Polymorphonuclear Leukocytes’ mRNA Biomarkers of Inflammation and Liver Function in Peripartal Dairy Cows. Animals. 2023; 13(2):301. https://doi.org/10.3390/ani13020301
Chicago/Turabian StyleCarpinelli, Nathaly Ana, Jessica Halfen, Tainara Cristina Michelotti, Fernanda Rosa, Erminio Trevisi, Jim D. Chapman, Evin S. Sharman, and Johan S. Osorio. 2023. "Yeast Culture Supplementation Effects on Systemic and Polymorphonuclear Leukocytes’ mRNA Biomarkers of Inflammation and Liver Function in Peripartal Dairy Cows" Animals 13, no. 2: 301. https://doi.org/10.3390/ani13020301
APA StyleCarpinelli, N. A., Halfen, J., Michelotti, T. C., Rosa, F., Trevisi, E., Chapman, J. D., Sharman, E. S., & Osorio, J. S. (2023). Yeast Culture Supplementation Effects on Systemic and Polymorphonuclear Leukocytes’ mRNA Biomarkers of Inflammation and Liver Function in Peripartal Dairy Cows. Animals, 13(2), 301. https://doi.org/10.3390/ani13020301