SARS-CoV-2 Infection in Captive Hippos (Hippopotamus amphibius), Belgium

 , , , , , ,
, , , , , ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Samples and Methods
3. Results
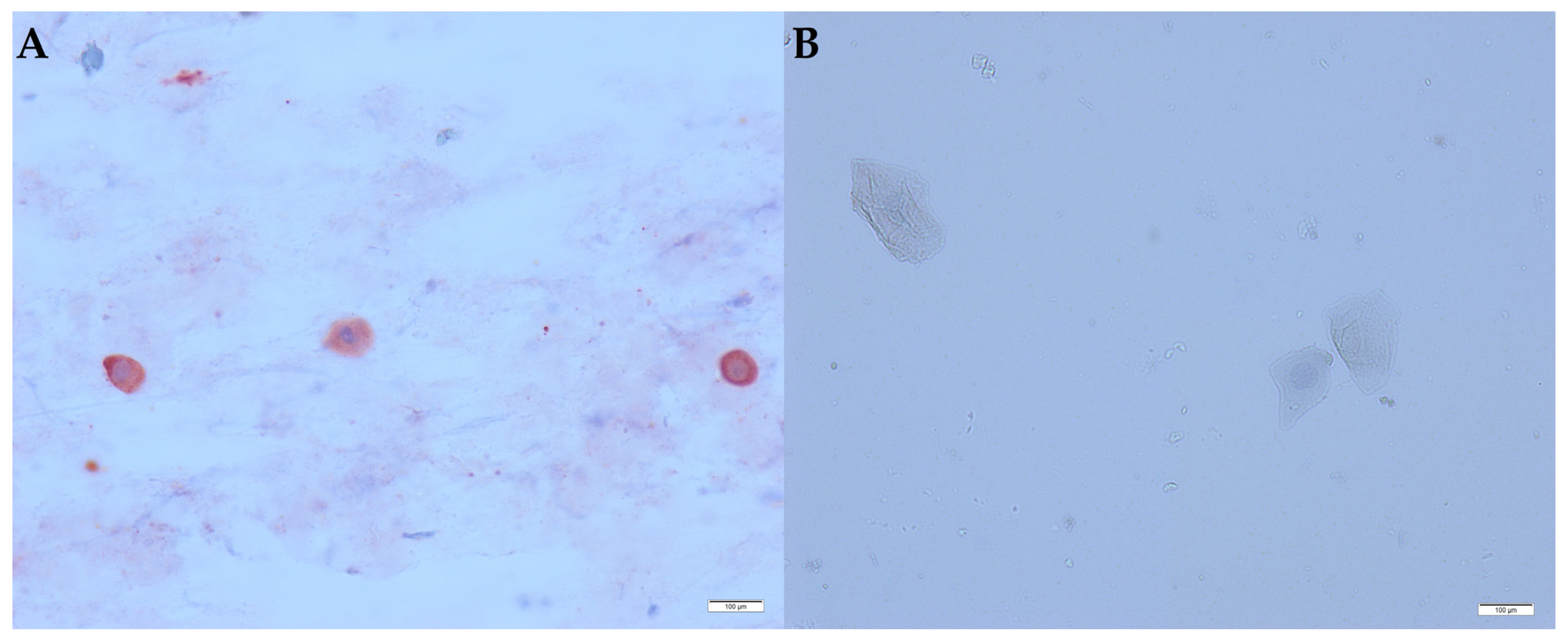
3.1. Interpretation of the Cytology and the Immunocytochemistry
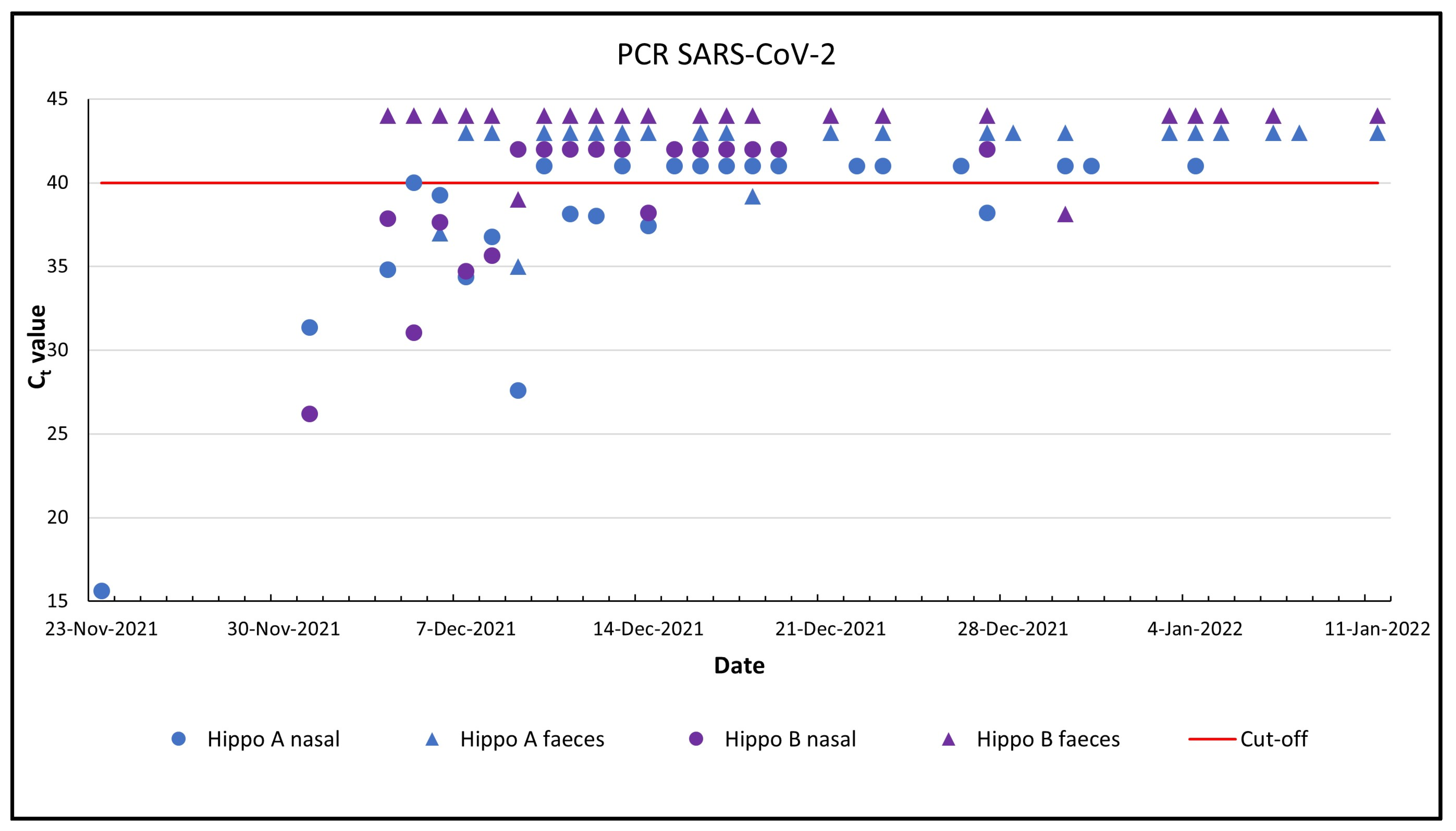
3.2. Analysis of Nasal Swabs with PCR and Sequencing
3.3. Analysis of Faeces and Pool Water with PCR
3.4. ELISA Analysis of Saliva and Serum Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gryseels, S.; De Bruyn, L.; Gyselings, R.; Calvignac-Spencer, S.; Leendertz, F.H.; Leirs, H. Risk of human-to-wildlife transmission of SARS-CoV-2. Mammal Rev. 2021, 51, 272–292. [Google Scholar] [CrossRef] [PubMed]
- Lecu, A. SARS-CoV-2 coronavirus infection in wild animals. In Fowler’s Zoo and Wild Animal Medicine Current Therapy; Miller, E., Calle, P.P., Lamberski, N., Eds.; Elsevier: St. Louis, MO, USA, 2023; Volume 10, pp. 137–143. [Google Scholar]
- Lecu, A.; Bertelsen, M.F.; Walzer, C. Science-Based Facts & Knowledge about Wild Animals, Zoos and SARS-CoV-2 Virus. Transmissible Diseases Handbook of the European Association of Zoo and Wildlife Veterinarians 2022, 17th January. Available online: https:/www.eazwv.org/page/inf_handbook (accessed on 18 April 2022).
- Oreshkova, N.; Molenaar, R.J.; Vreman, S.; Harders, F.; Oude Munnink, B.B.; Hakze-van der Honing, R.W.; Gerhards, N.; Tolsma, P.; Bouwstra, R.; Sikkema, R.; et al. SARS-CoV-2 infection in farmed minks, the Netherlands, April and May 2020. Euro Surveill. 2020, 25, 2001005. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.C.S.; Lam, S.D.; Damien, R.; Owen, C.J.; Berchtold, D.; Orengo, C.; Nair, M.S.; Kuchipudi, S.V.; Kapur, V.; van Dorp, L. Transmission of SARS-CoV-2 from humans to animals and potential host adaptation. Nat. Commun. 2022, 13, 2988. [Google Scholar] [CrossRef] [PubMed]
- Kuchipudia, S.V.; Surendran-Naira, M.; Ruden, M.R.; Yon, M.; Nissly, R.H.; Vandegrift, K.J.; Nelli, R.K.; Li, L.; Jayarao, B.M.; Maranas, C.D.; et al. Multiple spillovers from humans and onward transmission of SARS-CoV-2 in white-tailed deer. Proc. Natl. Acad. Sci. USA 2022, 119, e2121644119. [Google Scholar] [CrossRef] [PubMed]
- Lewison, R.; Pluháček, J. Hippopotamus amphibius. The IUCN Red List of Threatened Species 2017. Available online: https://www.iucnredlist.org/species/10103/18567364 (accessed on 18 April 2022).
- Sun, Y.; Ge, L.; Udhane, S.S.; Langenheim, J.F.; Rau, M.J.; Patton, M.D.; Gallan, A.; Felix, J.; Rui, H. Sensitive and Specific Immunohistochemistry Protocol for Nucleocapsid Protein from All Common SARS-CoV-2 Virus Strains in Formalin-Fixed, Paraffin Embedded Tissues. Methods Protoc. 2021, 4, 47. [Google Scholar] [CrossRef]
- CDC Centers for Disease Control and Prevention. 2019 nCoV CDC Kit. Available online: https://www.cdc.gov/coronavirus/2019-ncov/lab/guidelines-clinical-specimens.html (accessed on 18 April 2022).
- Freed, N.E.; Vlkova, M.; Faisal, M.B.; Silander, O.K. Rapid and inexpensive whole-genome sequencing of SARS-CoV-2 using 1200 bp tiled amplicons and Oxford Nanopore Rapid Barcoding. Biol. Methods Protoc. 2020, 5, bpaa014. [Google Scholar] [CrossRef] [PubMed]
- ARTIC Bioinformatics Pipeline. A Bioinformatics Pipeline for Working with Virus Sequencing Data Sequenced with Nanopore. Available online: https://github.com/artic-network/fieldbioinformatics (accessed on 18 April 2022).
- O’Toole, Á.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of epidemiological lineages in an emerging pandemic using the pangolin tool. Virus Evol. 2021, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Wang, L.; Sakthivel, S.K.; Whitaker, B.; Murray, J.; Kamili, S.; Lynch, B.; Malapati, L.; Burke, S.; Harcourt, J.; et al. US CDC Real-Time Reverse Transcription PCR Panel for Detection of Severe Acute Respiratory Syndrome Coronavirus 2. Emerg. Infect. Dis. 2020, 26, 1654–1665. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.W.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill. 2020, 25, 23–30. [Google Scholar] [CrossRef] [Green Version]
- GeurtsvanKessel, C.H.; Okba, N.M.A.; Igloi, Z.; Bogers, S.; Embregts, C.W.E.; Laksono, B.M.; Leijten, L.; Rokx, C.; Rijnders, B.; Rahamat-Langendoen, J.; et al. An evaluation of COVID-19 serological assays informs future diagnostics and exposure assessment. Nat. Commun. 2020, 11, 3436. [Google Scholar] [CrossRef]
- Damas, J.; Hughes, G.M.; Keough, K.C.; Painter, C.A.; Persky, N.S.; Corbo, M.; Hiller, M.; Koepfli, K.-P.; Pfenning, A.; Zhao, H.; et al. Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc. Natl. Acad. Sci. USA 2020, 117, 22311–22322. [Google Scholar] [CrossRef] [PubMed]
- Fischhoff, I.R.; Castellanos, A.A.; Rodrigues, J.P.G.L.M.; Varsani, A.; Han, B.A. Predicting the zoonotic capacity of mammals to transmit SARS-CoV-2. Proc. R. Soc. B. 2021, 288, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Praharaj, M.R.; Garg, P.; Kesarwani, V.; Topno, N.A.; Khan, R.I.N.; Sharma, S.; Panigrahi, M.; Mishra; Mishra, B.; Kumar, S.; et al. SARS-CoV-2 spike glycoprotein and ACE2 interaction reveals modulation of viral entry in wild and domestic animals. Front. Med. 2022, 8, 775572. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, S.L.; Suresh, M. Factors influencing the epidemiological characteristics of pandemic COVID 19: A TISM approach. Int. J. Healthc.Manag. 2020, 13, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Nagy, A.; Stará, M.; Vodička, R.; Černíková, L.; Jiřincová, H.; Křivda, V.; Sedlák, K. Reverse-zoonotic transmission of SARS-CoV-2 lineage alpha (B.1.1.7) to great apes and exotic felids in a zoo in the Czech Republic. Arch. Virol. 2022, 167, 1681–1685. [Google Scholar] [CrossRef]
- Bartlett, S.L.; Diel, G.; Wang, L.; Zec, S.; Laverack, M.; Martins, M.; Caserta, L.C.; Killian, M.L.; Terio, K.; Olmstead, C.; et al. Sars-COV-2 infection and longitudinal fecal screening in Malayan tigers (Panthera tigris jacksoni), Amur tigers (Panthera tigris altaica), and African lions (Panthera leo krugeri) at the bronx Zoo, New York, USA. J. Zoo Wildl. Med. 2020, 51, 733–744. [Google Scholar] [CrossRef]
- Fernández-Bellon, H.; Rodon, J.; Fernández-Bastit, L.; Almagro, V.; Padilla-Solé, P.; Lorca-Oró, C.; Valle, R.; Roca, N.; Grazioli, S.; Trogu, T.; et al. Monitoring natural SARS-CoV-2 infection in lions (Panthera leo) at the Barcelona zoo: Viral dynamics and host responses. Viruses 2021, 13, 1683. [Google Scholar] [CrossRef]
- McAloose, D.; Laverack, M.; Wang, L.; Killian, M.L.; Caserta, L.C.; Yuan, F.; Mitchell, P.K.; Queen, K.; Mauldin, M.R.; Cronk, B.D.; et al. From people to Panthera: Natural SARS-CoV-2 infection in tigers and lions at the Bronx Zoo. MBio 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Singanayagam, A.; Hakki, S.; Dunning, J.; Madon, K.J.; Crone, M.A.; Koycheva, A.; Derqui-Fernandez, N.; Barnett, J.; Whitfield, M.; Varro, R.; et al. Community transmission and viral load kinetics of the SARS-CoV-2 delta (B.1.617.2) variant in vaccinated and unvaccinated individuals in the UK: A prospective, longitudinal, cohort study. Lancet Infect. Dis. 2022, 22, 183–195. [Google Scholar] [CrossRef]
- Wurtzer, S.; Marechal, V.; Mouchel, J.M.; Maday, Y.; Teyssou, R.; Richard, E.; Almayrac, J.L.; Moulin, L. Evaluation of lockdown effect on SARS-CoV-2 dynamics through viral genome quantification in waste water, Greater Paris, France, 5 March to 23 April 2020. Euro Surveill. 2020, 25, 2000776. [Google Scholar] [CrossRef]
- Jones, D.L.; Baluja, M.Q.; Graham, D.W.; Corbishley, A.; McDonald, J.E.; Malham, S.K.; Hillary, L.; Connor, T.; Gaze, W.; Moura, I.; et al. Shedding of SARS-CoV-2 in feces and urine and its potential role in person-to-person transmission and the environment-based spread of COVID-19. Sci. Total Environ. 2020, 749, 141364. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.V.; Martins, M.; Falkenberg, S.; Buckley, A.; Caserta, L.C.; Mitchell, P.K.; Cassmann, E.; Rollins, A.; Zylich, N.; Renshaw, R.; et al. Susceptibility of White-Tailed Deer (Odocoileus virginianus) to SARS-CoV-2. J. Virol. 2021, 95, e00083-21. [Google Scholar] [CrossRef] [PubMed]
- Krüger, N.; Rocha, C.; Runft, S.; Krüger, J.; Färber, I.; Armando, F.; Leitzen, E.; Brogden, G.; Gerold, G.; Pöhlmann, S.; et al. The Upper Respiratory Tract of Felids Is Highly Susceptible to SARS-CoV-2 Infection. Int. J. Mol. Sci. 2021, 22, 10636. [Google Scholar] [CrossRef] [PubMed]
- Tomris, I.; Bouwman, K.M.; Adolfs, Y.; Noack, D.; van der Woude, R.; Kerster, G.; Herfst, S.; Sanders, R.; van Gils, M.; Boons, G.-J.; et al. Distinct spatial arrangements of ACE2 and TMPRSS2 expression in Syrian hamster lung lobes dictates SARS-CoV-2 infection patterns. PLoS Pathog. 2020, 18, e1010340. [Google Scholar] [CrossRef]
- Hancock, J.T.; Rouse, R.C.; Stone, E.; Greenhough, A. Interacting proteins, polymorphisms and the susceptibility of animals to sars-cov-2. Animals 2021, 11, 797. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Source | Faeces | Pool Water | |||
|---|---|---|---|---|---|
| Number of Positives | Ct values | Gene | Number of Positives | Ct values | |
| Hippo A Hippo B Pool Water Pool Water Pool Water Pool Water | 3 2 | 35.0–39.2 38.1–39.0 | N1 N2 E Total | 7 6 3 9 | 33.56–35.78 34.78–38.09 36.71–37.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vercammen, F.; Cay, B.; Gryseels, S.; Balmelle, N.; Joffrin, L.; Van Hoorde, K.; Verhaegen, B.; Mathijs, E.; Van Vredendaal, R.; Dharmadhikari, T.; et al. SARS-CoV-2 Infection in Captive Hippos (Hippopotamus amphibius), Belgium. Animals 2023, 13, 316. https://doi.org/10.3390/ani13020316
Vercammen F, Cay B, Gryseels S, Balmelle N, Joffrin L, Van Hoorde K, Verhaegen B, Mathijs E, Van Vredendaal R, Dharmadhikari T, et al. SARS-CoV-2 Infection in Captive Hippos (Hippopotamus amphibius), Belgium. Animals. 2023; 13(2):316. https://doi.org/10.3390/ani13020316
Chicago/Turabian StyleVercammen, Francis, Brigitte Cay, Sophie Gryseels, Nadège Balmelle, Léa Joffrin, Koenraad Van Hoorde, Bavo Verhaegen, Elisabeth Mathijs, Rianne Van Vredendaal, Tanmay Dharmadhikari, and et al. 2023. "SARS-CoV-2 Infection in Captive Hippos (Hippopotamus amphibius), Belgium" Animals 13, no. 2: 316. https://doi.org/10.3390/ani13020316
APA StyleVercammen, F., Cay, B., Gryseels, S., Balmelle, N., Joffrin, L., Van Hoorde, K., Verhaegen, B., Mathijs, E., Van Vredendaal, R., Dharmadhikari, T., Chiers, K., Van Olmen, T. J. S., Agliani, G., Van den Brand, J. M. A., & Leirs, H. (2023). SARS-CoV-2 Infection in Captive Hippos (Hippopotamus amphibius), Belgium. Animals, 13(2), 316. https://doi.org/10.3390/ani13020316