


The Application of Synthetic Flavors in Zebrafish (Danio rerio) Rearing with Emphasis on Attractive Ones: Effects on Fish Development, Welfare, and Appetite

 , , ,
, , ,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Synthetic Flavors
2.3. Preliminary Behavioral Preference Test
2.4. Production of Experimental Diets
2.5. Experimental Design
2.6. Biometry
2.7. Feed Intake
2.8. Histological Analysis
2.9. Molecular Analyses
2.10. Total Lipid Analysis
2.11. Statistical Analyses
3. Results
3.1. Behavioral Preference Test
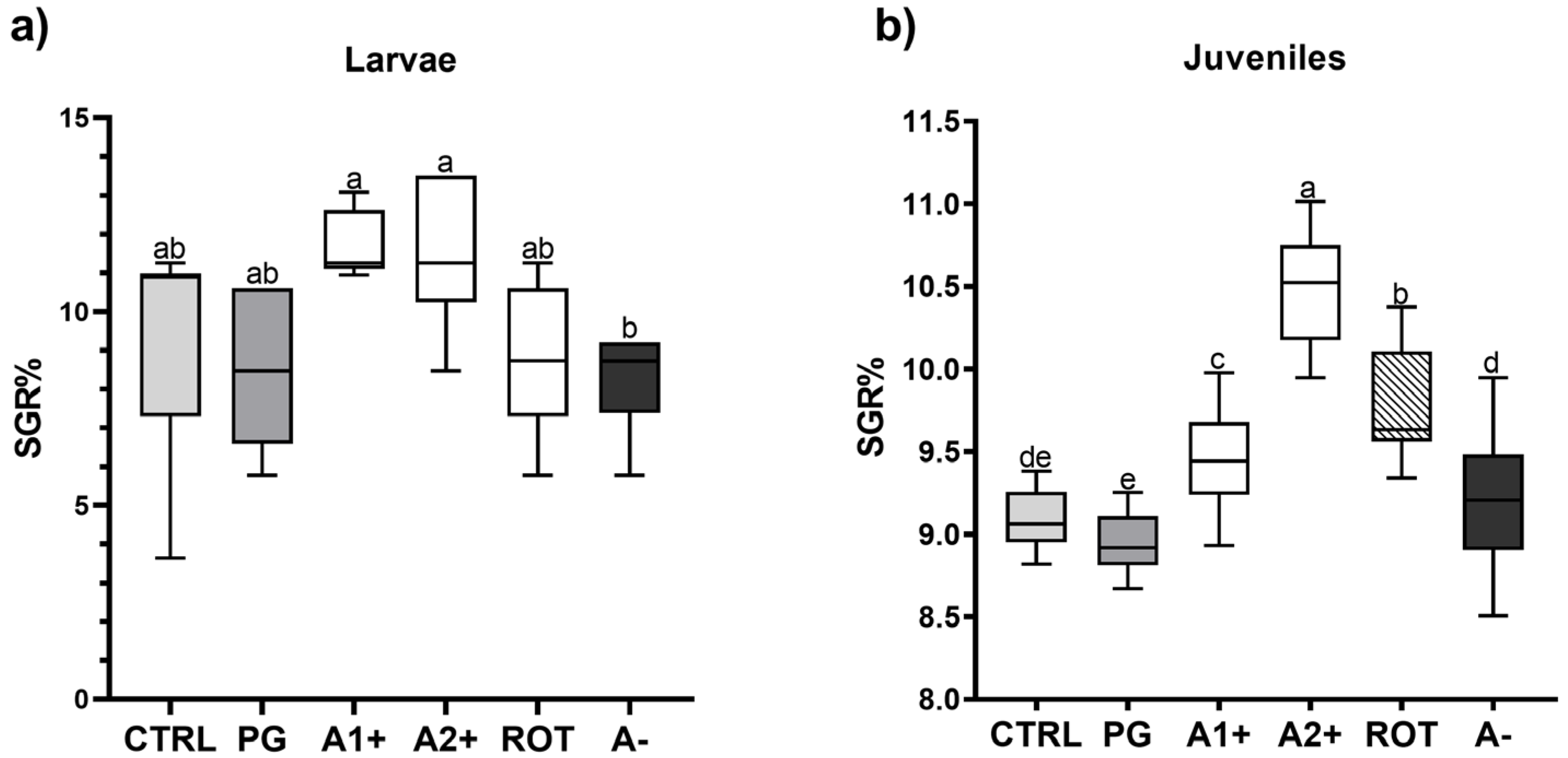
3.2. Biometry
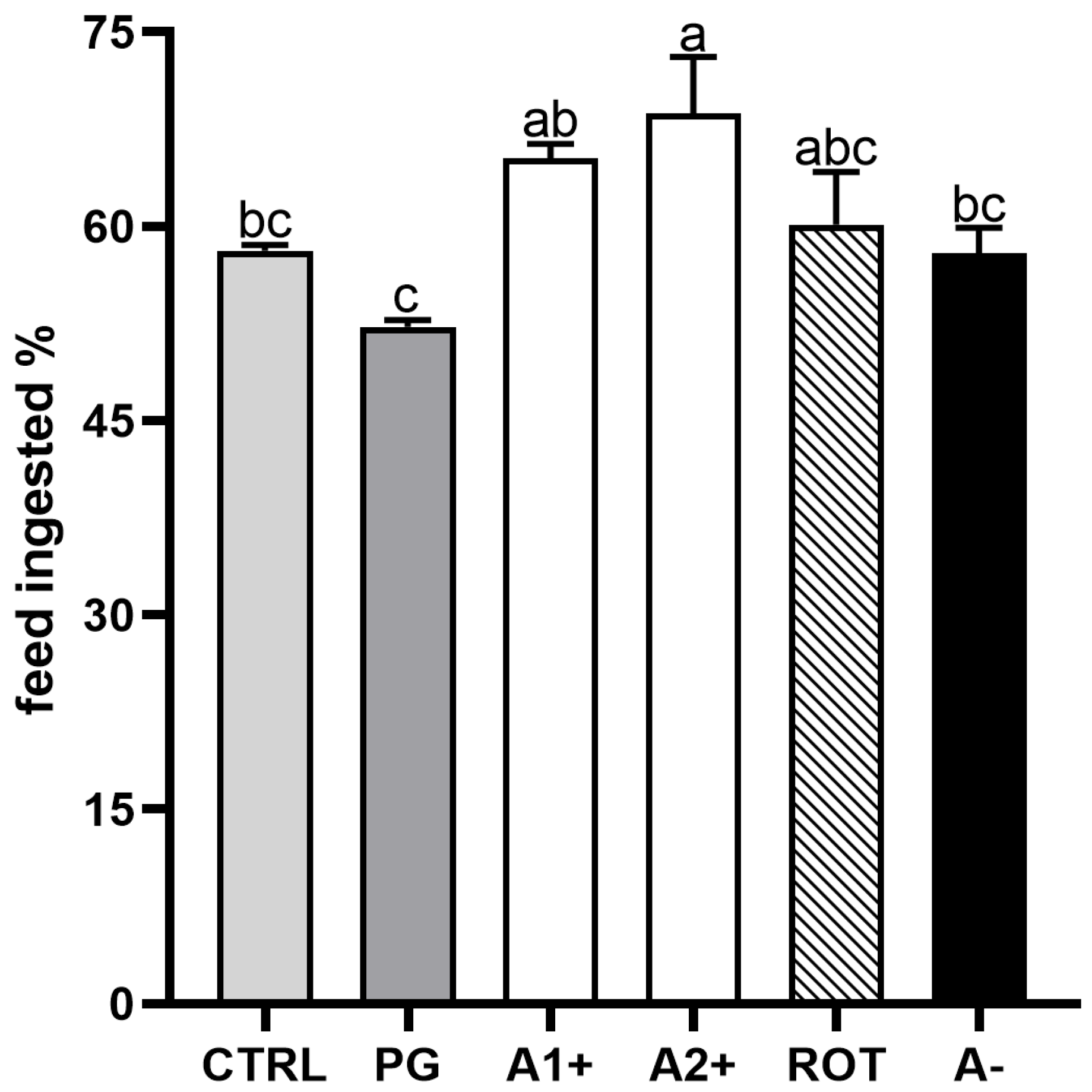
3.3. Feed Intake
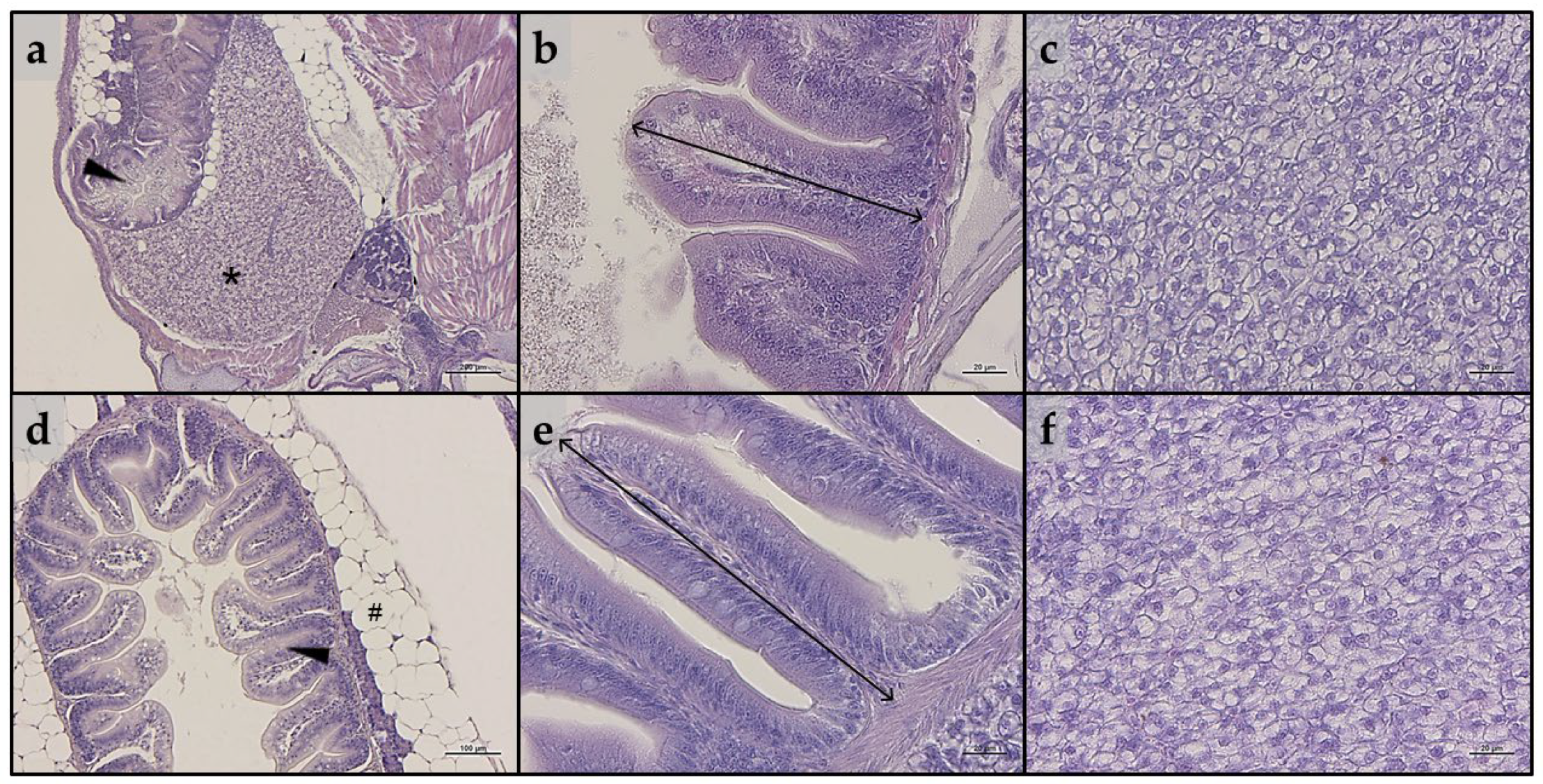
3.4. Histology
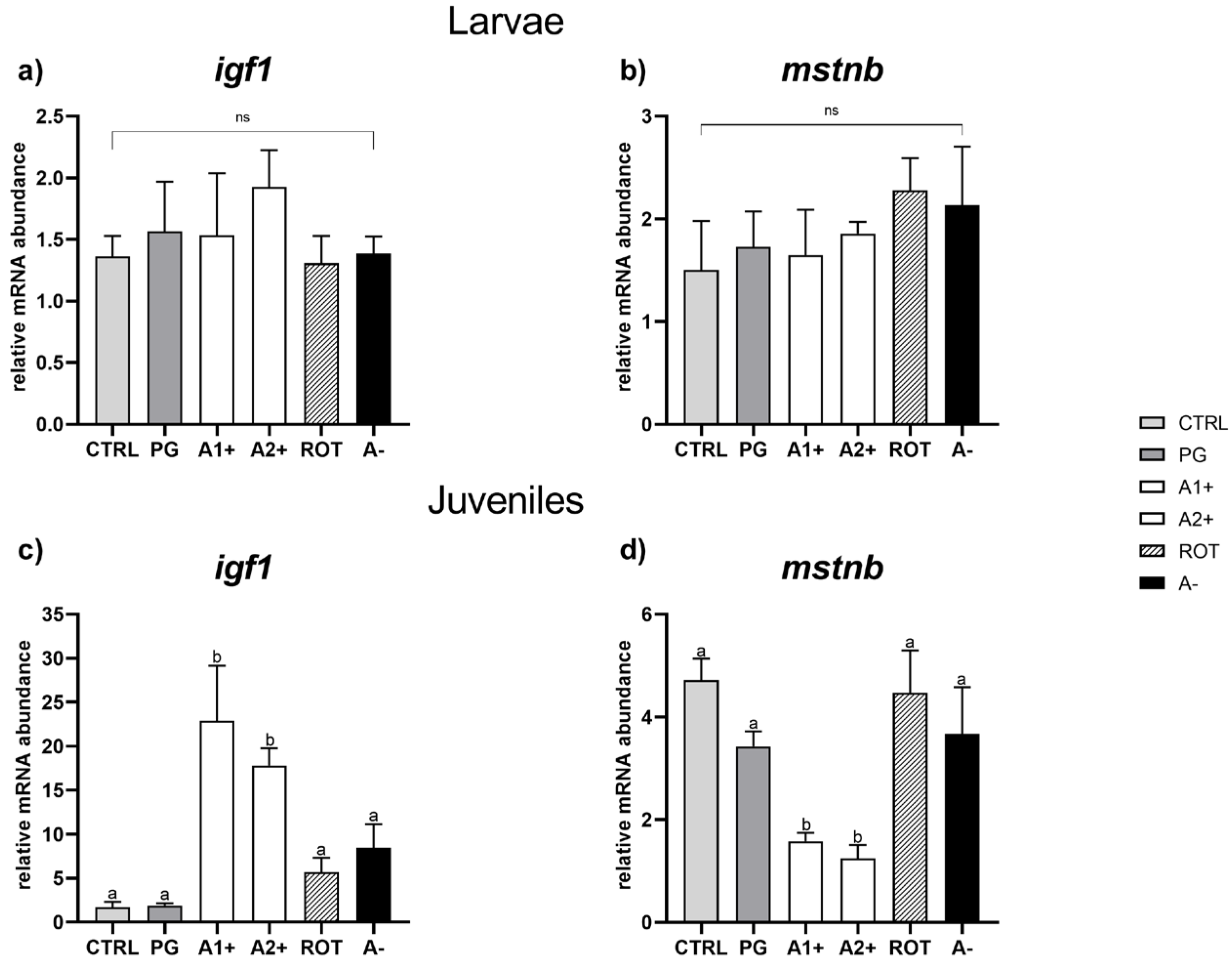
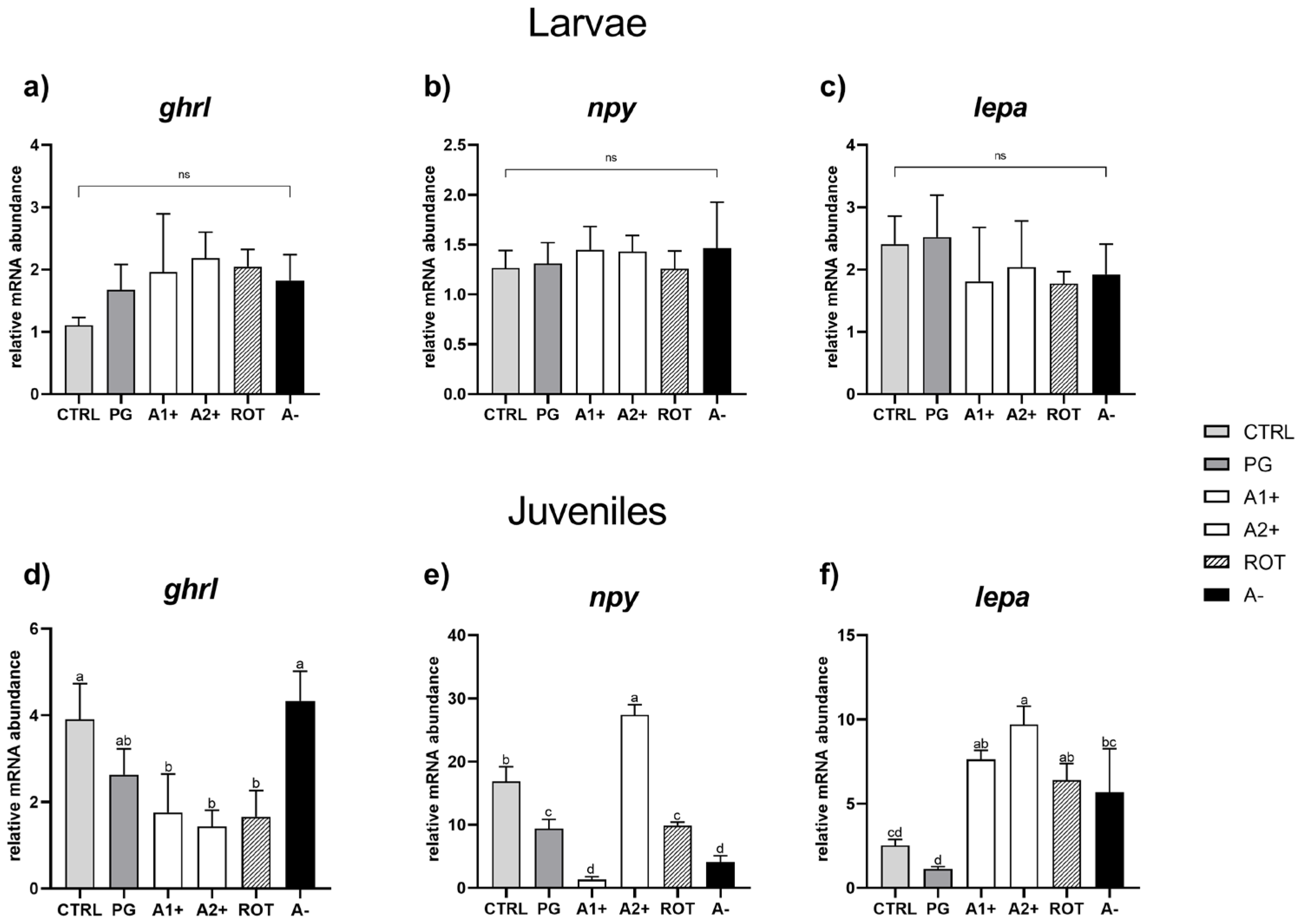
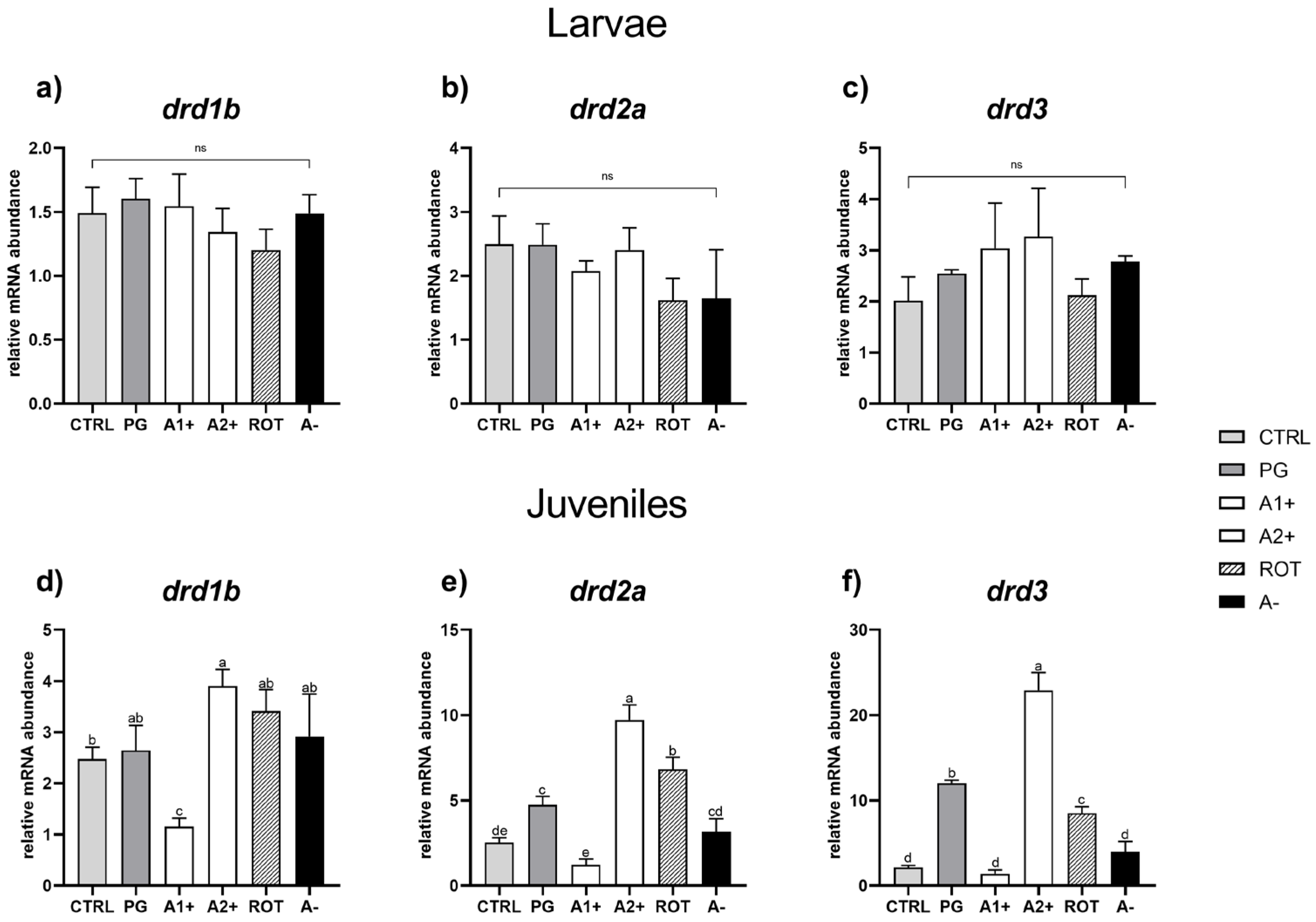
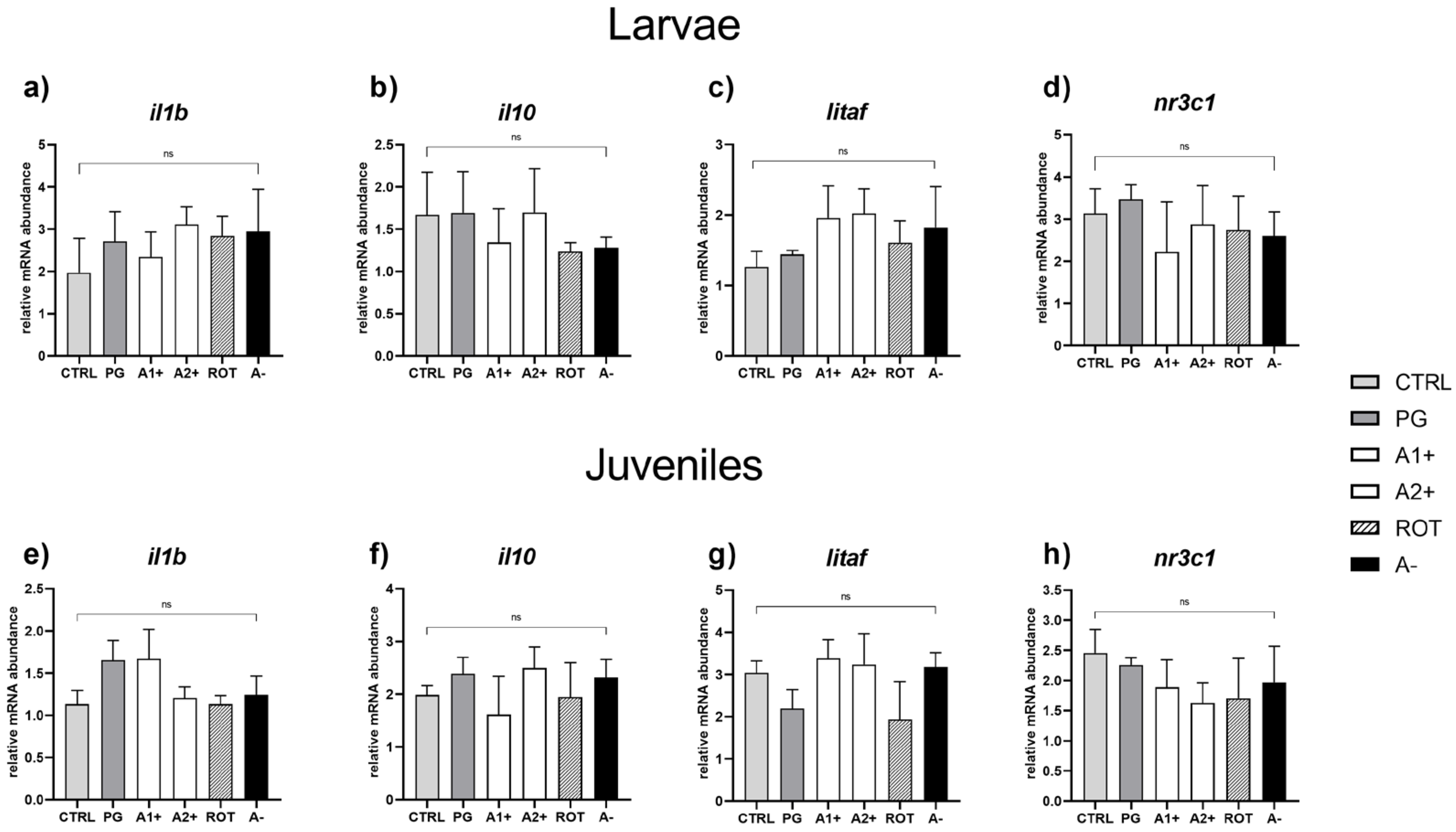
3.5. Real-Time PCR Results
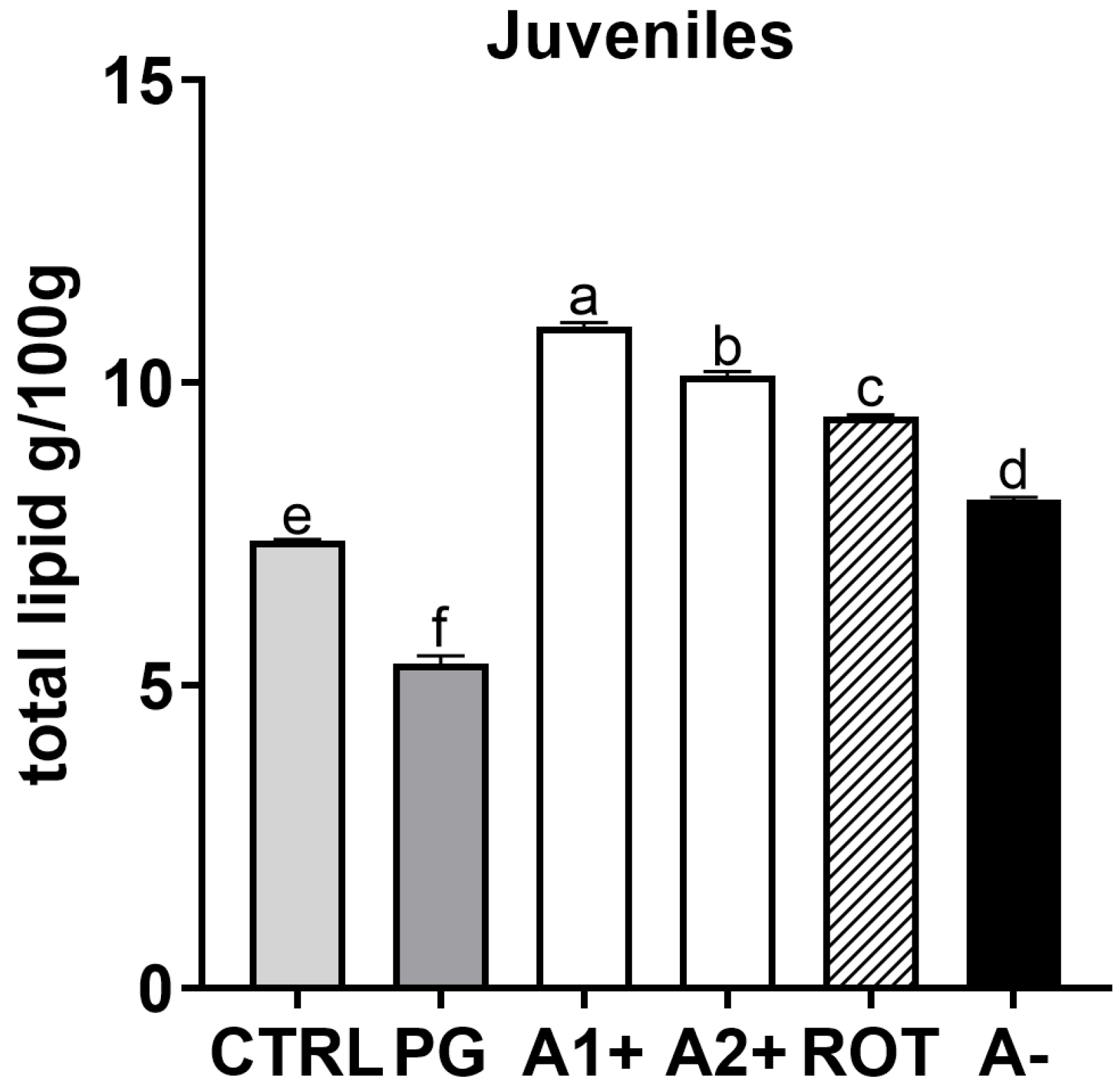
3.6. Total Lipid Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. In Brief to the State of World Fisheries and Aquaculture 2022. Towards Blue Transformation. 2022. Available online: https://www.fao.org/documents/card/en/c/cc0463en (accessed on 27 July 2023).
- Edwards, P. Aquaculture Environment Interactions: Past, Present and Likely Future Trends. Aquaculture 2015, 447, 2–14. [Google Scholar] [CrossRef]
- Kokou, F.; Fountoulaki, E. Aquaculture Waste Production Associated with Antinutrient Presence in Common Fish Feed Plant Ingredients. Aquaculture 2018, 495, 295–310. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Metian, M. Feed Matters: Satisfying the Feed Demand of Aquaculture. Rev. Fish. Sci. Aquac. 2015, 23, 1–10. [Google Scholar] [CrossRef]
- Estruch, G.; Collado, M.C.; Monge-Ortiz, R.; Tomás-Vidal, A.; Jover-Cerdá, M.; Peñaranda, D.S.; Pérez Martínez, G.; Martínez-Llorens, S. Long-Term Feeding with High Plant Protein Based Diets in Gilthead Seabream (Sparus aurata, L.) Leads to Changes in the Inflammatory and Immune Related Gene Expression at Intestinal Level. BMC Vet. Res. 2018, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Santigosa, E.; García-Meilán, I.; Valentin, J.M.; Pérez-Sánchez, J.; Médale, F.; Kaushik, S.; Gallardo, M.A. Modifications of Intestinal Nutrient Absorption in Response to Dietary Fish Meal Replacement by Plant Protein Sources in Sea Bream (Sparus aurata) and Rainbow Trout (Onchorynchus mykiss). Aquaculture 2011, 317, 146–154. [Google Scholar] [CrossRef]
- Maulu, S.; Langi, S.; Hasimuna, O.J.; Missinhoun, D.; Munganga, B.P.; Hampuwo, B.M.; Gabriel, N.N.; Elsabagh, M.; Van Doan, H.; Abdul Kari, Z.; et al. Recent advances in the utilization of insects as an ingredient in aquafeeds: A review. Anim. Nutr. 2022, 11, 334–349. [Google Scholar] [CrossRef] [PubMed]
- Randazzo, B.; Zarantoniello, M.; Cardinaletti, G.; Cerri, R.; Giorgini, E.; Belloni, A.; Contò, M.; Tibaldi, E.; Olivotto, I. Hermetia illucens and Poultry By-Product Meals as Alternatives to Plant Protein Sources in Gilthead Seabream (Sparus aurata) Diet: A Multidisciplinary Study on Fish Gut Status. Animals 2021, 11, 677. [Google Scholar] [CrossRef] [PubMed]
- Estensoro, I.; Ballester-Lozano, G.; Benedito-Palos, L.; Grammes, F.; Martos-Sitcha, J.A.; Mydland, L.-T.; Calduch-Giner, J.A.; Fuentes, J.; Karalazos, V.; Ortiz, A.; et al. Dietary Butyrate Helps to Restore the Intestinal Status of a Marine Teleost (Sparus aurata) Fed Extreme Diets Low in Fish Meal and Fish Oil. PLoS ONE 2016, 11, e0166564. [Google Scholar] [CrossRef]
- Alves, D.R.S.; de Oliveira, S.R.; Luczinski, T.G.; Paulo, I.G.P.; Boscolo, W.R.; Bittencourt, F.; Signor, A. Palatability of Protein Hydrolysates from Industrial Byproducts for Nile Tilapia Juveniles. Animals 2019, 9, 311. [Google Scholar] [CrossRef]
- Carvalho, M.; Torrecillas, S.; Montero, D.; Sanmartín, A.; Fontanillas, R.; Farías, A.; Moutou, K.; Velásquez, J.H.; Izquierdo, M. Insect and Single-Cell Protein Meals as Replacers of Fish Meal in Low Fish Meal and Fish Oil Diets for Gilthead Sea Bream (Sparus aurata) Juveniles. Aquaculture 2023, 566, 739215. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Gioacchini, G.; Cardinaletti, G.; Belloni, A.; Giorgini, E.; Faccenda, F.; Cerri, R.; Tibaldi, E.; Olivotto, I. Physiological Response of Rainbow Trout (Oncorhynchus mykiss) to Graded Levels of Hermetia illucens or Poultry by-Product Meals as Single or Combined Substitute Ingredients to Dietary Plant Proteins. Aquaculture 2021, 538, 736550. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Pulido Rodriguez, L.F.; Randazzo, B.; Cardinaletti, G.; Giorgini, E.; Belloni, A.; Secci, G.; Faccenda, F.; Pulcini, D.; Parisi, G.; et al. Conventional Feed Additives or Red Claw Crayfish Meal and Dried Microbial Biomass as Feed Supplement in Fish Meal-Free Diets for Rainbow Trout (Oncorhynchus mykiss): Possible Ameliorative Effects on Growth and Gut Health Status. Aquaculture 2022, 554, 738137. [Google Scholar] [CrossRef]
- Randazzo, B.; Di Marco, P.; Zarantoniello, M.; Daniso, E.; Cerri, R.; Finoia, M.G.; Capoccioni, F.; Tibaldi, E.; Olivotto, I.; Cardinaletti, G. Effects of Supplementing a Plant Protein-Rich Diet with Insect, Crayfish or Microalgae Meals on Gilthead Sea Bream (Sparus aurata) and European Seabass (Dicentrarchus labrax) Growth, Physiological Status and Gut Health. Aquaculture 2023, 575, 739811. [Google Scholar] [CrossRef]
- Zlaugotne, B.; Pubule, J.; Blumberga, D. Advantages and Disadvantages of Using More Sustainable Ingredients in Fish Feed. Heliyon 2022, 8, e10527. [Google Scholar] [CrossRef]
- Ballester-Moltó, M.; Sanchez-Jerez, P.; Cerezo-Valverde, J.; Aguado-Giménez, F. Particulate Waste Outflow from Fish-Farming Cages. How Much Is Uneaten Feed? Mar. Pollut. Bull. 2017, 119, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Huang, S.; Yang, Z.; Shi, F.; Feng, Y.; Khatoon, Z. Fish Feed Quality Is a Key Factor in Impacting Aquaculture Water Environment: Evidence from Incubator Experiments. Sci. Rep. 2020, 10, 187. [Google Scholar] [CrossRef]
- Amirkolaie, A.K. Reduction in the Environmental Impact of Waste Discharged by Fish Farms through Feed and Feeding. Rev. Aquac. 2011, 3, 19–26. [Google Scholar] [CrossRef]
- Jeong, H.S.; Kim, J.; Olowe, O.S.; Cho, S.H. Dietary Optimum Inclusion Level of Jack Mackerel Meal for Olive Flounder (Paralichthys olivaceus, Temminck & Schlegel, 1846). Aquaculture 2022, 559, 738432. [Google Scholar] [CrossRef]
- Kasumyan, A.O. The Taste System in Fishes and the Effects of Environmental Variables. J. Fish Biol. 2019, 95, 155–178. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H. The Neuroendocrine Regulation of Food Intake in Fish: A Review of Current Knowledge. Front. Neurosci. 2016, 10, 540. [Google Scholar] [CrossRef]
- Bertucci, J.I.; Blanco, A.M.; Sundarrajan, L.; Rajeswari, J.J.; Velasco, C.; Unniappan, S. Nutrient Regulation of Endocrine Factors Influencing Feeding and Growth in Fish. Front. Endocrinol. 2019, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Rønnestad, I.; Gomes, A.S.; Murashita, K.; Angotzi, R.; Jönsson, E.; Volkoff, H. Appetite-Controlling Endocrine Systems in Teleosts. Front. Endocrinol. 2017, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Flik, G.; Huising, M.; Gorissen, M. Peptides and Proteins Regulating Food Intake: A Comparative View. Anim. Biol. 2006, 56, 447–473. [Google Scholar] [CrossRef]
- Gong, N.; Jönsson, E.; Björnsson, B.T. Acute Anorexigenic Action of Leptin in Rainbow Trout Is Mediated by the Hypothalamic Pi3k Pathway. J. Mol. Endocrinol. 2016, 56, 227–238. [Google Scholar] [CrossRef]
- Soengas, J.L.; Cerdá-Reverter, J.M.; Delgado, M.J. Central Regulation of Food Intake in Fish: An Evolutionary Perspective. J. Mol. Endocrinol. 2018, 60, R171–R199. [Google Scholar] [CrossRef] [PubMed]
- de Macedo, I.C.; de Freitas, J.S.; da Silva Torres, I.L. The Influence of Palatable Diets in Reward System Activation: A Mini Review. Adv. Pharmacol. Sci. 2016, 2016, 7238679. [Google Scholar] [CrossRef]
- Hancz, C. Feed Efficiency, Nutrient Sensing and Feeding Stimulation in Aquaculture: A Review. Acta Agrar. Kaposváriensis 2020, 24, 35–54. [Google Scholar] [CrossRef]
- Al-Souti, A.; Gallardo, W.; Claereboudt, M.; Mahgoub, O. Attractability and Palatability of Formulated Diets Incorporated with Chicken Feather and Algal Meals for Juvenile Gilthead Seabream, Sparus aurata. Aquac. Rep. 2019, 14, 100199. [Google Scholar] [CrossRef]
- Nagel, F.; von Danwitz, A.; Schlachter, M.; Kroeckel, S.; Wagner, C.; Schulz, C. Blue Mussel Meal as Feed Attractant in Rapeseed Protein-Based Diets for Turbot (Psetta maxima, L.). Aquac. Res. 2014, 45, 1964–1978. [Google Scholar] [CrossRef]
- Kader, M.A.; Bulbul, M.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Nguyen, B.T.; Komilus, C.F. Effect of Complete Replacement of Fishmeal by Dehulled Soybean Meal with Crude Attractants Supplementation in Diets for Red Sea Bream, Pagrus major. Aquaculture 2012, 350, 109–116. [Google Scholar] [CrossRef]
- Ytrestøyl, T.; Aas, T.S.; Åsgård, T. Utilisation of Feed Resources in Production of Atlantic Salmon (Salmo salar) in Norway. Aquaculture 2015, 448, 365–374. [Google Scholar] [CrossRef]
- Bureau, D.P.; Harris, A.M.; Bevan, D.J.; Simmons, L.A.; Azevedo, P.A.; Cho, C.Y. Feather Meals and Meat and Bone Meals from Different Origins as Protein Sources in Rainbow Trout (Oncorhynchus mykiss) Diets. Aquaculture 2000, 181, 281–291. [Google Scholar] [CrossRef]
- Peng, D.; Peng, B.; Li, J.; Zhang, Y.; Luo, H.; Xiao, Q.; Tang, S.; Liang, X.-F. Effects of Three Feed Attractants on the Growth, Biochemical Indicators, Lipid Metabolism and Appetite of Chinese Perch (Siniperca chuatsi). Aquac. Rep. 2022, 23, 101075. [Google Scholar] [CrossRef]
- Pamungkas, G.Y.; Arief, M.; Arif, M.A.A. The Influence of Garlic (Allium sativum) and Turmeric (Curcuma longa) Extract as Attractant in Commercial Feed on Feeding Rate and Fat Retention in Gouramy (Osphronemus gouramy). In Proceedings of the IOP Conference Series: Earth and Environmental Science: The 1st International Conference on Biotechnology and Food Sciences, Surabaya, Indonesia, 11 September 2020; IOP Publishing: Bristol, UK, 2021; Volume 679, p. 12076. [Google Scholar] [CrossRef]
- Tusche, K.; Berends, K.; Wuertz, S.; Susenbeth, A.; Schulz, C. Evaluation of Feed Attractants in Potato Protein Concentrate Based Diets for Rainbow Trout (Oncorhynchus mykiss). Aquaculture 2011, 321, 54–60. [Google Scholar] [CrossRef]
- Kim, H.S.; Cho, S.H. Dietary Inclusion Effect of Feed Ingredients Showing High Feeding Attractiveness to Rockfish (Sebastes schlegeli, Hilgendorf 1880) on the Growth Performance, Feed Utilization, Condition Factor and Whole Body Composition of Fish (II). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 231, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fang, T.; Wang, J.; Wang, Z.; Guan, D.; Sun, H.; Yun, X.; Zhou, J. The Efficiency of Adding Amino Acid Mixtures to a Diet Free of Fishmeal and Soybean Meal as an Attractant in Yellow River Carp (Cyprinus carpio Var.). Aquac. Rep. 2022, 24, 101189. [Google Scholar] [CrossRef]
- Reig, L.; Ginovart, M.; Flos, R. Modification of the Feeding Behaviour of Sole (Solea sole) through the Addition of a Commercial Flavour as an Alternative to Betaine. Aquat. Living Resour. 2003, 16, 370–379. [Google Scholar] [CrossRef]
- Januszewska, R.; Giret, E.; Clement, F.; Van Leuven, I.; Goncalves, C.; Vladislavleva, E.; Pradal, P.; Nåbo, R.; Landuyt, A.; D’Heer, G.; et al. Impact of Vanilla Origins on Sensory Characteristics of Chocolate. Food Res. Int. 2020, 137, 109313. [Google Scholar] [CrossRef] [PubMed]
- Ulloa, A.M. The Aesthetic Life of Artificial Flavors. Senses Soc. 2018, 13, 60–74. [Google Scholar] [CrossRef]
- Torres, S.; Pandey, A.; Castro, G.R. Banana Flavor: Insights into Isoamyl Acetate Production. Cell 2010, 549, 776. [Google Scholar]
- Ribas, L.; Piferrer, F. The Zebrafish (Danio rerio) as a Model Organism, with Emphasis on Applications for Finfish Aquaculture Research. Rev. Aquac. 2014, 6, 209–240. [Google Scholar] [CrossRef]
- Ulloa, P.E.; Iturra, P.; Neira, R.; Araneda, C. Zebrafish as a Model Organism for Nutrition and Growth: Towards Comparative Studies of Nutritional Genomics Applied to Aquacultured Fishes. Rev. Fish Biol. Fish. 2011, 21, 649–666. [Google Scholar] [CrossRef]
- Dahm, R.; Geisler, R. Learning from Small Fry: The Zebrafish as a Genetic Model Organism for Aquaculture Fish Species. Mar. Biotechnol. 2006, 8, 329–345. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Gioacchini, G.; Truzzi, C.; Giorgini, E.; Riolo, P.; Gioia, G.; Bertolucci, C.; Osimani, A.; Cardinaletti, G.; et al. Zebrafish (Danio rerio) Physiological and Behavioural Responses to Insect-Based Diets: A Multidisciplinary Approach. Sci. Rep. 2020, 10, 10648. [Google Scholar] [CrossRef]
- Lucon-Xiccato, T.; De Russi, G.; Bertolucci, C. A Novel-Odour Exploration Test for Measuring Anxiety in Adult and Larval Zebrafish. J. Neurosci. Methods 2020, 335, 108619. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Randazzo, B.; Cardinaletti, G.; Truzzi, C.; Chemello, G.; Riolo, P.; Olivotto, I. Possible Dietary Effects of Insect-Based Diets across Zebrafish (Danio rerio) Generations: A Multidisciplinary Study on the Larval Phase. Animals 2021, 11, 751. [Google Scholar] [CrossRef]
- Cattaneo, N.; Zarantoniello, M.; Conti, F.; Frontini, A.; Chemello, G.; Dimichino, B.; Marongiu, F.; Cardinaletti, G.; Gioacchini, G.; Olivotto, I. Dietary Microplastic Administration during Zebrafish (Danio rerio) Development: A Comprehensive and Comparative Study between Larval and Juvenile Stages. Animals 2023, 13, 2256. [Google Scholar] [CrossRef] [PubMed]
- Zufall, F.; Leinders-Zufall, T. The Cellular and Molecular Basis of Odor Adaptation. Chem. Senses 2000, 25, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Olivotto, I.; Yasumasu, S.; Gioacchini, G.; Maradonna, F.; Cionna, C.; Carnevali, O. Cloning and Expression of High Choriolytic Enzyme, a Component of the Hatching Enzyme System, during Embryonic Development of the Marine Ornamental Fish Chrysiptera Parasema. Mar. Biol. 2004, 145, 1235–1241. [Google Scholar] [CrossRef]
- Rashidian, G.; Moghaddam, M.M.; Mirnejad, R.; Azad, Z.M. Supplementation of Zebrafish (Danio rerio) Diet Using a Short Antimicrobial Peptide: Evaluation of Growth Performance, Immunomodulatory Function, Antioxidant Activity, and Disease Resistance. Fish Shellfish Immunol. 2021, 119, 42–50. [Google Scholar] [CrossRef]
- Randazzo, B.; Rolla, L.; Ofelio, C.; Planas, M.; Gioacchini, G.; Vargas, A.; Giorgini, E.; Olivotto, I. The Influence of Diet on the Early Development of Two Seahorse Species (H. guttulatus and H. reidi): Traditional and Innovative Approaches. Aquaculture 2018, 490, 75–90. [Google Scholar] [CrossRef]
- Olivotto, I.; Tokle, N.E.; Nozzi, V.; Cossignani, L.; Carnevali, O. Preserved Copepods as a New Technology for the Marine Ornamental Fish Aquaculture: A Feeding Study. Aquaculture 2010, 308, 124–131. [Google Scholar] [CrossRef]
- Randazzo, B.; Zarantoniello, M.; Gioacchini, G.; Giorgini, E.; Truzzi, C.; Notarstefano, V.; Cardinaletti, G.; Huyen, K.T.; Carnevali, O.; Olivotto, I. Can Insect-Based Diets Affect Zebrafish (Danio rerio) Reproduction? A Multidisciplinary Study. Zebrafish 2020, 17, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A Simple Method for the Isolation and Purification of Total Lipids from Animal Tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Enes, P.; Peres, H. Replacing Fishmeal and Fish Oil in Industrial Aquafeeds for Carnivorous Fish. In Feed and Feeding Practices in Aquaculture; Elsevier: Amsterdam, The Netherlands, 2015; pp. 203–233. ISBN 9780081005064. [Google Scholar] [CrossRef]
- Hilton, J.W.; Atkinson, J.L.; Slinger, S.J. Effect of Propylene Glycol on Feed Digestibility and the Growth and Physiological Response of Rainbow Trout. Can. J. Anim. Sci. 1986, 66, 1057–1063. [Google Scholar] [CrossRef]
- Soaudy, M.R.; Mohammady, E.Y.; Elashry, M.A.; Ali, M.M.; Ahmed, N.M.; Hegab, M.H.; El-Garhy, H.A.S.; El-Haroun, E.R.; Hassaan, M.S. Possibility Mitigation of Cold Stress in Nile Tilapia under Biofloc System by Dietary Propylene Glycol: Performance Feeding Status, Immune, Physiological Responses and Transcriptional Response of Delta-9-Desaturase Gene. Aquaculture 2021, 538, 736519. [Google Scholar] [CrossRef]
- Smith, R.L.; Cohen, S.M.; Fukushima, S.; Gooderham, N.J.; Hecht, S.S.; Guengerich, F.P.; Rietjens, I.M.C.M.; Bastaki, M.; Harman, C.L.; McGowen, M.M.; et al. The Safety Evaluation of Food Flavouring Substances: The Role of Metabolic Studies. Toxicol. Res. (Camb) 2018, 7, 618–646. [Google Scholar] [CrossRef] [PubMed]
- Olivotto, I.; Mosconi, G.; Maradonna, F.; Cardinali, M.; Carnevali, O. Diplodus Sargus Interrenal—Pituitary Response: Chemical Communication in Stressed Fish. Gen. Comp. Endocrinol. 2002, 127, 66–70. [Google Scholar] [CrossRef]
- Opazo Salas, R.; Valladares Boasi, L.; Romero Ormazábal, J. Comparison of Gene Expression Patterns of Key Growth Genes between Different Rate Growths in Zebrafish (Danio rerio) Siblings. Lat. Am. J. Aquat. Res. 2017, 45, 766–775. [Google Scholar] [CrossRef]
- Hamre, K.; Yufera, M.; Rønnestad, I.; Boglione, C.; Conceição, L.E.C.; Izquierdo, M. Fish Larval Nutrition and Feed Formulation: Knowledge Gaps and Bottlenecks for Advances in Larval Rearing. Rev. Aquac. 2013, 5, S26–S58. [Google Scholar] [CrossRef]
- Volkoff, H. Fish as Models for Understanding the Vertebrate Endocrine Regulation of Feeding and Weight. Mol. Cell. Endocrinol. 2019, 497, 110437. [Google Scholar] [CrossRef] [PubMed]
- Kwasek, K.; Wojno, M.; Iannini, F.; McCracken, V.J.; Molinari, G.S.; Terova, G. Nutritional Programming Improves Dietary Plant Protein Utilization in Zebrafish Danio rerio. PLoS ONE 2020, 15, e0225917. [Google Scholar] [CrossRef] [PubMed]
- Cerdá-Reverter, J.M.; Larhammar, D. Neuropeptide Y Family of Peptides: Structure, Anatomical Expression, Function, and Molecular Evolution. Biochem. Cell Biol. 2000, 78, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Campos, V.F.; Collares, T.; Deschamps, J.C.; Seixas, F.K.; Dellagostin, O.A.; Lanes, C.F.C.; Sandrini, J.; Marins, L.F.; Okamoto, M.; Sampaio, L.A.; et al. Identification, Tissue Distribution and Evaluation of Brain Neuropeptide Y Gene Expression in the Brazilian Flounder Paralichthys orbignyanus. J. Biosci. 2010, 35, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Volkoff, H.; Canosa, L.F.; Unniappan, S.; Cerdá-Reverter, J.M.; Bernier, N.J.; Kelly, S.P.; Peter, R.E. Neuropeptides and the Control of Food Intake in Fish. Gen. Comp. Endocrinol. 2005, 142, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Opazo, R.; Plaza-Parrochia, F.; dos Santos, G.R.; Carneiro, G.R.A.; Sardela, V.F.; Romero, J.; Valladares, L. Fasting Upregulates Npy, Agrp, and Ghsr without Increasing Ghrelin Levels in Zebrafish (Danio rerio) Larvae. Front. Physiol. 2019, 9, 1901. [Google Scholar] [CrossRef]
- Giaquinto, D.; De Felice, E.; Attanasio, C.; Palladino, A.; Schiano, V.; Mollo, E.; Lucini, C.; de Girolamo, P.; D’Angelo, L. Central and Peripheral NPY Age-Related Regulation: A Comparative Analysis in Fish Translational Models. Int. J. Mol. Sci. 2022, 23, 3839. [Google Scholar] [CrossRef]
- Matsuda, K.; Sakashita, A.; Yokobori, E.; Azuma, M. Neuroendocrine Control of Feeding Behavior and Psychomotor Activity by NeuropeptideY in Fish. Neuropeptides 2012, 46, 275–283. [Google Scholar] [CrossRef]
- Baik, J.-H. Dopamine Signaling in Reward-Related Behaviors. Front. Neural Circuits 2013, 7, 152. [Google Scholar] [CrossRef]
- Thörnqvist, P.-O.; McCarrick, S.; Ericsson, M.; Roman, E.; Winberg, S. Bold Zebrafish (Danio rerio) Express Higher Levels of Delta Opioid and Dopamine D2 Receptors in the Brain Compared to Shy Fish. Behav. Brain Res. 2019, 359, 927–934. [Google Scholar] [CrossRef]
- Le Crom, S.; Kapsimali, M.; Barôme, P.-O.; Vernier, P. Dopamine Receptors for Every Species: Gene Duplications and Functional Diversification in Craniates. J. Struct. Funct. Genom. 2003, 3, 161–176. [Google Scholar] [CrossRef]
- Alsiö, J.; Olszewski, P.K.; Norbäck, A.H.; Gunnarsson, Z.E.A.; Levine, A.S.; Pickering, C.; Schiöth, H.B. Dopamine D1 Receptor Gene Expression Decreases in the Nucleus Accumbens upon Long-Term Exposure to Palatable Food and Differs Depending on Diet-Induced Obesity Phenotype in Rats. Neuroscience 2010, 171, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.M.; Kenny, P.J. Dopamine D2 Receptors in Addiction-like Reward Dysfunction and Compulsive Eating in Obese Rats. Nat. Neurosci. 2010, 13, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wise, R.A.; Baler, R. The Dopamine Motive System: Implications for Drug and Food Addiction. Nat. Rev. Neurosci. 2017, 18, 741–752. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | AT (°C) | ID |
|---|---|---|---|---|
| igf1 | GGCAAATCTCCACGATCTCTAC | CGGTTTCTCTTGTCTCTCTCAG | 53 | ZDB-GENE-010607-2 |
| mstnb | GGACTGGACTGCGATGAG | GATGGGTGTGGGGATACTTC | 58 | ZDB-GENE-990415-165 |
| ghrl | CAGCATGTTTCTGCTCCTGTG | TCTTCTGCCCACTCTTGGTG | 58 | ZDB-GENE-070622-2 |
| npy | GTCTGCTTGGGGACTCTCAC | CGGGACTCTGTTTCACCAAT | 60 | ZDB-GENE-980526-438 |
| lepa | CTCCAGTGACGAAGGCAACTT | GGGAAGGAGCCGGAAATGT | 60 | ZDB-GENE-081001-1 |
| drd1b | CTGCGACTCCAGCCTTAATC | AGATGCGGGTGTAAGTGACC | 58 | ZDB-GENE-070524-2 |
| drd2a | TGGTACTCCGGAAAAGACG | ATCGGGATGGGTGCATTTC | 58 | ZDB-GENE-021119-2 |
| drd3 | ATCAGTATCGACAGGTATACAGC | CCAAACAGTAGAGGGCAGG | 60 | ZDB-GENE-021119-1 |
| nr3c1 | AGACCTTGGTCCCCTTCACT | CGCCTTTAATCATGGGAGAA | 58 | ZDB-GENE-050522-503 |
| il1b | GCTGGGGATGTGGACTTC | GTGGATTGGGGTTTGATGTG | 54 | ZDB-GENE-040702-2 |
| il10 | ATTTGTGGAGGGCTTTCCTT | AGAGCTGTTGGCAGAATGGT | 56 | ZDB-GENE-051111-1 |
| litaf | TTGTGGTGGGGTTTGATG | TTGGGGCATTTTATTTTGTAAG | 53 | ZDB-GENE-040704-23 |
| rpl13 | TCTGGAGGACTGTAAGAGGTATGC | AGACGCACAATCTTGAGAGCAG | 59 | ZDB-GENE-031007-1 |
| arpc1a | CTGAACATCTCGCCCTTCTC | TAGCCGATCTGCAGACACAC | 60 | ZDB-GENE-040116-1 |
| CTRL | PG | A1+ | A2+ | ROT | A− | |
|---|---|---|---|---|---|---|
| Larvae | ||||||
| Mucosal fold height | 80.7 ± 0.6 | 93.0 ± 7.2 | 84.5 ± 5.5 | 95.4 ± 7.5 | 92.5 ± 7.2 | 75.0 ± 4.5 |
| Inflammatory influx | + | + | + | + | + | + |
| Mucosal fold fusion | + | + | + | + | + | + |
| Juveniles | ||||||
| Mucosal fold height | 166.2 ± 23.1 | 168.0 ± 23.0 | 160.0 ± 12.0 | 175.3 ± 26.2 | 162.4 ± 12.7 | 164.7 ± 25.5 |
| Inflammatory influx | + | + | + | + | + | + |
| Mucosal fold fusion | + | + | + | + | + | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conti, F.; Zarantoniello, M.; Antonucci, M.; Cattaneo, N.; Rattin, M.; De Russi, G.; Secci, G.; Lucon-Xiccato, T.; Lira de Medeiros, A.C.; Olivotto, I. The Application of Synthetic Flavors in Zebrafish (Danio rerio) Rearing with Emphasis on Attractive Ones: Effects on Fish Development, Welfare, and Appetite. Animals 2023, 13, 3368. https://doi.org/10.3390/ani13213368
Conti F, Zarantoniello M, Antonucci M, Cattaneo N, Rattin M, De Russi G, Secci G, Lucon-Xiccato T, Lira de Medeiros AC, Olivotto I. The Application of Synthetic Flavors in Zebrafish (Danio rerio) Rearing with Emphasis on Attractive Ones: Effects on Fish Development, Welfare, and Appetite. Animals. 2023; 13(21):3368. https://doi.org/10.3390/ani13213368
Chicago/Turabian StyleConti, Federico, Matteo Zarantoniello, Matteo Antonucci, Nico Cattaneo, Mirko Rattin, Gaia De Russi, Giulia Secci, Tyrone Lucon-Xiccato, Adja Cristina Lira de Medeiros, and Ike Olivotto. 2023. "The Application of Synthetic Flavors in Zebrafish (Danio rerio) Rearing with Emphasis on Attractive Ones: Effects on Fish Development, Welfare, and Appetite" Animals 13, no. 21: 3368. https://doi.org/10.3390/ani13213368
APA StyleConti, F., Zarantoniello, M., Antonucci, M., Cattaneo, N., Rattin, M., De Russi, G., Secci, G., Lucon-Xiccato, T., Lira de Medeiros, A. C., & Olivotto, I. (2023). The Application of Synthetic Flavors in Zebrafish (Danio rerio) Rearing with Emphasis on Attractive Ones: Effects on Fish Development, Welfare, and Appetite. Animals, 13(21), 3368. https://doi.org/10.3390/ani13213368