1. Introduction
Under modern intensive farming conditions, breeder hens are prone to inflammation, fat deposition, autophagy and oxidative stress in the liver [
1]. The influence of nutrition, stress and other factors in the late laying period can alter the homeostasis of liver fat, affect lipid metabolism and easily cause fatty liver bleeding syndrome [
2].
The liver is a major site of lipid biosynthesis and has an important function in controlling systemic glucose and lipid metabolism [
3]. It also supports nutrient absorption and utilization [
4]. The liver is also the site of the synthesis of lipids and proteins necessary for yolk production [
5]. As chickens age, lipid accumulation gradually leads to oxidative stress damage, resulting in reduced antioxidant capacity, impaired liver function and hepatic steatosis [
6,
7,
8]. Due to the diverse functions and vital importance of the liver, any reduction in its function can lead to metabolic disorders and a deterioration in the quality of egg production [
9]. Many factors can cause metabolic diseases in breeder hens, but nutritional factors have been key to the etiology of hapatic lipid metabolism [
10].
Perilla (
Perilla frutescens L.) is an annual plant in the Labiatae family and is widely cultivated as an important crop in East Asian countries [
11]. Perilla seeds (PS) contain numerous active ingredients with different effects, including anti-allergic, antibacterial, anti-inflammatory, hypolipidemic, antioxidant and anticancer properties [
12]. In addition, PS effectively prevents cell aging [
13]. Perilla seed meal (PSM) is a byproduct of PS oil extraction. PSM is a nutrient-rich substance containing large amounts of proteins (35 to 45%), fiber (55 to 65%), and small amounts of polysaccharides, polyols, fatty acids, phytosterols, flavonoids and phenolic compounds [
14,
15]. A growing body of research suggests that plant products such as flavonoids and polyphenols play a crucial role in performing antioxidant and anti-inflammatory functions in animals [
16,
17,
18].
Wenchang chicken features heat resistance and delicious meat quality [
19]. However, it should be noted that the Wenchang chicken is a breed of chicken native in China with relatively low egg production [
20]. The peak laying age of Wenchang chickens is 23 weeks old [
21]. The egg production rate of Wenchang chickens peaks in the 7th or 8th postpartum week, and more than 60% of the egg production rate can be maintained for 15–18 weeks [
22]. Compared to research on the meat quality of Wenchang chickens, there are few studies on their production performance.
Previous studies have found that perilla seed extract (PSE) lowers blood lipids in mice and breeder hens and affects egg quality and fatty acid composition [
23,
24,
25]. PS consumption by the hens could also influence the unsaturated fatty acid content of the eggs and thus improve their nutrition [
26,
27]. Since PSM is rich in high-quality fats and nutrients, these experiments examined whether PSM can affect liver lipid metabolism and antioxidant capacity in breeder hens, as well as the effects of PSM on laying performance, egg quality and fatty acid content of the eggs. The aim of this study was to create a theoretical basis for the application of PSM in egg production.
2. Materials and Methods
2.1. Preparation of Perilla Seed Meal
The PS was manufactured in Weiyuan Town, Changshun County, Qiannan Buyi Miao Autonomou Prefecture, Guizhou Province. PSM was the residual product after pressing the perilla seeds for oil production and roasting them at 120 °C for 30 min in a microcomputer roaster. The nutritional ingredients and active ingredients of the PSM are listed in
Table S1. The nutritious ingredients of PSM have been published in a previous article [
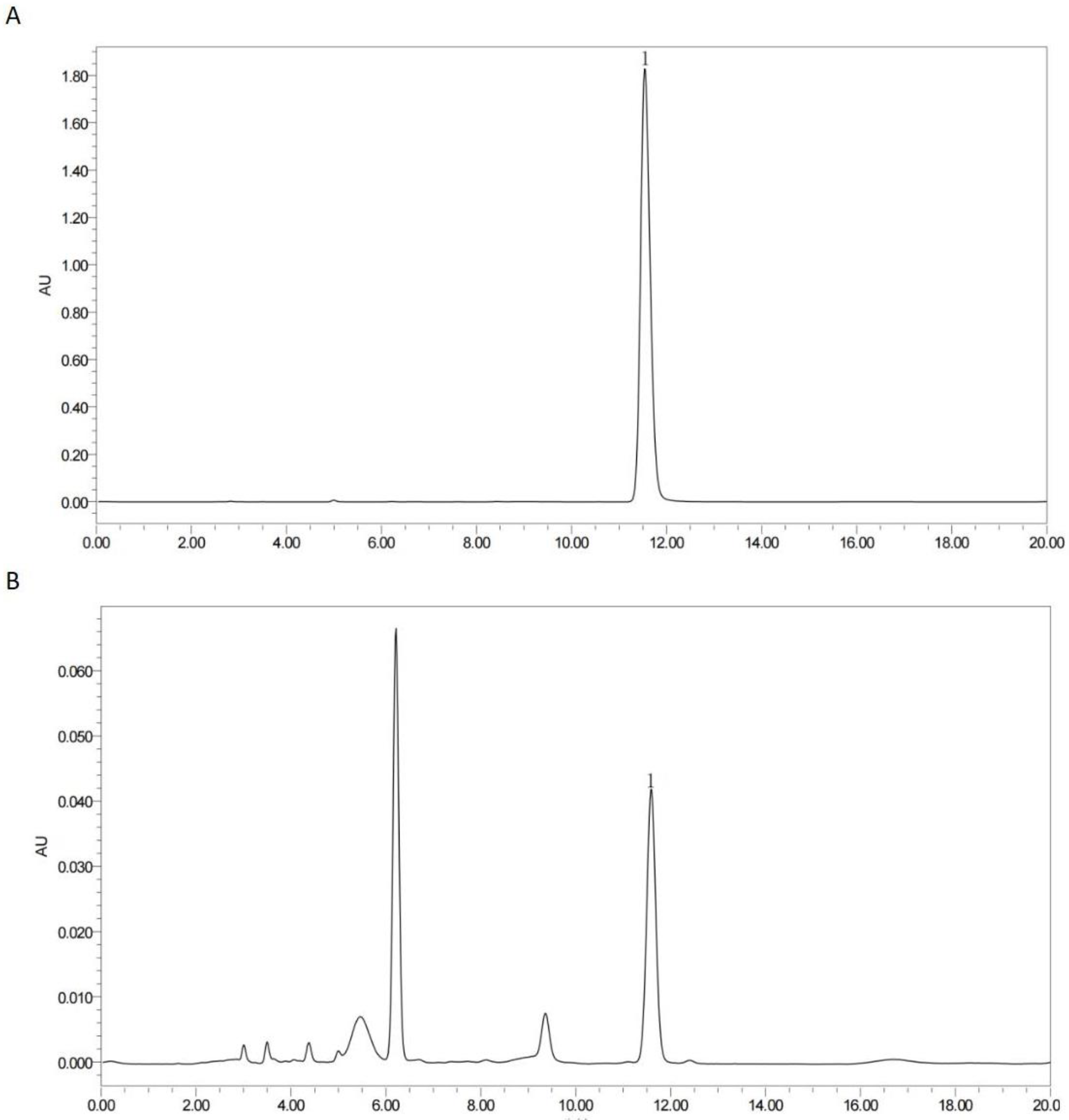
28]. The total polysaccharides of the PSM were determined using the phenol-sulfuric acid method with glucose as a standard. The total polysaccharide content of the PSM was measured to be 1.33%. The determination of the total flavonoids and total phenolics was carried out using colorimetric kits (Jiancheng Bioengineering Institute, Nanjing, China). The total flavonoid and total phenolic contents were 1.32% and 0.44%, respectively. The rosemarinic acid content was measured using Waters Breeze QS high-performance liquid chromatography (HPLC), and the results are shown in
Figure 1. The amount of rosmarinic acid was calculated to be 0.19%.
2.2. Animals, Diet and Experimental Design
A total of 192 Wenchang hens (31 weeks old, 1.65 ± 0.20 kg body weight) with similar laying rates were obtained. All experimental protocols were approved by the Animal Care and Use Committee of the South China Agricultural University (approval number: SYXK 2019–0136, Guangzhou, China).
The hens were randomly divided into 4 groups (each group has 6 replicates with 8 hens in one replicate) and subjected to the following feeding strategies: (1) control group, basal diet; (2) PSM-L group, basal diet + 3 g/kg PSM; (3) PSM-M group, basal diet + 6 g/kg PSM and (4) PSM-H group, basal diet + 10 g/kg PSM. The photoperiod was set at 16 L:8 D throughout the study. The chickens were housed in three-tiered cages (45 × 40 × 40 cm, two chickens per cage), and they received 85 g of feed per bird per day to prevent overfeeding and had access to fresh water ad libitum. The breeder hens received artificial insemination every 3 days with 35 μL of pooled semen per bird, as described by Liu [
29]. The ingredients and nutrient composition of the basic diets are shown in
Table 1.
2.3. Productive Performance and Egg Quality
Regarding the productive performance, the daily egg production, egg weight, number of abnormal eggs (broken, small, large or shell-less eggs) and number of dead chickens were recorded for each replicate during the experimental period. Egg production records were used to calculate the average egg weight, laying rate, abnormal egg rate and mortality rate.
To evaluate egg quality, 24 qualified eggs from each group (4 eggs/replication × 6 replicates/group) were collected after 56 days of the feeding experiment and used for egg quality analysis. The weight of the egg, yolk, white and eggshell was measured using an electronic scale. The long/short diameter and eggshell thickness of the eggs were measured using a vernier caliper. The eggshell strength and protein height were determined using an eggshell strength tester (RH-DQ200, Runhu International Co., Ltd., Guangzhou, China). The egg yolk color was determined using an Egg Analyzer (EA-01, Orka Technology Ltd., Ramat Hasharon, Israel).
To evaluate the hatching performance, during the 8th week of the experiment, eggs were collected on 4 consecutive days, and 3 eggs were selected per day from each replicate. The eggs were stored in the same incubator (Bengbu Sanyuan Incubation Equipment Co., Ltd., Anhui, China) and incubated at 37.2 °C to 38.0 °C and 60 to 75% relative humidity. Unfertilized hatching eggs and dead embryonic eggs were identified using egg illumination on the 13th day of incubation. At the end of incubation on the day 21, the number of dead eggs was counted, and the number of chicks was recorded. The fertilization rate, dead embryo rate and hatching rate of the hatched eggs were counted in each replicate.
2.4. Measurements of the Fatty Acid Content of Egg Yolks
At the end of the experiment, 12 eggs per group were collected (2 eggs/replication × 6 replicates/group) to measure the fatty acid content of the yolks using a Shimadzu GC-2010 PRO gas chromatograph. The mean content of each fatty acid was used to calculate the total saturated fatty acids (SFA), total monounsaturated fatty acids (MUFA) and total PUFA. The fatty acid measurement services were provided by Waltek Testing Group (Foshan, China) Co., Ltd.
2.5. Collection of Blood and Tissue Samples
At the end of the feeding period, six hens (one hen per replicate) were randomly selected and fasted for 12 h for sampling. The blood samples were collected from the wing vein into tubes, and the tubes were left at room temperature for 20 min and then centrifuged at 4 °C for 10 min at 3000× g to collected the serum. The serum samples were stored at −20 °C for subsequent analysis. The chickens were euthanized by exsanguination, and the liver and middle jejunum were immediately removed and quickly frozen at −80 °C for further analysis.
2.6. Biochemical Variables and Antioxidant Capacity of Plasma and Liver
The serum glucose (GLU), alanine aminotransferase (ALT), and aspartate aminotransferase (AST) levels were determined using an automatic analyzer (COBUS MIRA Plus, Roche Diagnostic System Inc., Rotkreuz, Switzerland) according to the manufacturer’s guidelines. The plasma concentrations of total cholesterol (TC), triglyceride (TG), low-density lipoproteins (LDL), malondialdehyde (MDA), superoxide dismutase (SOD), total antioxidant capacity (T-AOC) and glutathione (GSH) were analyzed using colorimetric kits (Jiancheng Bioengineering Institute, Nanjing, China). The plasma concentrations of follicle-stimulating hormone (FSH) and estradiol (E2) were analyzed using ELISA kits (Shanghai Yuangju Biotechnology Center, Shanghai, China). The antioxidant capacity of the liver was determined using SOD and MDA assay kits (Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions. The liver TG and TC were assayed using assay kits (Jiancheng Bioengineering Institute, Nanjing, China). For sample preparation, a small amount (approximately 1 g) of the tissue samples was accurately weighed, and pre-cooled saline solution was added in a ratio of weight (g) to volume (mL) = 1:9, homogenized in an ice-water bath and centrifuged at 3600× g rpm for 10 min at 4 °C. The supernatant was collected and used for testing.
2.7. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
The total RNA was extracted from the liver samples using TRIzol reagent (Vazyme Biotech Co., Ltd., Nanjing, China). The total cDNA was synthesized with the total RNA (1 μg) using HiScript III RT SuperMix for qPCR (+gDNA Wiper) (Vazyme) and quantitative real-time PCR (RT-qPCR) amplification using the ChamQ universal SYBR qPCR Master Mix (Vazyme) and a QuantStudio
®5 (Thermo Fisher Scientific, Inc., Waltham, MA, USA). The primer sequences used for PCR are listed in
Table 2, and the relative expression of the target gene was analyzed using the 2
−ΔΔCt method after normalization against the geometric mean of expression of β-actin and GAPDH.
2.8. Histopathological Evaluation of Liver Tissues
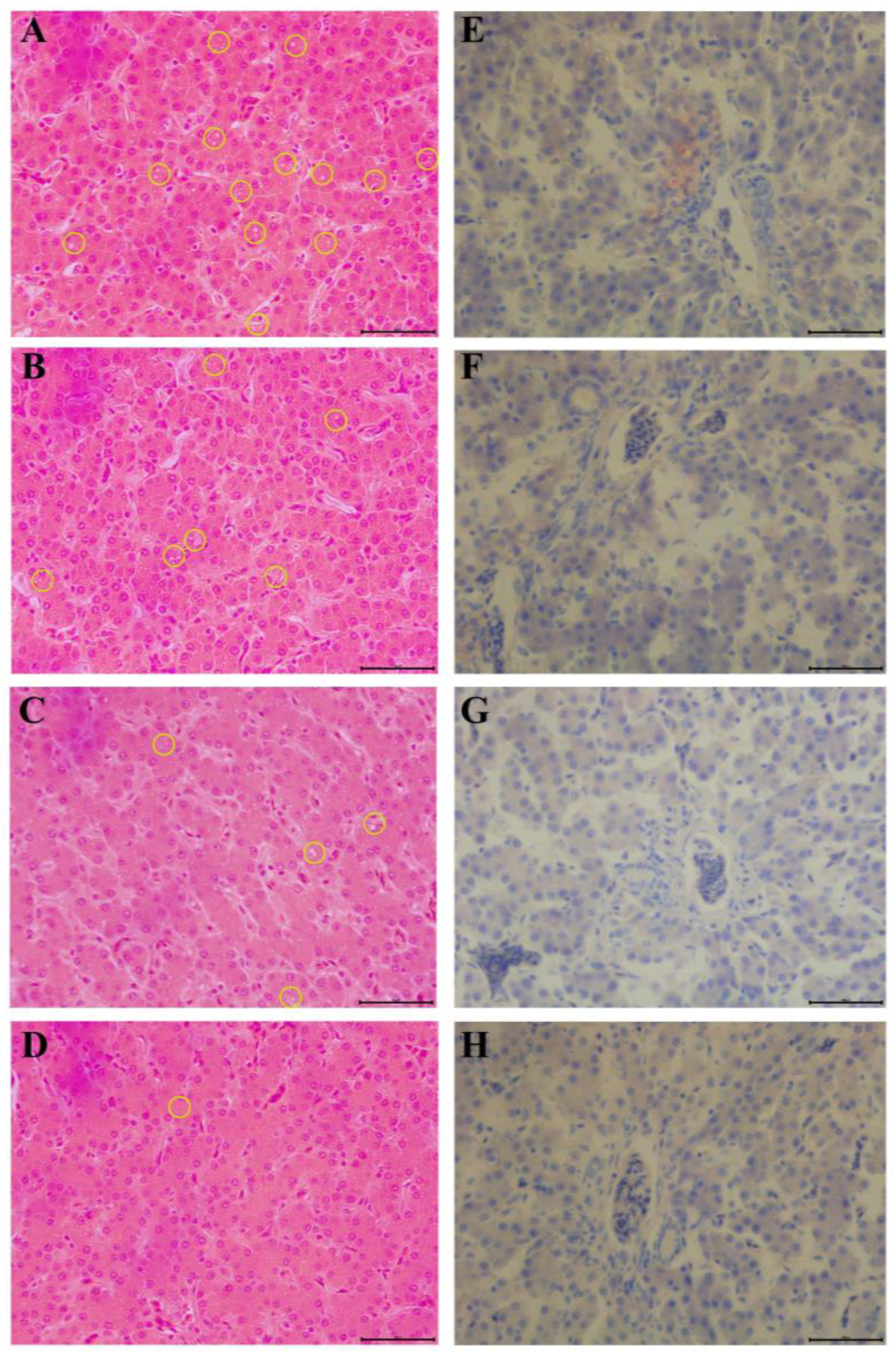
The liver tissues were fixed in 4% paraformaldehyde for 24 h and embedded in paraffin, followed by preparation of hematoxylin–eosin (H&E staining) and Oil Red O sections. The histological samples were evaluated based on tissue structural integrity and oil amount (Service-Bio, Wuhan, China).
2.9. Statistical Analysis
All thedata were first organized using Excel software and then statistically analyzed using SPSS 20 and GraphPad Prism 7.0 software. A one-way ANOVA was used to analyze the differences among groups for multiple group data comparison, and the least significant difference (LSD) method was used for multiple comparisons with Duncan’s new repolarization difference test. p < 0.05 indicated that differences were statistically significant.
4. Discussion
It was found that the addition of 300 mg/kg or 250 mg/kg of PSE to the diet significantly increased the rate of egg production, egg qualification, fertilization and hatching, while reducing the mortality of chickens [
30,
31]. In contrast, Sun [
32] found that replacing soybean meal with 10 to 20% PSM in the ration had no significant impact on egg production rate. In this experiment, we used PSM directly instead of PSM extracts. The addition of PSM to the diet had no significant effect on egg production rates. However, the fertilization rate and hatchability rate improved when the diet was supplemented with 1% PSM. In addition, the 0.6% PSM diet significantly reduced the mortality of the hens. Further research is needed to determine whether directly increasing the PSM dose can improve egg production, or whether an active ingredient in PSE increases the production of laying hens. The results showed that adding 0.6% PSM to the diet significantly increased the eggs’ protein contents and Haugh units. These results are inconsistent with those of Sun [
32]. They observed no significant effect of the 10 to 20% percent PSM diet combination on the protein contents or Haugh units of the eggs. These differences may be related to the different types of PS.
Previous studies have shown that PS supplementation in the diet of laying hens significantly increases the levels of PUFAs in their eggs [
26,
27]. Consistent with previous research, dietary PSM had a significant impact on the fatty acid composition of the egg yolks. MUFAs and PUFAs were increased in all the PSM groups. PSM still contains a certain amount of oil after extraction. These oils should enter the egg via the hen and thereby influence the fatty acid composition of the yolk [
33].
In rats fed a low-fat diet (7% by weight), the weight of their epididymal adipose tissue was slightly but not significantly lower than a perilla pomace-fed group [
34]. The abdominal fat index of laying hens was reduced in the 0.6% PSM group in this experiment. According to Anene [
35], poor feed efficiency in chickens leads to increased abdominal fat pad weight and liver weight, eventually leading to fatty liver hemorrhagic syndrome. PSM containing rosmarinic acid can improve nutrient digestibility [
36]. PSM could reduce the abdominal fat index by improving feed digestibility in hens.
The hormone FSH can promote the growth of immature follicles in the ovary and increase the rate of egg laying [
37]. Previous studies have shown that E2 levels are closely related to laying performance, with E2 concentrations being highest during peak laying periods [
38]. In this experiment, E2 was significantly increased in the 1% PSM group. The flavonoids found in PSE have weak estrogenic activity and can bind to estrogen receptors in vivo, resulting in bidirectional regulation of animal performance [
39]. It is likely that for this reason, the number of preovulatory follicles and serum E2 levels of the hens could be increased by the flavonoids contained in the 1% PSM diet.
The SOD and GSH-Px are important antioxidant enzymes. The activity of T-AOC demonstrates its ability to eliminate oxygen radicals in the body. Another indicator was MDA, one of the end products of lipid peroxidation and a marker of oxidative stress [
40]. Oxidative stress is caused by the excessive accumulation of reactive oxygen species (ROS). Excessive ROS accumulation leads to damage to macromolecules such as proteins, lipids and nucleic acids in the body, ultimately causing disease [
41]. Rosmarinic acid plays an antioxidant role by reducing the production of reactive oxygen species, regulating lipid peroxides and improving the activity of antioxidant enzymes [
42]. The polysaccharides in PSM exhibit antioxidant potential by scavenging free radicals [
43]. This reduces H
2O
2-induced intracellular ROS generation in cells by increasing GSH, SOD and CAT levels and decreasing MDA levels [
43]. Rosmarinic acid in PSM activates the antioxidant defense mechanism system via the
Nrf2-mediated pathway and alleviates heat stress in laying hens [
36,
44]. Activation of the
Nrf2/
HO-1 signaling pathway can increase the expression of various antioxidant genes and reduce oxidative stress in tissues; for example, by increasing the expression of
CAT,
SOD,
HO-1 and
GSH synthesis [
45,
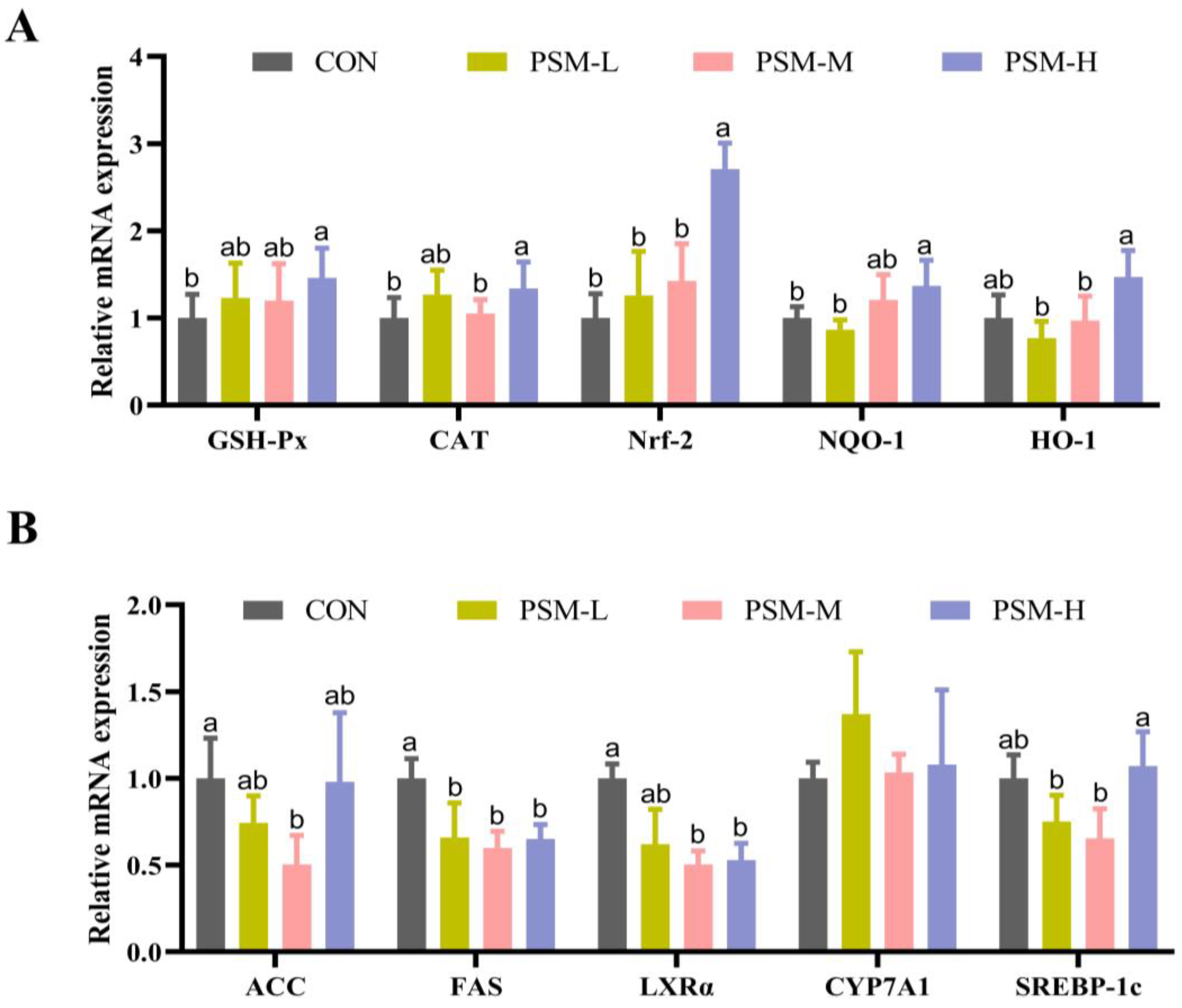
46]. Similar to the above results, the PSM diet upregulated the gene expression of factors related to liver antioxidant and lipid metabolism, as well as serum SOD and T-AOC levels. These studies suggest that PSM may increase antioxidant capacity through multiple pathways. However, no significant reduction in MDA was observed, and further animal and cell studies are needed to explore this reason.
Food sources rich in PUFAs typically contain antioxidants and may also regulate glucose or cholesterol metabolism [
47]. The PUFAs in PSM are thought to affect the lipid metabolism of chickens, which in turn alters the corresponding glucose and lipid indicators. Glucose content was an important indicator of insulin resistance and hyperglycemia, which was reduced by the PSM diet in the chickens in these experiments [
48]. According to Cha et al., perilla oil (0.1 or 0.3%) not only reduced the development of hypercholesterolemia and atherosclerosis in rabbits on a high-cholesterol diet (HCD), but also reduced fat accumulation and lipid peroxidation in their liver and kidney tissues [
49]. After absorption from the intestinal tract, TG is collected in the liver and enters the bloodstream as a component of LDL-C. To evaluate the health status of birds, blood lipid levels such as glucose, TG, TCH, HDL-C and LDL-C are usually analyzed. Dietary PSE decreased serum levels of TC, TG and LDL-C but increased HDL-C in mice on a high-fat diet [
23,
50]. We found that PSM reduced serum TC, TG and LDL-C in chickens. It also reduced TC and TG in the liver. These results are consistent with those of previous studies. Furthermore, TG is the main component of lipid droplets in hepatocytes [
51]. These findings are consistent with the results of H&E staining and Oil Red O staining of liver tissue samples.
LXRα is highly expressed in liver tissue as a member of the nuclear receptor superfamily. It activates its important downstream target gene,
SREBP1, which in turn regulates genes related to lipid synthesis [
52], thus contributing to the regulation of lipid metabolism [
52,
53]. The main function of
SREBP-1c is to regulate fatty acid and triglyceride synthesis. Overexpression of this protein leads to the accumulation of lipids in non-adipose tissues, ultimately leading to tissue damage. The genes of
ACC and
FAS are the target genes associated with
SREBP-1c. They are related to lipid synthesis and glucose metabolism [
54,
55]. They also function as two rate-limiting enzymes involved in fatty acid synthesis, and their activities may reflect the body’s ability to synthesize fat to some extent [
56]. The
CYP7A1 gene plays a crucial role in regulating cholesterol homeostasis and bile acid biosynthesis. Similarly, Zhang et al. found that perilla pomace significantly reduced hepatic
FAS activity in rats [
34]. Based on the expression of
ACC and
FAS genes, it is speculated that PSM leads to a decrease in lipid synthesis levels by downregulating
ACC and
FAS expression, leading to a decrease in serum TG. Meanwhile, PSM inhibits lipid synthesis and lowers glucose levels via the
LXRα/SREBP1 signaling pathway.


 ,
, 



{kind=link}
{kind=link}
{kind=link}