
Monitoring of the Endangered Cave Salamander Speleomantes sarrabusensis



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results

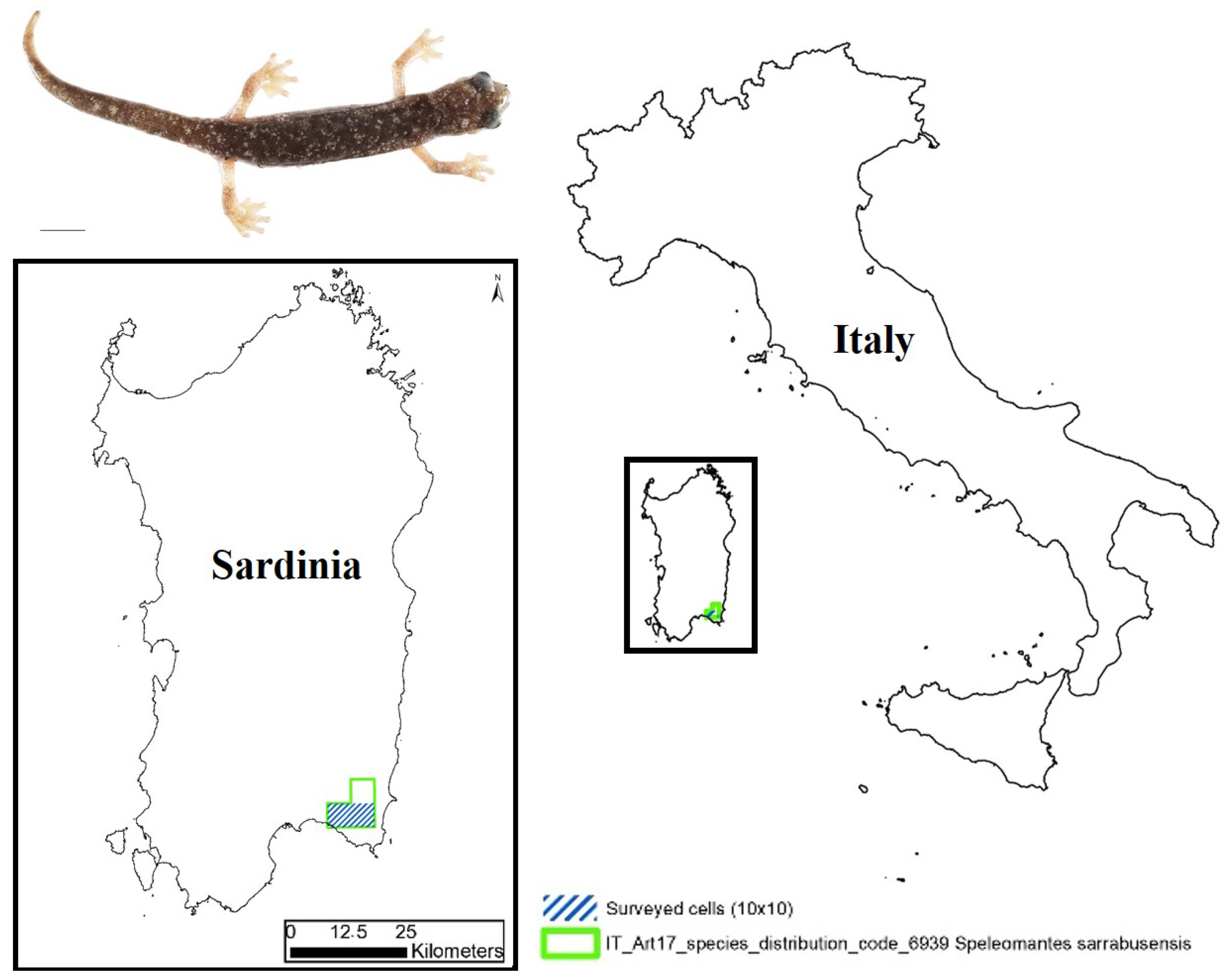{kind=link}
{kind=link}
| Number of Observed Individuals | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | Year | January | February | March | April | May | June | July | August | September | October | November | December |
| Spring 1 | 2015 | - | - | 4 | 40 | 72 | 79 | 66 | 50 | 83 | 16 | 1 | - |
| 2016 | 3 | - | 10 | 70 | 84 | 90 | 36 | 39 | 42 | 22 | 0 | 1 | |
| 2017 | 1 | 1 | 5 | 29 | 18 | 23 | 36 | 33 | 20 | 35 | 2 | 1 | |
| 2018 | 2 | 6 | 9 | 21 | 39 | 59 | - | - | - | - | - | - | |
| Spring 2 | 2015 | - | - | 0 | 6 | 10 | 2 | 5 | 5 | 7 | 18 | 3 | - |
| 2016 | 3 | - | 8 | 8 | 5 | 3 | 3 | 1 | 1 | 4 | 5 | 3 | |
| 2017 | 2 | 2 | 8 | 12 | 6 | 5 | 6 | 5 | 3 | 7 | 5 | 3 | |
| 2018 | 3 | 5 | 6 | 0 | 8 | 6 | - | - | - | - | - | - | |
| Data on the inner microclimate | |||||||||||||
| Site | January | February | March | April | May | June | July | August | September | October | November | December | |
| Spring 1 | Temperature | 10.2 (±0.8) | 10.3 (±0.1) | 10.3 (±0.3) | 12.2 (±1.3) | 13.2 (±1) | 14.2 | 16.4 (±0.8) | 17.3 (±0.8) | 16.5 | 15.2 (±0.3) | 13.0 (±0.7) | 9.8 |
| Humidity | 81.2 (±2.2) | 81.8 (±0.3) | 82.7 (±0.9) | 83.4 (±2.1) | 84.1 (±3) | 81.0 | 78.8 (±1.4) | 79.7 (±2.6) | 79.5 | 82.2 (±1) | 82.4 (±1.5) | 81.6 | |
| Spring 2 | Temperature | 10.0 (±1.3) | 9.9 (±0.3) | 10.9 (±0.1) | 11.9 (±0.8) | 12.6 (±0.7) | 13.9 | 15.4 (±0.6) | 16.0 (±0.9) | 15.7 | 14.2 (±0.1) | 12.8 (±1.3) | 9.3 |
| Humidity | 82.7 (±1) | 82.6 (±1.1) | 82.6 (±0.9) | 84.1 (±2.8) | 84.5 (±3.4) | 82.6 | 82.2 (±0.4) | 81.8 (±1) | 81.6 | 82.9 (±1.2) | 80.0 (±2.8) | 82.4 | |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lanza, B.; Pastorelli, C.; Laghi, P.; Cimmaruta, R. A review of systematics, taxonomy, genetics, biogeography and natural history of the genus Speleomantes Dubois, 1984 (Amphibia Caudata Plethodontidae). Atti. Mus. Civ. Stor. Nat. Trieste 2006, 52, 5–135. [Google Scholar]
- Ficetola, G.F.; Pennati, R.; Manenti, R. Do cave salamanders occur randomly in cavities? An analysis with Hydromantes strinatii. Amphib.-Reptil. 2012, 33, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Do cave features affect underground habitat exploitation by non-troglobite species? Acta. Oecol. 2014, 55, 29–35. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Lunghi, E.; Canedoli, C.; Padoa-Schioppa, E.; Pennati, R.; Manenti, R. Differences between microhabitat and broad-scale patterns of niche evolution in terrestrial salamanders. Sci. Rep. 2018, 8, 10575. [Google Scholar] [CrossRef] [Green Version]
- IUCN SSC Amphibian Specialist Group. Speleomantes Sarrabusensis. The IUCN Red List of Threatened Species 2022. 2022. Available online: https://www.iucnredlist.org/species/135825/89699335 (accessed on 3 December 2022).
- Lanza, B.; Leo, P. Prima osservazione sicura di riproduzione vivipara nel genere Speleomantes (Amphibia: Caudata: Plethodontidae). In Proceedings of the Atti 3° Congresso Nazionale Societas Herpetologica Italica (Pavia), Pianura, Cremona, 14–16 September 2000; pp. 317–319. [Google Scholar]
- Lunghi, E.; Corti, C.; Manenti, R.; Barzaghi, B.; Buschettu, S.; Canedoli, C.; Cogoni, R.; De Falco, G.; Fais, F.; Manca, A.; et al. Comparative reproductive biology of European cave salamanders (genus Hydromantes): Nesting selection and multiple annual breeding. Salamandra 2018, 54, 101–108. [Google Scholar]
- Lunghi, E.; Manenti, R.; Mulargia, M.; Veith, M.; Corti, C.; Ficetola, G.F. Environmental suitability models predict population density, performance and body condition for microendemic salamanders. Sci. Rep. 2018, 8, 7527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Seasonal variation in microhabitat of salamanders: Environmental variation or shift of habitat selection? PeerJ 2015, 3, e1122. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, M.K.; Deban, S.M. Cling performance and surface area of attachment in plethodontid salamanders. J. Exp. Biol. 2020, 233, jeb211706. [Google Scholar] [CrossRef]
- Costa, A.; Crovetto, F.; Salvidio, S. European plethodontid salamanders on the forest floor: Local abundance is related to fine-scale environmental factors. Herpetol. Conserv. Biol. 2016, 11, 344–349. [Google Scholar]
- Lunghi, E.; Romeo, D.; Mulargia, M.; Cogoni, R.; Manenti, R.; Corti, C.; Ficetola, G.F.; Veith, M. On the stability of the dorsal pattern of European cave salamanders (genus Hydromantes). Herpetozoa 2019, 32, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Manenti, R.; Mancinelli, G.; Corti, C.; Ficetola, G.F. What shapes the trophic niche of European plethodontid salamanders? PLoS ONE 2018, 13, e0205672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunghi, E.; Ficetola, G.F.; Mulargia, M.; Cogoni, R.; Veith, M.; Corti, C.; Manenti, R. Batracobdella leeches, environmental features and Hydromantes salamanders. Int. J. Parasitol. Parasites Wildl. 2018, 7, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Ficetola, G.F.; Romano, A.; Salvidio, S.; Sindaco, R. Optimizing monitoring schemes to detect trends in abundance over broad scales. Anim. Conserv. 2017, 21, 221–231. [Google Scholar] [CrossRef]
- Lunghi, E.; Bruni, G. Long-term reliability of Visual Implant Elastomers in the Italian cave salamander (Hydromantes italicus). Salamandra 2018, 54, 283–286. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; Team, R.C. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 2016, 3, 1–128. [Google Scholar]
- Lunghi, E.; Corti, C.; Manenti, R.; Ficetola, G.F. Consider species specialism when publishing datasets. Nat. Ecol. Evol. 2019, 3, 319. [Google Scholar] [CrossRef]
- Lunghi, E.; Corti, C.; Mulargia, M.; Zhao, Y.; Manenti, R.; Ficetola, G.F.; Veith, M. Cave morphology, microclimate and abundance of five cave predators from the Monte Albo (Sardinia, Italy). Biodivers Data. J. 2020, 8, e48623. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.L.; Hines, J.E. Occupancy Estimation and Modeling. Inferring Patterns and Dynamics of Species Occurrence; Academic Press: San Diego, CA, USA, 2006. [Google Scholar]
- Cogoni, R.; Mulargia, M.; Manca, S.; Croubu, V.; Giachello, S.; Lunghi, E. New observations on the tree-dwelling behaviour of European cave salamanders (genus Speleomantes). In Proceedings of the XIV Congresso Nazionale della Societas Herpetologica Italica, Torino, Italia, 13–17 September 2022. [Google Scholar]
- Ficetola, G.F.; Barzaghi, B.; Melotto, A.; Muraro, M.; Lunghi, E.; Canedoli, C.; Lo Parrino, E.; Nanni, V.; Silva-Rocha, I.; Urso, A.; et al. N-mixture models reliably estimate the abundance of small vertebrates. Sci. Rep. 2018, 8, 10357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunghi, E.; Bacci, F.; Zhao, Y. How can we record reliable information on animal colouration in the wild? Diversity 2021, 13, 356. [Google Scholar] [CrossRef]
- Lunghi, E.; Giachello, S.; Manenti, R.; Zhao, Y.; Corti, C.; Ficetola, G.F.; Bradley, J.G. The post hoc measurement as a safe and reliable method to age and size plethodontid salamanders. Ecol. Evol. 2020, 10, 11111–11116. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Salvidio, S.; Posillico, M.; Altea, T.; Matteucci, G.; Romano, A. What goes in does not come out: Different non-lethal dietary methods give contradictory interpretation of prey selectivity in amphibians. Amphib.-Reptil. 2014, 35, 255–262. [Google Scholar] [CrossRef]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Mulargia, M.; Cogoni, R.; Barzaghi, B.; Cornago, L.; Avitabile, D.; Veith, M.; Manenti, R.; et al. Field-recorded data on the diet of six species of European Hydromantes cave salamanders. Sci. Data. 2018, 5, 180083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunghi, E.; Giachello, S.; Zhao, Y.; Corti, C.; Ficetola, G.F.; Manenti, R. Photographic database of the European cave salamanders, genus Hydromantes. Sci. Data. 2020, 7, 171. [Google Scholar] [CrossRef]
- Lunghi, E.; Manenti, R.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Corti, C.; Ficetola, G.F.; Mancinelli, G. Interspecific and inter-population variation in individual diet specialization: Do environmental factors have a role? Ecology 2020, 101, e03088. [Google Scholar] [CrossRef]
- Araújo, M.S.; Bolnick, D.L.; Layman, C.A. The ecological causes of individual specialisation. Ecol. Lett. 2011, 14, 948–958. [Google Scholar] [CrossRef]
- Roughgarden, J. Evolution of niche width. Am. Nat. 1972, 106, 683–718. [Google Scholar] [CrossRef]
- Bellinger, P.F.; Christiansen, K.A.; Janssens, F. Checklist of the Collembola of the World. Available online: https://www.collembola.org/ (accessed on 3 December 2022).
- Araújo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species–climate impact models under climate change. Glob. Chang. Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Halliday, T.R. Declining amphibians in Europe, with particular emphasis on the situation in Britain. Environ. Rev. 1993, 1, 21–25. [Google Scholar] [CrossRef]
- Taylor, S.J.; Krejca, J.K.; Niemiller, M.L.; Dreslik, M.J.; Phillips, C.A. Life history and demographic differences between cave and surface populations of the Western slimy salamander, Plethodon albagula (Caudata: Plethodontidae), in central Texas Herpetol. Conserv. Biol. 2015, 10, 740–752. [Google Scholar]


| Population | Latitude | Longitude | Elevation (m a.s.l.) | Surveyed Area (m2) | Microclimate | Survey #1 | Survey #2 | Survey #3 | Estimated Abundance | 2.5% | 97.5% |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Springs | 39°29′ | 9°44′ | 783 | 70 | 15.4 °C—79.2% | 21 | 14 | 3 | 34 | 28 | 41 |
| Forest #1 | 39°12′ | 9°28′ | 444 | 2454 | 10.3 °C—80% | 10 | 12 | 28 | 41 | 35 | 48 |
| Forest #2 | 39°14′ | 9°28′ | 569 | 2628 | 10.3 °C—80.4% | 2 | 0 | 1 | 9 | 5 | 16 |
| Scrub #1 | 39°15′ | 9°23′ | 620 | 320 | 8.7 °C—79.9% | 0 | 2 | 0 | 9 | 4 | 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cogoni, R.; Di Gregorio, M.; Cianferoni, F.; Lunghi, E. Monitoring of the Endangered Cave Salamander Speleomantes sarrabusensis. Animals 2023, 13, 391. https://doi.org/10.3390/ani13030391
Cogoni R, Di Gregorio M, Cianferoni F, Lunghi E. Monitoring of the Endangered Cave Salamander Speleomantes sarrabusensis. Animals. 2023; 13(3):391. https://doi.org/10.3390/ani13030391
Chicago/Turabian StyleCogoni, Roberto, Milos Di Gregorio, Fabio Cianferoni, and Enrico Lunghi. 2023. "Monitoring of the Endangered Cave Salamander Speleomantes sarrabusensis" Animals 13, no. 3: 391. https://doi.org/10.3390/ani13030391
APA StyleCogoni, R., Di Gregorio, M., Cianferoni, F., & Lunghi, E. (2023). Monitoring of the Endangered Cave Salamander Speleomantes sarrabusensis. Animals, 13(3), 391. https://doi.org/10.3390/ani13030391