
The Contribution of Phytate-Degrading Enzymes to Chicken-Meat Production


Abstract
:Simple Summary
Abstract
1. Introduction
2. Background
2.1. Phytate
2.2. Phytase
3. Extra-Phosphoric Effects of Exogenous Phytase
3.1. Mechanisms Underlying the Protein Effect of Phytate and Phytase
3.1.1. Mucin
3.1.2. Sodium
3.1.3. Amino Acid Absorption
3.1.4. Monomeric Amino Acid Co-Absorption with Sodium Driven by Na⁺/K⁺-ATPase
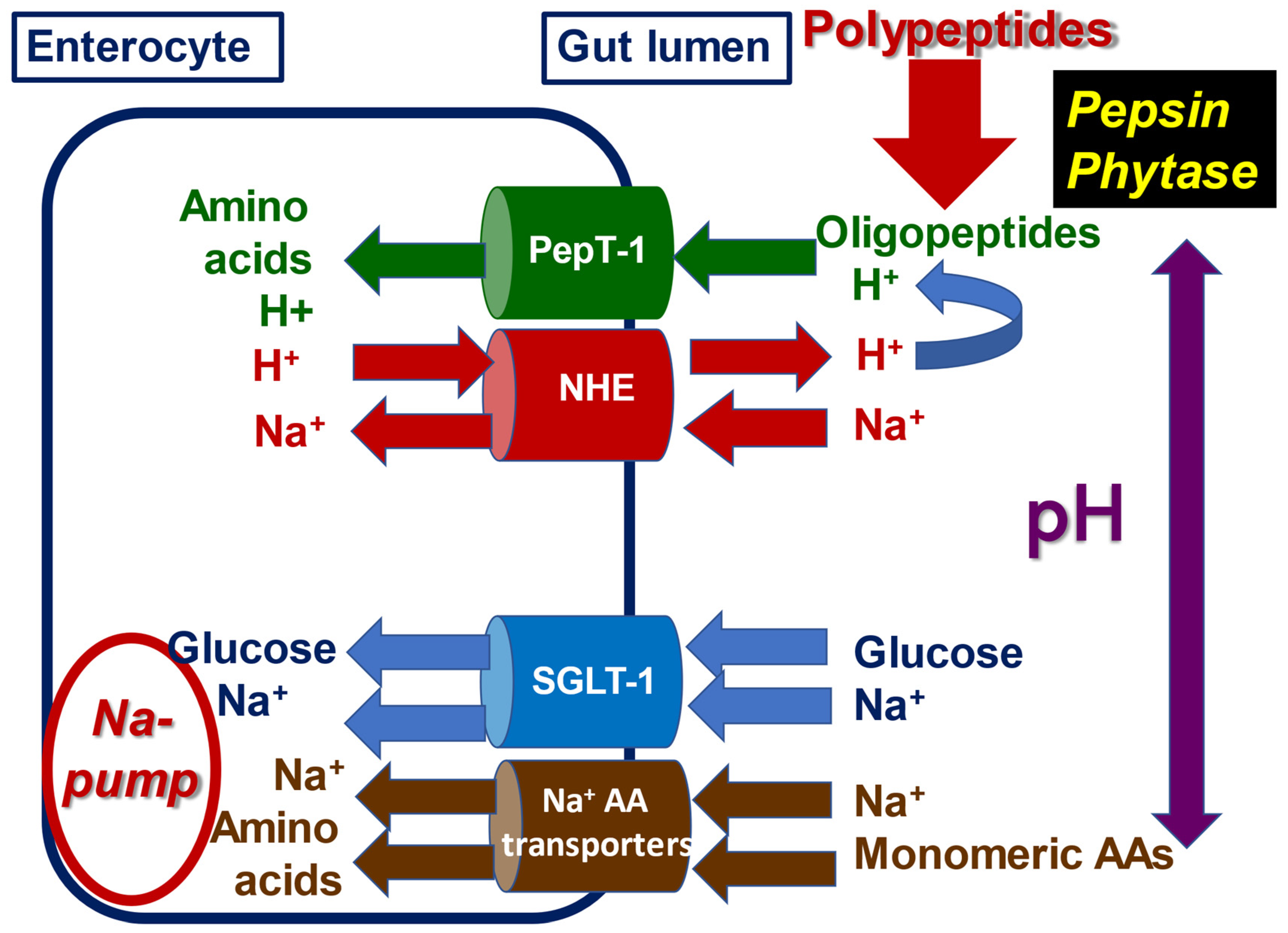
3.1.5. Di- and Tripeptide (Oligopeptide) Absorption via PepT-1 and NHE
3.1.6. Digesta pH along the Small Intestine
3.1.7. Rationale for a Novel Postulate
3.2. Energy Effect of Phytase
4. Phytase Matrix Values
5. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simons, P.C.M.; Versteegh, H.A.J.; Jongbloed, A.W.; Kemme, P.A.; Slump, P.; Bos, K.D.; Wolters, M.G.E.; Beudeker, R.F.; Verschoor, G.J. Improvement of phosphorus availability by microbial phytase in broilers and pigs. Br. J. Nutr. 1990, 64, 525–540. [Google Scholar] [CrossRef]
- Ravindran, V.; Bryden, W.L.; Kornegay, E.T. Phytates: Occurrence, bioavailability and implications in poultry nutrition. Poult. Avian Biol. Rev. 1995, 6, 125–143. [Google Scholar]
- Angel, R.; Tamim, N.M.; Applegate, T.J.; Dhandu, S.; Ellestad, L.E. Phytic acid chemistry: Influence on phytin-phosphorus availability and phytase efficacy. J. Appl. Poult. Res. 2002, 11, 471–480. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Acamovic, T.; Bedford, M.R. Phytic acid and phytase: Implications for protein utilization by poultry. Poult. Sci. 2006, 85, 878–885. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 2007, 135, 1–41. [Google Scholar] [CrossRef]
- Selle, P.H.; Cowieson, A.J.; Cowieson, N.P.; Ravindran, V. Protein-phytate interactions in pig and poultry nutrition: A reappraisal. Nutr. Res. Rev. 2012, 25, 1–17. [Google Scholar] [CrossRef]
- Lei, X.G.; Weaver, J.D.; Mullaney, E.; Ullah, A.H.; Azain, M.J. Phytase, a new life for an “old” enzyme. Ann. Rev. Anim. Biosci. 2013, 1, 283–309. [Google Scholar] [CrossRef]
- Woyengo, T.A.; Nyachoti, C.M. Review: Anti-nutritional effects of phytic acid in diets for pigs and poultry—Current knowledge and directions for future research. Can. J. Anim. Sci. 2013, 93, 9–21. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Awati, A.; Schulze, H.; Partridge, G. Phytase in non-ruminant animal nutrition: A critical review on phytase activities in the gastrointestinal tract and influencing factors. J. Sci. Food Agric. 2014, 95, 878–896. [Google Scholar] [CrossRef]
- Moss, A.F.; Liu, S.Y.; Selle, P.H. Progress in comprehending the phytate-phytase axis in chicken-meat production. Anim. Prod. Sci. 2018, 58, 1767–1778. [Google Scholar] [CrossRef]
- Hartig, T. Uber das Klebermehl. Bot. Z. 1855, 13, 881–885. [Google Scholar]
- Lott, J.N.A.; Ockenden, I.; Raboy, V.; Batten, G.D. Phytic acid and phosphorus in crop seeds and fruit: A global estimate. Seed Sci. Res. 2000, 10, 11–33. [Google Scholar] [CrossRef]
- Correll, D.L. The role of phosphorus in the eutrophication of receiving waters: A review. J. Environ. Qual. 1998, 27, 261–266. [Google Scholar] [CrossRef]
- Sharpley, A. Agricultural phosphorus, water quality and poultry production: Are they compatible? Poult. Sci. 1999, 78, 660–673. [Google Scholar] [CrossRef]
- Abelson, P.H. A potential phosphorus crisis. Science 1999, 283, 2015. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.-O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Cordell, D.; White, S. Peak phosphorus: Clarifying the key issues of a vigorous debate about long-term phosphorus security. Sustainability 2011, 3, 2027–2049. [Google Scholar] [CrossRef]
- Prud’homme, M. Peak Phosphorous: An Issue to Be Addressed. In Fertilizers & Agriculture; International Fertilizer Industry Association (IFA): Paris, France, 2010; Volume 1. [Google Scholar]
- De Sousa, J.P.L.; Albino, L.F.T.; Vaz, R.G.M.; Rodrigues, K.F.; Da Silva, G.F.; Renno, L.N.; Barros, V.R.S.; Kaneko, I.N. The effect of dietary phytase on broiler performance and digestive, bone, and blood biochemistry characteristics. Braz. J. Poult. Sci. 2015, 17, 69–76. [Google Scholar] [CrossRef]
- Alltech Agri-Food Outlook. 2022. Available online: https://www.alltech.com/agri-food-outlook (accessed on 14 November 2022).
- Suzuki, U.; Yoshimura, K.; Takaishi, M. Uber ein Enzym “Phytase” das Anhydro-oxy-methylendiphosphosaure spaltet. Coll. Agric. Bull. Tokyo Imp. Uni. 1907, 7, 503–505. [Google Scholar]
- Selle, P.H.; Walker, A.R.; Bryden, W.L. Total and phytate-phosphorus contents and phytase activity of Australian-sourced feed ingredients for pigs and poultry. Aust. J. Exp. Agric. 2003, 45, 475–479. [Google Scholar] [CrossRef]
- Nelson, T.S.; Kirby, L.K. The calcium binding properties of natural phytate in chick diets. Nutr. Rep. Int. 1987, 35, 949–956. [Google Scholar]
- Nelson, T.S. Available calcium for poultry. In Proceedings of the Florida Nutrition Conference for Feed Manufacturers, Orlando, FL, USA, 1984; pp. 1–7. [Google Scholar]
- Schöner, F.-J.; Hoppe, P.P.; Schwartz, G. Comparative effects of microbial phytase and inorganic phosphorus on performance and retentions of phosphorus, calcium and crude ash in broilers. J. Anim. Physiol. Anim. Nutr. 1991, 66, 248–255. [Google Scholar] [CrossRef]
- Schöner, F.-J.; Hoppe, P.P.; Schwartz, G.; Weische, H. Comparison of microbial phytase and inorganic phosphate in male chickens: The influence on performance data, mineral retention and dietary calcium. J. Anim. Physiol. Anim. Nutr. 1993, 69, 235–244. [Google Scholar]
- Ptak, A.; Damian, J.; Bartosz, K.; Mateusz, R.; Krzysztof, Z.; Sylwester, S. Effect of different phytases on the performance, nutrient retention and tibia composition in broiler chickens. Arch. Anim. Breed. 2013, 56, 104–112. [Google Scholar] [CrossRef]
- Menezes-Blackburn, D.; Gabler, S.; Greiner, G. Performance of seven commercial phytases in an in vitro simulation of poultry digestive tract. J. Agric. Food Chem. 2015, 63, 6142–6149. [Google Scholar] [CrossRef] [PubMed]
- Igbasan, F.A.; Manner, K.; Miksch, G.; Borriss, R.; Farouk, A.; Simon, O. Comparative studies on the in vitro properties of phytases from various microbial origins. Arch. Für Tierernaehrung 2000, 53, 353–373. [Google Scholar] [CrossRef] [PubMed]
- Onyango, E.M.; Bedford, M.R.; Adeola, O. Phytase activity along the digestive tract of the broiler chick: A comparative study of an Escherichia coli-derived and Peniophora lycii phytase. Can. J. Anim. Sci. 2005, 85, 61–68. [Google Scholar] [CrossRef]
- Takemasa, M.; Murakami, H.; Yamazaki, M. Reduction of phosphorus excretion in chicks by addition of yeast phytase. J. Poult. Sci. 1996, 33, 104–111. [Google Scholar] [CrossRef]
- Truong, H.H.; Yu, S.; Moss, A.F.; Liu, S.Y.; Selle, P.H. Phytate degradation in the gizzard is pivotal to phytase responses in broiler chickens. In Proceedings of the Annual Australian Poultry Science Symposium, Sydney, Australia, 14–17 February 2016; Volume 27, pp. 174–177. [Google Scholar]
- Luttrell, B.M. The biological relevance of the binding of calcium ions by inositol phosphates. J. Biol. Chem. 1993, 268, 1521–1524. [Google Scholar] [CrossRef]
- Walk, C.L.; Santos, T.T.; Bedford, M.R. Influence of superdoses of a novel microbial phytase on growth performance, tibia ash, and gizzard phytate and inositol in young broilers. Poult. Sci. 2014, 93, 1172–1177. [Google Scholar] [CrossRef]
- Lee, S.A.; Bedford, M.R. Inositol—An effective growth promoter? World’s Poult. Sci. J. 2016, 72, 743–760. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Ptak, A.; Maćkowiak, P.; Sassek, M.; Pruszyńska-Oszmałek, E.; Żyła, K.; Świątkiewicz, S.; Kaczmarek, S.; Jozefiak, D. The effect of microbial phytase and myo-inositol on performance and blood biochemistry of broiler chickens fed wheat/corn-based diets. Poult. Sci. 2013, 92, 2124–2134. [Google Scholar] [CrossRef] [PubMed]
- Marchal, L.; Bello, A.; Sobotik, E.B.; Archer, G.; Dersjant-Li, Y. A novel consensus bacterial 6-phytase variant completely replaced inorganic phosphate in broiler diets, maintaining growth performance and bone quality: Data from two independent trials. Poult. Sci. 2021, 100, 100962. [Google Scholar] [CrossRef] [PubMed]
- Bello, A.; Dersjant-Li, Y.; van Eerden, E.; Kwakernaak, C.; Marchal, L. Supplementation of an all-plant-based inorganic phosphate-free diet with a novel phytase maintained tibia ash and performance in broilers under a commercial production setting. J. Appl. Poult. Res. 2021, 31, 100253. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Bello, A.; Esteve-Garcia, E.; Ramirez Crues, C.; Marchal, L. A novel consensus bacterial 6-phytase variant added to phytate-rich diets totally replaced inorganic phosphate in broilers. J. App. Anim. Nutr. 2022, 10, 59–70. [Google Scholar] [CrossRef]
- Ravindran, V. Phytases in poultry nutrition—An overview. In Proceedings of the Annual Australian Poultry Science Symposium; 1995; Volume 7, pp. 135–139. [Google Scholar]
- Hill, R.; Tyler, C. The reaction between protein and phytate. J. Agric. Sci. 1954, 44, 324–326. [Google Scholar] [CrossRef]
- Adeola, O.; Sands, J.S. Does supplementary microbial phytase improve amino acid utilization? A perspective that it does not. J. Anim. Sci. 2003, 81 (Suppl. 2), E78–E85. [Google Scholar]
- Ravindran, V.; Cabahug, S.; Ravindran, G.; Selle, P.H.; Bryden, W.L. Response of broiler chickens to microbial phytase supplementation as influenced by dietary phytic acid and non-phytate phosphorus levels. II. Effects on apparent metabolisable energy, nutrient digestibility and nutrient retention. Brit. Poult. Sci. 2000, 41, 193–200. [Google Scholar] [CrossRef]
- Ravindran, V.; Selle, P.H.; Ravindran, G.; Morel, P.C.H.; Kies, A.K.; Bryden, W.L. Microbial phytase improves performance, apparent metabolizable energy and ileal amino acid digestibility of broilers fed a lysine-deficient diet. Poult. Sci. 2001, 80, 338–344. [Google Scholar] [CrossRef]
- Amerah, A.M.; Plumstead, P.W.; Barnard, L.P.; Kumar, A. Effect of calcium level and phytase addition on ileal phytate degradation and amino acid digestibility of broilers fed corn-based diets. Poult. Sci. 2014, 93, 906–915. [Google Scholar] [CrossRef]
- Martínez-Vallespín, B.; Männer, K.; Ader, P.; Zentek, J. Evaluation of high doses of phytase in a low-phosphorus diet in comparison to a phytate-free diet on performance, apparent ileal digestibility of nutrients, bone mineralization, intestinal morphology, and immune traits in 21-day-old broiler chickens. Animals 2022, 12, 1955. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Bello, A.; Stormink, T.; Abdollahi, M.R.; Ravindran, V.; Babatunde, O.O.; Adeola, O.; Toghyani, M.; Liu, S.Y.; Selle, P.H.; et al. Modeling improvements in ileal digestible amino acids by a novel consensus bacterial 6-phytase variant in broilers. Poult. Sci. 2022, 101, 101666. [Google Scholar] [CrossRef] [PubMed]
- Cowieson, A.J.; Ruckebusch, J.-P.; Sorbara, J.O.B.; Wilson, J.W.; Guggenbuhl, P.; Roos, F.F. A systematic view on the effect of phytase on ileal amino acid digestibility in broilers. Anim. Feed Sci. Technol. 2017, 225, 182–194. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V.; Caldwell, R.A.; Bryden, W.L. Phytate and phytase: Consequences for protein utilisation. Nutr. Res. Rev. 2000, 13, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. The chemistry and biochemistry of inositol polyphosphates. Rev. Pure Appl. Chem. 1966, 16, 209–224. [Google Scholar]
- Rajendran, S.; Prakash, V. Kinetics and thermodynamics of the mechanism of interaction of sodium phytate with α-globulin. Biochemistry 1993, 32, 3474–3478. [Google Scholar] [CrossRef]
- Bye, J.W.; Cowieson, N.P.; Cowieson, A.J.; Selle, P.H.; Falconer, R.J. Dual effects of sodium phytate on the structural stability and solubility of proteins. J. Agric. Food Chem. 2013, 61, 290–295. [Google Scholar] [CrossRef]
- Darby, S.J.; Platts, L.; Daniel, M.S.; Cowieson, A.J.; Falconer, R.J. An isothermal titration calorimetry study of phytate binding to lysozyme. J. Therm. Anal. Calorim. 2017, 127, 1201–1208. [Google Scholar] [CrossRef]
- Wang, R.; Guo, S. Phytic acid and its interactions: Contributions to protein functionality, food processing, and safety. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2081–2105. [Google Scholar] [CrossRef]
- Janin, J.; Miller, S.; Chothia, C. Surface, subunit interfaces and interior of oligomeric proteins. J. Mol. Biol. 1988, 204, 155–164. [Google Scholar] [CrossRef]
- Anderson, P.A. Interactions between proteins and constituents that affect protein quality. In Digestibility and Amino Acid Availability in Cereals and Oilseeds; Finley, J.W., Hopkins, D.T., Eds.; American Association of Cereal Chemists: St Paul, MN, USA, 1985; pp. 31–45. [Google Scholar]
- Prattley, C.A.; Stanley, D.W.; van der Voort, F.R. Protein–phytate interactions in soybeans. II. Mechanism of protein–phytate binding as affected by calcium. J. Food Biochem. 1982, 16, 255–271. [Google Scholar] [CrossRef]
- Kies, A.K.; de Jonge, L.H.; Kemme, P.A.; Jongbloed, A.W. Interaction between protein, phytate, and microbial phytase. In vitro studies. J. Agric. Food Chem. 2006, 54, 1753–1758. [Google Scholar] [CrossRef] [PubMed]
- Csonka, F.A.; Murphy, J.C.; Jones, D.B. The iso-electric points of various proteins. J. Am. Chem. Soc. 1926, 48, 763–768. [Google Scholar] [CrossRef]
- Farner, D.S. The hydrogen ion concentration in avian digestive tracts. Poult. Sci. 1942, 21, 445–450. [Google Scholar] [CrossRef]
- Lee, S.A.; Dunne, J.; Frebery, E.; Wilcock, P.; Mottram, T.; Bedford, M.R. Superdosing phytase reduces real-time gastric pH in broilers and weaned piglets. Br. Poult. Sci. 2018, 59, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Shafey, T.M.; McDonald, M.W.; Dingle, J.G. Effects of dietary calcium and available phosphorus concentration on digesta pH and on the availability of calcium, iron, manganese and zinc from the intestinal contents of meat chickens. Br. Poult. Sci. 1991, 32, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, V.; Cabahug, S.; Ravindran, G.; Bryden, W.L. Influence of microbial phytase on apparent ileal amino acid digestibility in feedstuffs for broilers. Poult. Sci. 1999, 78, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Camus, M.C.; Laporte, J.C. Inhibition de la proteolyse pesique par le ble’. Role de l’acide phytique des issues (Inhibition of pepsin proteolysis by wheat. Role of phytic acid in the outcome). Annal. Biol. Anim. Biochim. Biophys. 1976, 16, 719–729. [Google Scholar] [CrossRef]
- Kanaya, K.; Yasumoto, K.; Mitsuda, H. Pepsin inhibition by phytate contained in rice bran. Rice Chem. Technol. 1976, 29, 341–346. [Google Scholar]
- Inagawa, J.; Kiyosawa, I.; Nagasawa, T. Effects of phytic acid on the digestion of casein and soybean protein with trypsin, pancreatin and pepsin. Nippon Eiyo Shokuryo Gakkaishi 1987, 40, 367–373. [Google Scholar] [CrossRef]
- Knuckles, B.E.; Kuzmicky, D.D.; Gumbmann, M.R.; Betschart, A.A. Effect of myo-inositol phosphate esters on in vitro and in vivo digestion of protein. J. Food Sci. 1989, 54, 1348–1350. [Google Scholar] [CrossRef]
- Vaintraub, I.A.; Bulmaga, V.P. Effect of phytate on the in vitro activity of digestive proteinases. J. Agric. Food Chem. 1991, 39, 859–861. [Google Scholar] [CrossRef]
- Yu, S.; Cowieson, A.; Gilbert, C.; Plumstead, P.; Dalsgaard, S. Interactions of phytate and myo-inositol phosphate esters (IP1–5) including IP5 isomers with dietary protein and iron and inhibition of pepsin. J. Anim. Sci. 2012, 90, 1824–1832. [Google Scholar] [CrossRef]
- Bohak, Z. Purification and characterization of chicken pepsinogen and chicken pepsin. J. Biol. Chem. 1969, 244, 4638–4648. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.L.; Marques, M.R.; Olivera, M.E.; Stippler, E.S. Enzymatic activity in the presence of surfactants commonly used in dissolution media, Part 1: Pepsin. Results Pharma Sci. 2016, 6, 15–19. [Google Scholar] [CrossRef]
- Woyengo, T.A.; Adeola, O.; Udenigwe, C.C.; Nyachoti, C.M. Gastro-intestinal digesta pH, pepsin activity and soluble mineral concentration responses to supplemental phytic acid and phytase in piglets. Livestock Sci. 2010, 134, 91–93. [Google Scholar] [CrossRef]
- Liu, N.; Ru, Y.J.; Li, F.D.; Wang, J.-P.; Lei, X.-G. Effect of dietary phytate and phytase on proteolytic digestion and growth regulation of broilers. Arch. Anim. Nutr. 2009, 63, 292–303. [Google Scholar] [CrossRef]
- Hersey, S.J. Pepsin secretion. In Physiology of the Gastrointestinal Tract; Johnson, L.R., Ed.; Raven Press: New York, NY, USA, 1987; Volume 2, pp. 947–957. [Google Scholar]
- Selle, P.H.; Ravindran, V.; Bryden, W.L.; Scott, T. Influence of dietary phytate and exogenous phytase on amino acid digestibility in poultry: A review. J. Poult. Sci. 2006, 43, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Van Burén, J.P.; Robinson, W.G. Formation of complexes between protein and tannic acid. J. Agric. Food Chem. 1969, 17, 772–777. [Google Scholar] [CrossRef]
- Mitjavila, S.; de Saint Blanquat, G.; Derache, R. Effect de l’acide tannique sur la secretion gastrique chez le rat. Ann. Nutr. Metab. 1973, 15, 163–170. [Google Scholar] [CrossRef]
- Mitjavila, S.; Lacombe, C.; Carrera, G.; Derache, R. Tannic acid and oxidized tannic acid on the functional state of rat intestinal epithelium. J. Nutr. 1997, 107, 2113–2121. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.; Flemström, G. Gastroduodenal mucus bicarbonate barrier: Protection against acid and pepsin. Am. J. Physiol.-Cell Physiol. 2005, 288, C1–C19. [Google Scholar] [CrossRef] [PubMed]
- Moss, A.F.; Chrystal, P.V.; McQuade, L.R.; Cadogan, D.J.; Liu, S.Y.; Selle, P.H. The influence of exogenous phytase on the post-enteral availability of amino acids in broiler chickens offered wheat-based diets. Anim. Feed Sci. Technol. 2019, 258, 114300. [Google Scholar] [CrossRef]
- Lien, K.A.; Sauer, W.C.; Fenton, M. Mucin output in ileal digesta of pigs fed a protein-free diet. Z. Ernährungswissenschaft 1997, 36, 182–190. [Google Scholar] [CrossRef]
- Fang, R.; Mantle, M.; Ceri, H. Characterization of quail intestinal mucin as a ligand for endogenous quail lectin. Biochem. J. 1993, 293, 867–872. [Google Scholar] [CrossRef]
- Onyango, E.M.; Asem, E.K.; Adeola, O. Phytic acid increases mucin and endogenous amino acid losses from the gastrointestinal tract of chickens. Br. Poult. Sci. 2009, 101, 836–842. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Ravindran, V.; Selle, P.H. Influence of dietary phytic acid and source of microbial phytase on ileal endogenous amino acid flows in broiler chickens. Poult. Sci. 2008, 87, 2287–2299. [Google Scholar] [CrossRef]
- Ravindran, V. Feed-induced specific ileal endogenous amino acid losses: Measurement and significance in the protein nutrition of monogastric animals. Anim. Feed Sci. Technol. 2016, 221, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Souffrant, W.B.; Rerat, A.; Laplace, J.B.; Darcy, B.V.; Kohler, R.; Corring, T.; Gebhardt, G. Exogenous and endogenous contributions to nitrogen fluxes in the digestive tract of pigs fed a casein diet. iii. refluxing of endogenous nitrogen. Reprod. Nutr. Dev. 1993, 33, 373–382. [Google Scholar] [CrossRef]
- Greenhalgh, S.; Lemme, A.; Dorigam, J.C.d.e.P.; Chrystal, P.V.; Macelline, S.P.; Liu, S.Y.; Selle, P.H. Dietary crude protein concentrations, feed grains and whey protein interactively influence apparent digestibility coefficients of amino acids, protein, starch and performance of broiler chickens. Poult. Sci. 2022, 101, 102131. [Google Scholar] [CrossRef]
- Krieg, J.; Borda-Molina, D.; Siegert, W.; Sommerfeld, V.; Chi, Y.P.; Taheri, H.R.; Feuerstein, D.; Camarinha-Silva, A.; Rodehutscord, M. Effects of calcium level and source, formic acid, and phytase on phytate degradation and the microbiota in the digestive tract of broiler chickens. Anim. Microbiome 2021, 3, 23. [Google Scholar] [CrossRef]
- Lien, K.A.; Sauer, W.C.; He, J.M. Dietary influences on the secretion into and degradation of mucin in the digestive tract of monogastric animals and humans. J. Anim. Feed Sci. 2001, 10, 223–245. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Acamovic, T.; Bedford, M.R. The effects of phytase and phytic acid on the loss of endogenous amino acids and minerals from broiler chickens. Br. Poult. Sci. 2004, 45, 101–108. [Google Scholar] [CrossRef]
- Ravindran, V.; Morel, P.C.H.; Partridge, G.G.; Hruby, M.; Sands, J.S. Influence of an E. coli-derived phytase on nutrient utilization in broiler starters fed diets containing varying concentrations of phytic acid. Poult. Sci. 2006, 85, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Selle, P.H.; Partridge, G.G.; Ravindran, V. Beneficial effects of xylanase and/or phytase inclusions on ileal amino acid digestibility energy utilisation mineral retention and growth performance in wheat-based broiler diets. Anim. Feed Sci. Technol. 2009, 153, 303–313. [Google Scholar] [CrossRef]
- Truong, H.H.; Yu, S.; Peron, A.; Cadogan, D.J.; Khoddami, A.; Roberts, T.H.; Liu, S.Y.; Selle, P.H. Phytase supplementation of maize-, sorghum- and wheat-based broiler diets with identified starch pasting properties influences phytate (IP6) and sodium jejunal and ileal digestibility. Anim. Feed Sci. Technol. 2014, 198, 248–256. [Google Scholar] [CrossRef]
- Truong, H.H.; Bold, R.M.; Liu, S.Y.; Selle, P.H. Standard phytase inclusion in maize-based broiler diets enhances digestibility coefficients of starch, amino acids and sodium in four small intestinal segments and digestive dynamics of starch and protein. Anim. Feed Sci. Technol. 2015, 209, 240–248. [Google Scholar] [CrossRef]
- Truong, H.H.; Yu, S.; Moss, A.F.; Partridge, G.G.; Liu, S.Y.; Selle, P.H. Phytase inclusions of 500 and 2000 FTU/kg in maize-based broiler diets impact on growth performance, nutrient utilisation, digestive dynamics of starch, protein (N), sodium and IP6 phytate degradation in the gizzard and four small intestinal segments. Anim. Feed Sci. Technol. 2017, 223, 13–22. [Google Scholar] [CrossRef]
- Van der Klis, J.D.; Verstegen, M.W.A.; De Wit, W. Absorption of minerals and retention time of dry matter in the gastrointestinal tract of broilers. Poult. Sci. 1990, 69, 2185–2194. [Google Scholar] [CrossRef]
- Case, R.M.; Scratcherd, T.; Wynne, R.D.A. The origin and secretion of pancreatic juice bicarbonate. J. Physiol. 1970, 210, 1–15. [Google Scholar] [CrossRef]
- Croom, W.J.; Brake, J.; Coles, B.A.; Havenstein, G.B.; Christensen, V.L.; McBride, B.W.; Peebles, E.D.; Taylor, I.L. Is intestinal absorption rate-limiting for performance in poultry? J. Appl. Poult. Res. 1999, 8, 242–252. [Google Scholar] [CrossRef]
- Kiela, P.R.; Ghishan, F.K. Physiology of intestinal absorption and secretion. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Wright, E.M. The intestinal Na+/glucose co-transporter. Ann. Rev. Physiol. 1993, 55, 575–589. [Google Scholar] [CrossRef]
- Hyde, R.; Taylor, P.M.; Hundal, H.S. Amino acid transporters: Roles in amino acid sensing and signaling in animal cells. Biochem. J. 2003, 373, 1–18. [Google Scholar] [CrossRef]
- Miska, K.B.; Fetterer, R.H.; Wong, E.A. mRNA expression of amino acid transporters, aminopeptidase, and the di- and tri-peptide transporter PepT1 in the intestine and liver of the post-hatch chick. Poult. Sci. 2015, 94, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Glynn, I.M. All hands to the sodium pump. J. Physiol. 1993, 462, 1–30. [Google Scholar] [CrossRef]
- Glynn, I.M. A hundred years of sodium pumping. Ann. Rev. Physiol. 2002, 64, 1–18. [Google Scholar] [CrossRef]
- Dilworth, L.L.; Omoruyi, F.O.; Asemota, H.N. Digestive and absorptive enzymes in rats fed phytic acid extract from sweet potato (Ipomoea batatas). Diabetol. Croat. 2005, 34, 59–65. [Google Scholar]
- Liu, N.; Ru, Y.; Li, F.; Cowieson, A.J. Effect of diet containing phytate and phytase on the activity and messenger ribonucleic acid expression of carbohydrase and transporter in chickens. J. Anim. Sci. 2008, 86, 3432–3439. [Google Scholar] [CrossRef] [Green Version]
- Therien, A.G.; Blostein, R. Mechanisms of sodium pump regulation. Am. J. Physiol. 2000, 279, C541–C566. [Google Scholar] [CrossRef]
- Clarke, R.J.; Kane, D.J. Two gears of pumping by the sodium pump. Biophys. J. 2007, 93, 4187–4196. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Amezcua, C.; Parsons, C.M.; Baker, D.H. Effect of microbial phytase on phosphorus bioavailability, apparent metabolizable energy, and amino acid digestibility in distillers dried grains with solubles in chicks. Poult. Sci. 2006, 85, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Bhartur, S.G.; Bookstein, C.; Much, M.W.; Boxer, R.; Chang, E.B.; Rao, M.C. An avian sodium-hydrogen exchanger. Comp. Biochem. Physiol. 1997, 118, 883–889. [Google Scholar] [CrossRef]
- Krehbiel, C.R.; Matthews, J.C. Absorption of amino acids and peptides. In Amino Acids in Animal Nutrition, 2nd ed.; D’Mello, J.P.F., Ed.; CABI: Wallingford, UK, 2003; pp. 41–70. [Google Scholar]
- Daniel, H. Molecular and integrative physiology of intestinal peptide transport. Ann. Rev. Physiol. 2004, 66, 361–384. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Pan, Y.-X.; Wong, E.A.; Webb, K.E. Dietary protein level and stage of development affect expression of an intestinal peptide transporter (cPepT1) in chickens. J. Nutr. 2005, 135, 193–198. [Google Scholar] [CrossRef]
- Zwarycz, B.; Wong, E.A. Expression of the peptide transporters PepT1, PepT2, and PHT1 in the embryonic and posthatch chick. Poult. Sci. 2013, 92, 1314–1321. [Google Scholar] [CrossRef]
- Chen, C.; Yin, Y.; Tu, Q.; Yang, H. Glucose and amino acids in enterocyte: Absorption, metabolism and maturation. Front. Biosci. 2018, 23, 1721–1739. [Google Scholar]
- Spanier, B. Transcriptional and functional regulation of the intestinal peptide transporter PEPT1. J. Physiol. 2014, 592, 871–879. [Google Scholar] [CrossRef]
- Osmanyan, A.K.; Harsini, S.G.; Mahdavi, R.; Fisinin, V.I.; Arkhipova, A.L.; Glazko, T.T.; Kovalchuk, S.N.; Kovosky, G.Y. Intestinal amino acid and peptide transporters in broiler are modulated by dietary amino acids and protein. Amino Acids 2018, 50, 353–357. [Google Scholar] [CrossRef]
- Adibi, S.A. Regulation of expression of the intestinal oligopeptide transporter (Pept-1) in health and disease. Am. J. Physiol.-Gastrointest. Liver Physiol. 2003, 285, G779–G788. [Google Scholar] [CrossRef]
- Tokutake, Y.; Taciak, M.; Sato, K.; Toyomizu, M.; Kikusato, M. Effect of dipeptide on intestinal peptide transporter 1 gene expression: An evaluation using primary cultured chicken intestinal epithelial cells. Am. Sci. J. 2021, 92, e13604. [Google Scholar] [CrossRef] [PubMed]
- Steel, A.; Nussberger, S.; Romero, M.F.; Boron, W.F.; Boyd, C.A.; Hediger, M.A. Stoichiometry and pH dependence of the rabbit proton-dependent oligopeptide transporter PepT1. J. Physiol. 1997, 498, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.J.; Leibach, F.H.; Ganapathy, V.; Thwaites, D.T. Optimal absorptive transport of the dipeptide glycylsarcosine is dependent on functional Na+/H+ exchange activity. Pflügers Archiv. 2002, 445, 139–146. [Google Scholar] [PubMed]
- Adejumo, I.O.; Bryson, B.; Olojede, O.C.; Bedford, M.R.; Adedokun, S.A. Effect of sodium sources and exogenous phytase supplementation on growth performance, nutrient digestibility, and digesta pH of 21-day-old broilers. Poult. Sci. 2021, 100, 101467. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, P.G.; Lynch, P.B.; Caffrey, P.J.; O’Reilly, J.J.; O’Connell, M.K. Measurements of the acid-binding capacity of ingredients used in pig diets. Irish Vet. J. 2005, 58, 447–452. [Google Scholar] [CrossRef]
- Siegert, W.; Krieg, J.; Sommerfeld, V.; Borda-Molina, D.; Feuerstein, D.; Camarinha-Silva, A.; Rodehutscord, M. Phytase supplementation effects on amino acid digestibility in broiler chickens are influenced by dietary calcium concentrations but not by acid-binding capacity. Curr. Dev. Nutr. 2021, 5, nzab103. [Google Scholar] [CrossRef] [PubMed]
- Stas, E.B.; Tokach, M.D.; De Rouchey, J.M.; Goodband, R.D.; Woodworth, J.C.; Gebhardt, J.T. Evaluation of the acid-binding capacity of ingredients and complete diets commonly used for weanling pigs. Trans. Anim. Sci. 2022, 6, 1–9. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Liu, S.; Xie, X.-N.; Tan, Z.-R. Regulation profile of the intestinal peptide transporter (PepT1). Drug Des. Dev. Ther. 2017, 11, 3511–3517. [Google Scholar] [CrossRef]
- Moss, A.F.; Chrystal, P.V.; Dersjant-Li, Y.; Selle, P.H.; Liu, S.Y. The influence of phytase, pre-pellet cracked maize and dietary crude protein level on broiler performance via response surface methodology. J. Anim. Sci. Biotechnol. 2019, 10, 80. [Google Scholar] [CrossRef]
- Barth, H.G.; Boyes, B.E.; Jackson, C. Size exclusion chromatography. Anal. Chem. 1994, 66, 595 R–620 R. [Google Scholar] [CrossRef]
- Liu, S.Y.; Cadogan, D.J.; Péron, A.; Truong, H.H.; Selle, P.H. Effects of phytase supplementation on growth performance, nutrient utilization and digestive dynamics of starch and protein in broiler chickens offered maize-, sorghum- and wheat-based diets. Anim. Feed Sci. Technol. 2014, 197, 164–175. [Google Scholar] [CrossRef]
- Camden, B.J.; Morel, P.C.H.; Thomas, D.V.; Ravindran, V.; Bedford, M.R. Effectiveness of exogenous microbial phytase in improving the bioavailabilities of phosphorus and other nutrients in maize–soya-bean meal diets for broilers. Anim. Sci. 2021, 73, 289–297. [Google Scholar] [CrossRef]
- Truong, H.H.; Liu, S.Y.; Selle, P.H. Phytase influences the inherently different starch digestive dynamics of wheat- and maize-based broiler diets. In Proceedings of the 26th Australian Poultry Science Symposium, Sydney, Australia, 8–11 February 2015; Volume 26, pp. 126–129. [Google Scholar]
- Moran, E.T. Starch digestion in fowl. Poult. Sci. 1982, 61, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Bedford, M.R.; Cowieson, A.J. Matrix values for exogenous enzymes and their application in the real world. J. Appl. Poult. Res. 2020, 29, 15–22. [Google Scholar] [CrossRef]
- Cowieson, A.J.; Wilcock, P.; Bedford, M.R. Super-dosing effects of phytase in poultry and other monogastrics. World’s Poult. Sci. J. 2011, 67, 225–236. [Google Scholar] [CrossRef]
- De Freitas, H.B.; De Souza Nascimento, K.M.R.; Kiefer, C.; Gomes, G.A.; Dos Santos, T.T.; Garcia, E.R.M.; Da Silva, T.R.; Paiva, L.L.; Berno, P.R. Graded levels of phytase on performance, bone mineralization and carcass traits of broiler fed reduced dicalcium phosphate. Asian-Australas. J. Anim. Sci. 2019, 32, 691. [Google Scholar] [CrossRef]
- Moss, A.F.; Ghane, A.; Dersjant-Li, Y.; Dao, T.H.; Suleman, M.; Morgan, N.; Crowley, T.M. Appraisal of matrix values for exogenous phytase alone or in combination with other enzymes in diets for broiler chickens. In Proceedings of the 33rd Australian Poultry Science Symposium, Sydney, Australia, 7–9 February 2022; Volume 33, pp. 85–88. [Google Scholar]
- Lemme, A.; Hiller, P.; Klahsen, M.; Taube, V.; Stegemann, J.; Simon, I. Reduction of dietary protein in broiler diets not only reduces n-emissions but is also accompanied by several further benefits. J. Appl. Poult. Res. 2019, 28, 867–880. [Google Scholar] [CrossRef]
- Greenhalgh, S.; Chrystal, P.V.; Selle, P.H.; Liu, S.Y. Reduced-crude protein diets in chicken-meat production: Justification for an imperative. World’s Poult. Sci. J. 2020, 76, 537–548. [Google Scholar] [CrossRef]
- Alfonso-Avila, A.R.; Cirot, O.; Lambert, W.; Létourneau-Montminy, M.P. Effect of low-protein corn and soybean meal-based diets on nitrogen utilization, litter quality, and water consumption in broiler chicken production: Insight from meta-analysis. Animal 2022, 16, 100458. [Google Scholar] [CrossRef]
- Selle, P.H.; Dorigam, J.C.d.e.P.; Lemme, A.; Chrystal, P.V.; Liu, S.Y. Synthetic and crystalline amino acids: Alternatives to soybean meal in chicken-meat production. Animals 2020, 10, 729. [Google Scholar] [CrossRef] [Green Version]
- Chrystal, P.V.; Moss, A.F.; Khoddami, A.; Naranjo, V.D.; Selle, P.H.; Liu, S.Y. Impacts of reduced-crude protein diets on key parameters in male broiler chickens offered maize-based diets. Poult. Sci. 2020, 99, 505–516. [Google Scholar] [CrossRef]
- Chrystal, P.V.; Moss, A.F.; Khoddami, A.; Naranjo, V.D.; Selle, P.H.; Liu, S.Y. Effects of reduced crude protein levels, dietary electrolyte balance and energy density on the performance of broiler chickens offered maize-based diets with evaluations of starch, protein and amino acid metabolism. Poult. Sci. 2020, 99, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Chrystal, P.V.; Moss, A.F.; Yin, D.; Khoddami, A.; Naranjo, V.D.; Selle, P.H.; Liu, S.Y. Glycine equivalent and threonine inclusions in reduced-crude protein, maize-based diets impact on growth performance, fat deposition starch-protein digestive dynamics and amino acid metabolism in broiler chickens. Anim. Feed Sci. Technol. 2020, 261, 114387. [Google Scholar] [CrossRef]
- Chrystal, P.V.; Greenhalgh, S.; McInerney, B.V.; McQuade, L.R.; Selle, P.H.; Liu, S.Y. Maize-based diets are more conducive to crude protein reductions than wheat-based diets for broiler chickens. Anim. Feed Sci. Technol. 2021, 275, 114867. [Google Scholar] [CrossRef]
- Cabahug, S.; Ravindran, V.; Selle, P.H.; Bryden, W.L. Response of broiler chickens to microbial phytase as influenced by dietary phytic acid and non-phytate phosphorus levels. I. Effects on bird performance and toe ash content. Br. Poult. Sci. 1999, 40, 660–666. [Google Scholar] [CrossRef]
- Jacob, J.P.; Ibrahim, S.; Blair, R.; Namkung, H.; Paok, I.K. Using enzyme supplemented, reduced protein diets to decrease nitrogen and phosphorus excretion of broilers. Asian-Australas. J. Anim. Sci. 2000, 13, 1561–1567. [Google Scholar] [CrossRef]
- Hofmann, P.; Kreig, J.; Francesh, M.; Feuertstein, D.; Rodehutscord, M. Effects of added phytase on growth performance, carcass traits, and tibia ash of broiler chickens fed diets with reduced amino acid, crude protein, and phosphorus concentration. J. Appl. Poult. Res. 2022, 31, 100258. [Google Scholar] [CrossRef]
- Liu, S.Y.; Macelline, S.P.; Chrystal, P.V.; Selle, P.H. Progress towards reduced-crude protein diets for broiler chickens and sustainable chicken-meat production. J. Anim. Sci. Biotech. 2021, 12, 20. [Google Scholar] [CrossRef]
- Donkoh, A.; Moughan, P.J. The effect of dietary crude protein content on apparent and true ileal nitrogen and amino acid digestibilities. Br. Poult. Sci. 1994, 72, 59–68. [Google Scholar] [CrossRef]
- Rutherfurd, S.M.; Chung, T.K.; Moughan, P.J. The effect of microbial phytase on ileal phosphorus and amino acid digestibility in the broiler chicken. Brit. Poult. Sci. 2002, 43, 598–608. [Google Scholar] [CrossRef]
- Lemme, A.; Rostagno, H.S.; Petri, A.; Albino, L.F. Standardised ileal digestibility of crystalline amino acids. In Proceedings of the 15th European Symposium on Poultry Nutrition, Balatonfüred, Hungary, 25–29 September 2005; World’s Poultry Science Association: Beekbergen, The Netherlands, 2005; pp. 462–464. [Google Scholar]
- Rutherfurd, S.M.; Edwards, A.C.E.; Selle, P.H. Effect of phytase on lysine-rice pollard complexes. In Manipulating Pig Production VI; Australasian Pig Science Association: Werribee, Australia, 1999; p. 248. [Google Scholar]


{kind=link}
| Amino Acid | Cowieson et al. [48] | Martinez-Vallespin et al. [47] | Amerah et al. [46] | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Nil | Plus | Response (%) | Nil | 1000 FTU/kg | Response (%) | Nil | 1000 FTU/kg | Response (%) | |
| Arginine | 0.86 | 0.89 | 3.49 | 0.859 | 0.899 | 4.66 | 0.823 | 0.900 | 9.36 |
| Histidine | 0.80 | 0.84 | 5.00 | 0.725 | 0.841 | 16.00 | 0.730 | 0.821 | 12.47 |
| Isoleucine | 0.79 | 0.85 | 7.59 | 0.743 | 0.794 | 6.86 | 0.721 | 0.821 | 13.87 |
| Leucine | 0.82 | 0.85 | 3.66 | 0.749 | 0.826 | 10.28 | 0.722 | 0.826 | 14.40 |
| Lysine | 0.83 | 0.86 | 3.61 | 0.804 | 0.862 | 7.21 | 0.806 | 0.889 | 10.30 |
| Methionine | 0.89 | 0.90 | 1.12 | 0.768 | 0.883 | 14.97 | 0.880 | 0.928 | 5.45 |
| Phenylalanine | 0.82 | 0.85 | 3.66 | 0.806 | 0.858 | 6.45 | 0.737 | 0.834 | 13.16 |
| Threonine | 0.73 | 0.77 | 5.48 | 0.646 | 0.752 | 16.41 | 0.661 | 0.765 | 15.73 |
| Valine | 0.78 | 0.82 | 5.13 | 0.689 | 0.730 | 5.95 | 0.669 | 0.774 | 15.70 |
| Alanine | 0.80 | 0.83 | 3.75 | 0.731 | 0.818 | 11.90 | 0.699 | 0.807 | 15.45 |
| Aspartic acid | 0.79 | 0.82 | 3.80 | 0.796 | 0.835 | 4.90 | 0.756 | 0.850 | 12.43 |
| Cysteine | 0.68 | 0.72 | 5.88 | 0.478 | 0.681 | 42.47 | 0.543 | 0.685 | 26.15 |
| Glutamic acid | 0.87 | 0.90 | 3.45 | 0.833 | 0.888 | 6.60 | 0.807 | 0.882 | 9.29 |
| Glycine | 0.76 | 0.79 | 3.95 | 0.636 | 0.749 | 17.77 | 0.658 | 0.765 | 16.26 |
| Proline | 0.80 | 0.83 | 3.75 | 0.789 | 0.809 | 2.53 | 0.722 | 0.812 | 12.47 |
| Serine | 0.78 | 0.81 | 3.85 | 0.723 | 0.805 | 11.34 | 0.684 | 0.809 | 18.27 |
| Tyrosine | 0.80 | 0.83 | 3.75 | 0.767 | 0.813 | 6.00 | 0.755 | 0.844 | 11.79 |
| Mean | 0.800 | 0.833 | 4.13 | 0.738 | 0.814 | 10.30 | 0.728 | 0.824 | 13.19 |
| Feed Grains | Vegetable Protein Meals | ||
|---|---|---|---|
| Source | iP | Source | iP |
| Maize–zein | 6.2 | Cottonseed–α-globulin | 5.5 |
| Rye–gliadin | 6.6 | Cottonseed–ß-globulin | 5.4 |
| Sorghum–kafirin | 5.9 | Navy bean–phaseolin | 4.5 |
| Wheat–gliadin | 6.5 | Peanut–arachnin | 5.4 |
| Wheat bran–prolamin | 6.2 | Soybean–glycinin | 4.7 |
| Mean | 6.28 | Mean | 5.10 |
| Amino Acid | Proximal Jejunum | Distal Jejunum | Proximal Ileum | Distal Ileum | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 FTU/kg | 500 FTU | Response | 0 FTU/kg | 500 FTU | Response | 0 FTU/kg | 500 FTU | Response | 0 FTU/kg | 500 FTU | Response | |
| Arginine | 0.609 | 0.798 | 31.0% | 0.770 | 0.869 | 12.9% | 0.872 | 0.922 | 5.73% | 0.901 | 0.942 | 4.55% |
| Histidine | 0.481 | 0.723 | 50.3% | 0.678 | 0.813 | 19.9% | 0.824 | 0.889 | 7.89% | 0.862 | 0.915 | 6.15% |
| Isoleucine | 0.455 | 0.700 | 53.9% | 0.656 | 0.789 | 20.3% | 0.797 | 0.870 | 9.16% | 0.839 | 0.900 | 7.27% |
| Leucine | 0.474 | 0.700 | 47.7% | 0.664 | 0.792 | 19.3% | 0.817 | 0.881 | 7.83% | 0.856 | 0.910 | 6.31% |
| Lysine | 0.684 | 0.848 | 24.0% | 0.799 | 0.890 | 11.4% | 0.867 | 0.923 | 6.46% | 0.881 | 0.934 | 6.02% |
| Methionine | 0.700 | 0.850 | 21.4% | 0.843 | 0.917 | 8.78% | 0.920 | 0.957 | 4.02% | 0.948 | 0.970 | 2.32% |
| Phenylalanine | 0.509 | 0.720 | 41.5% | 0.682 | 0.803 | 17.7% | 0.820 | 0.885 | 7.93% | 0.860 | 0.914 | 6.28% |
| Threonine | 0.394 | 0.681 | 72.8% | 0.588 | 0.758 | 28.9% | 0.733 | 0.836 | 14.1% | 0.767 | 0.861 | 12.3% |
| Valine | 0.403 | 0.664 | 64.8% | 0.613 | 0.758 | 23.7% | 0.774 | 0.849 | 9.69% | 0.813 | 0.878 | 8.00% |
| Alanine | 0.465 | 0.699 | 50.3% | 0.656 | 0.788 | 20.1% | 0.801 | 0.870 | 8.61% | 0.831 | 0.895 | 7.70% |
| Aspartate | 0.429 | 0.698 | 62.7% | 0.643 | 0.787 | 22.4% | 0.776 | 0.861 | 11.0% | 0.817 | 0.888 | 8.69% |
| Glutamate | 0.554 | 0.762 | 37.6% | 0.731 | 0.844 | 15.5% | 0.851 | 0.907 | 6.58% | 0.884 | 0.929 | 5.09% |
| Glycine | 0.390 | 0.675 | 73.1% | 0.587 | 0.756 | 28.8% | 0.745 | 0.839 | 12.6% | 0.787 | 0.868 | 10.3% |
| Proline | 0.415 | 0.685 | 65.1% | 0.621 | 0.778 | 25.3% | 0.793 | 0.869 | 9.58% | 0.837 | 0.898 | 7.29% |
| Serine | 0.455 | 0.704 | 54.7% | 0.639 | 0.783 | 22.5% | 0.776 | 0.861 | 11.0% | 0.819 | 0.890 | 8.67% |
| Tyrosine | 0.277 | 0.605 | 118% | 0.800 | 0.712 | 42.4% | 0.717 | 0.830 | 15.8% | 0.778 | 0.875 | 12.5% |
| Mean | 0.481 | 0.720 | 49.7% | 0.667 | 0.802 | 20.2% | 0.805 | 0.878 | 9.07% | 0.843 | 0.904 | 7.24% |
| Item (g/kg) | Starter Diet 1 to 10 Days | Grower Diet 11 to 21 Days | Finisher Diet 22 to 35 Days | Withdrawal Diet 36 to 42 Days |
|---|---|---|---|---|
| Phosphorus | 1.97 | 1.97 | 1.97 | 1.97 |
| Calcium | 2.09 | 2.09 | 2.09 | 2.09 |
| Sodium | 0.43 | 0.43 | 0.43 | 0.43 |
| Crude protein | 6.04 | 5.29 | 4.59 | 4.59 |
| Lysine | 0.34 | 0.30 | 0.23 | 0.26 |
| Methionine | 0.10 | 0.09 | 0.07 | 0.08 |
| Cysteine | 0.19 | 0.16 | 0.14 | 0.15 |
| Threonine | 0.28 | 0.25 | 0.19 | 0.22 |
| Isoleucine | 0.27 | 0.22 | 0.19 | 0.19 |
| Leucine | 0.59 | 0.46 | 0.44 | 0.44 |
| Tryptophan | 0.08 | 0.07 | 0.06 | 0.07 |
| Proline | 0.41 | 0.32 | 0.26 | 0.26 |
| Serine | 0.37 | 0.29 | 0.24 | 0.24 |
| Valine | 0.34 | 0.27 | 0.24 | 0.24 |
| Arginine | 0.27 | 0.21 | 0.18 | 0.18 |
| Energy (MJ/kg) | 0.29 | 0.28 | 0.27 | 0.28 |
| Diet | Control Diet (AUD/tonne) | Phytase Diet (AUD/tonne) | Reduction (AUD/tonne) | Proportion of Intake (%) |
|---|---|---|---|---|
| Starter (1 to 10 days) | 648.14 | 572.15 | 76.02 | 6.0 |
| Grower (11 to 21 days) | 632.87 | 558.56 | 74.31 | 18.4 |
| Finisher (22 to 35 days) | 603.28 | 539.68 | 63.60 | 45.0 |
| Withdrawal (36 to 42 days) | 572.31 | 511.46 | 60.85 | 30.6 |
| Weighted mean | 601.94 | 536.74 | 65.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selle, P.H.; Macelline, S.P.; Chrystal, P.V.; Liu, S.Y. The Contribution of Phytate-Degrading Enzymes to Chicken-Meat Production. Animals 2023, 13, 603. https://doi.org/10.3390/ani13040603
Selle PH, Macelline SP, Chrystal PV, Liu SY. The Contribution of Phytate-Degrading Enzymes to Chicken-Meat Production. Animals. 2023; 13(4):603. https://doi.org/10.3390/ani13040603
Chicago/Turabian StyleSelle, Peter H., Shemil P. Macelline, Peter V. Chrystal, and Sonia Yun Liu. 2023. "The Contribution of Phytate-Degrading Enzymes to Chicken-Meat Production" Animals 13, no. 4: 603. https://doi.org/10.3390/ani13040603
APA StyleSelle, P. H., Macelline, S. P., Chrystal, P. V., & Liu, S. Y. (2023). The Contribution of Phytate-Degrading Enzymes to Chicken-Meat Production. Animals, 13(4), 603. https://doi.org/10.3390/ani13040603