
Genetic Diversity and Trends of Ancestral and New Inbreeding in German Sheep Breeds by Pedigree Data

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peter, C. Molekulargenetische Charakterisierung von Schafrassen Europas und des Nahen Ostens auf der Basis von Mikrosatelliten; VVB Laufersweiler: Gießen, Germany, 2006. [Google Scholar]
- Rochus, C.M.; Johansson, A.M. Estimation of genetic diversity in Gute sheep: Pedigree and microsatellite analyses of an ancient Swedish breed. Hereditas 2017, 154, 4. [Google Scholar] [CrossRef] [PubMed]
- Lawson Handley, L.; Byrne, K.; Santucci, F.; Townsend, S.; Taylor, M.; Bruford, M.W.; Hewitt, G. Genetic structure of European sheep breeds. Heredity 2007, 99, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Galal, S.; Boyazoglu, J. Animal Genetic Resources Information; Publishing and Multimedia Service, Information Division, FAO: Rome, Italy, 1997. [Google Scholar]
- Frankham, R. Conservation genetics. Annu. Rev. Genet. 1995, 29, 305–327. [Google Scholar] [CrossRef] [PubMed]
- Illa, S.K.; Gollamoori, G.; Nath, S. Evaluation of selection program by assessing the genetic diversity and inbreeding effects on Nellore sheep growth through pedigree analysis. Asian-Australas J. Anim. Sci. 2019, 33, 1369–1377. [Google Scholar] [CrossRef]
- Oravcova, M.; Krupa, E. Pedigree analysis of the former Valachian sheep. Slovak J. Anim. Sci. 2011, 44, 6–12. [Google Scholar]
- Addo, S.; Klingel, S.; Thaller, G.; Hinrichs, D. Genetic diversity and the application of runs of homozygosity-based methods for inbreeding estimation in German White-headed Mutton sheep. PLoS ONE 2021, 16, e0250608. [Google Scholar] [CrossRef]
- Vozzi, P.; Marcondes, C.; Bezerra, L.; Lôbo, R. Pedigree analyses in the breeding program for nellore cattle. Genet. Mol. Res. 2007, 6, 1044–1050. [Google Scholar]
- Antonios, S.; Rodríguez-Ramilo, S.T.; Aguilar, I.; Astruc, J.M.; Legarra, A.; Vitezica, Z.G. Genomic and pedigree estimation of inbreeding depression for semen traits in the Basco-Béarnaise dairy sheep breed. J. Dairy Sci. 2021, 104, 3221–3230. [Google Scholar] [CrossRef]
- Moreno, A.; Salgado, C.; Piqueras, P.; Gutiérrez, J.; Toro, M.; Ibáñez-Escriche, N.; Nieto, B. Restricting inbreeding while maintaining selection response for weight gain in Mus musculus. J. Anim. Breed. Genet. 2011, 128, 276–283. [Google Scholar] [CrossRef]
- Gizaw, S.; Getachew, T.; Haile, A.; Rischkowsky, B.; Sölkner, J.; Tibbo, M. Optimization of selection for growth in Menz sheep while minimizing inbreeding depression in fitness traits. Genet. Sel. Evol. 2013, 45, 20. [Google Scholar] [CrossRef]
- Goyache, F.; Gutiérrez, J.P.; Fernández, I.; Gomez, E.; Alvarez, I.; Díez, J.; Royo, L.J. Using pedigree information to monitor genetic variability of endangered populations: The Xalda sheep breed of Asturias as an example. J. Anim. Breed. Genet. 2003, 120, 95–105. [Google Scholar] [CrossRef]
- Pedrosa, V.; Santana Jr, M.; Oliveira, P.; Eler, J.; Ferraz, J. Population structure and inbreeding effects on growth traits of Santa Inês sheep in Brazil. Small Rumin. Res. 2010, 93, 135–139. [Google Scholar] [CrossRef]
- Ghafouri-Kesbi, F. Analysis of genetic diversity in a close population of Zandi sheep using genealogical information. J. Genet. 2010, 89, 479. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Kesbi, F. Using pedigree information to study genetic diversity and re-evaluating a selection program in an experimental flock of Afshari sheep. Arch. Anim. Breed. 2012, 55, 375–384. [Google Scholar] [CrossRef]
- Notter, D.R. The importance of genetic diversity in livestock populations of the future. J. Anim. Sci. 1999, 77, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Melka, M.; Stachowicz, K.; Miglior, F.; Schenkel, F. Analyses of genetic diversity in five Canadian dairy breeds using pedigree data. J. Anim. Breed. Genet. 2013, 130, 476–486. [Google Scholar] [CrossRef]
- Boichard, D. Pedig: A fortran package for pedigree analysis suited for large populations. In Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, 19–23 August 2002. [Google Scholar]
- Maignel, L.; Boichard, D.; Verrier, E. Genetic variability of French dairy breeds estimated from pedigree information. Interbull Bull. 1996, 16, 49. [Google Scholar]
- James, J. A note on selection differential and generation length when generations overlap. Anim. Sci. 1977, 24, 109–112. [Google Scholar] [CrossRef]
- Boichard, D.; Maignel, L.; Verrier, E. The value of using probabilities of gene origin to measure genetic variability in a population. Genet. Sel. Evol. 1997, 29, 5–23. [Google Scholar] [CrossRef]
- Lacy, R.C. Analysis of founder representation in pedigrees: Founder equivalents and founder genome equivalents. Zoo Biology 1989, 8, 111–123. [Google Scholar] [CrossRef]
- Lacy, R.C. Clarification of Genetic Terms and Their Use in the Management of Captive Populations; Wiley Online Library: Hoboken, NJ, USA, 1995. [Google Scholar]
- Meuwissen, T.; Luo, Z. Computing inbreeding coefficients in large populations. Genet. Sel. Evol. 1992, 24, 305–313. [Google Scholar] [CrossRef]
- Ballou, J. Ancestral inbreeding only minimally affects inbreeding depression in mammalian populations. J. Hered. 1997, 88, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, S.T.; Hedrick, P.W.; Miller, P.S. Inbreeding depression in the Speke’s gazelle captive breeding program. Conserv. Biol. 2000, 14, 1375–1384. [Google Scholar] [CrossRef]
- Baumung, R.; Farkas, J.; Boichard, D.; Mészáros, G.; Sölkner, J.; Curik, I. GRAIN: A computer program to calculate ancestral and partial inbreeding coefficients using a gene dropping approach. J. Anim. Breed. Genet. 2015, 132, 100–108. [Google Scholar] [CrossRef]
- Doekes, H.P.; Curik, I.; Nagy, I.; Farkas, J.; Kövér, G.; Windig, J.J. Revised calculation of Kalinowski’s ancestral and new inbreeding coefficients. Diversity 2020, 12, 155. [Google Scholar] [CrossRef]
- Caballero, A.; Toro, M.A. Interrelations between effective population size and other pedigree tools for the management of conserved populations. Genet. Res. 2000, 75, 331–343. [Google Scholar] [CrossRef]
- Gutiérrez, J.; Cervantes, I.; Goyache, F. Improving the estimation of realized effective population sizes in farm animals. J. Anim. Breed. Genet. 2009, 126, 327–332. [Google Scholar] [CrossRef]
- Cervantes, I.; Goyache, F.; Molina, A.; Valera, M.; Gutiérrez, J. Application of individual increase in inbreeding to estimate realized effective sizes from real pedigrees. J. Anim. Breed. Genet. 2008, 125, 301–310. [Google Scholar] [CrossRef]
- Leroy, G.; Baumung, R. Mating practices and the dissemination of genetic disorders in domestic animals, based on the example of dog breeding. Anim. Genet. 2011, 42, 66–74. [Google Scholar] [CrossRef]
- Baumung, R.; Simianer, H.; Hoffmann, I. Genetic diversity studies in farm animals—A survey. J. Anim. Breed. Genet. 2004, 121, 361–373. [Google Scholar] [CrossRef]
- Mokhtari, M.; Shahrbabak, M.M.; Esmailizadeh, A.; Abdollahi-Arpanahi, R.; Gutierrez, J. Genetic diversity in Kermani sheep assessed from pedigree analysis. Small Rumin. Res. 2013, 114, 202–205. [Google Scholar] [CrossRef]
- Leroy, G.; Verrier, E.; Meriaux, J.C.; Rognon, X. Genetic diversity of dog breeds: Within-breed diversity comparing genealogical and molecular data. Anim. Genet. 2009, 40, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Cassell, B.G.; Adamec, V.; Pearson, R.E. Effect of incomplete pedigrees on estimates of inbreeding and inbreeding depression for days to first service and summit milk yield in Holsteins and Jerseys. J. Dairy Sci. 2003, 86, 2967–2976. [Google Scholar] [CrossRef] [Green Version]
- Danchin-Burge, C.; Palhiere, I.; François, D.; Bibé, B.; Leroy, G.; Verrier, E. Pedigree analysis of seven small French sheep populations and implications for the management of rare breeds. J. Anim. Sci. 2010, 88, 505–516. [Google Scholar] [CrossRef]
- Rodríguez-Ramilo, S.T.; Elsen, J.M.; Legarra, A. Inbreeding and effective population size in French dairy sheep: Comparison between genomic and pedigree estimates. J. Dairy Sci. 2019, 102, 4227–4237. [Google Scholar] [CrossRef] [PubMed]
- Meyermans, R.; Gorssen, W.; Wijnrocx, K.; Lenstra, J.; Vellema, P.; Buys, N.; Janssens, S. Unraveling the genetic diversity of Belgian Milk Sheep using medium-density SNP genotypes. Anim. Genet. 2020, 51, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, M.; Shahrbabak, M.M.; Esmailizadeh, A.; Shahrbabak, H.M.; Gutierrez, J. Pedigree analysis of Iran-Black sheep and inbreeding effects on growth and reproduction traits. Small Rumin. Res. 2014, 116, 14–20. [Google Scholar] [CrossRef]
- Tahmoorespur, M.; Sheikhloo, M. Pedigree analysis of the closed nucleus of Iranian Baluchi sheep. Small Rumin. Res. 2011, 99, 1–6. [Google Scholar] [CrossRef]
- Sheikhlou, M.; Abbasi, M. Genetic diversity of Iranian Lori-Bakhtiari sheep assessed by pedigree analysis. Small Rumin. Res. 2016, 141, 99–105. [Google Scholar] [CrossRef]
- Gutiérrez, J.P.; Altarriba, J.; Díaz, C.; Quintanilla, R.; Cañón, J.; Piedrafita, J. Pedigree analysis of eight Spanish beef cattle breeds. Genet. Sel. Evol. 2003, 35, 43–63. [Google Scholar] [CrossRef]
- Mallick, P.; Chauhan, I.; Thirumaran, S.; Pourouchttamane, R.; Kumar, A. Genetic variability of Bharat merino sheep derived from pedigree information. Indian J. Anim. Res. 2020, 54, 1324–1331. [Google Scholar] [CrossRef]
- De Vries, F.; Hamann, H.; Distl, O. Auswirkungen verschiedener Strategien der Zucht auf Scrapie-Resistenz auf Inzucht-entwicklung in kleinen Populationen. Züchtungskunde 2006, 78, 28–43. [Google Scholar]
- Windig, J.; Kaal, L. An effective rotational mating scheme for inbreeding reduction in captive populations illustrated by the rare sheep breed Kempisch Heideschaap. Animal 2008, 2, 1733–1741. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.L.; Buchenauer, D. Genetic Diversity of European Livestock Breeds; Wageningen Pers: Wageningen, The Netherlands, 1993. [Google Scholar]








{kind=link}
{kind=link}
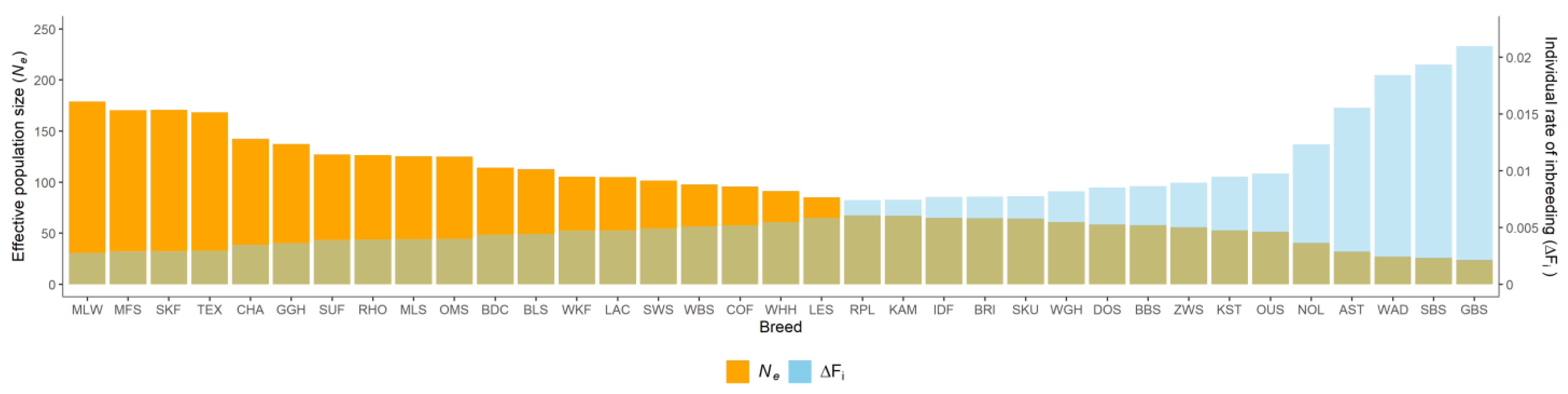{kind=link}
{kind=link}
{kind=link}
{kind=link}
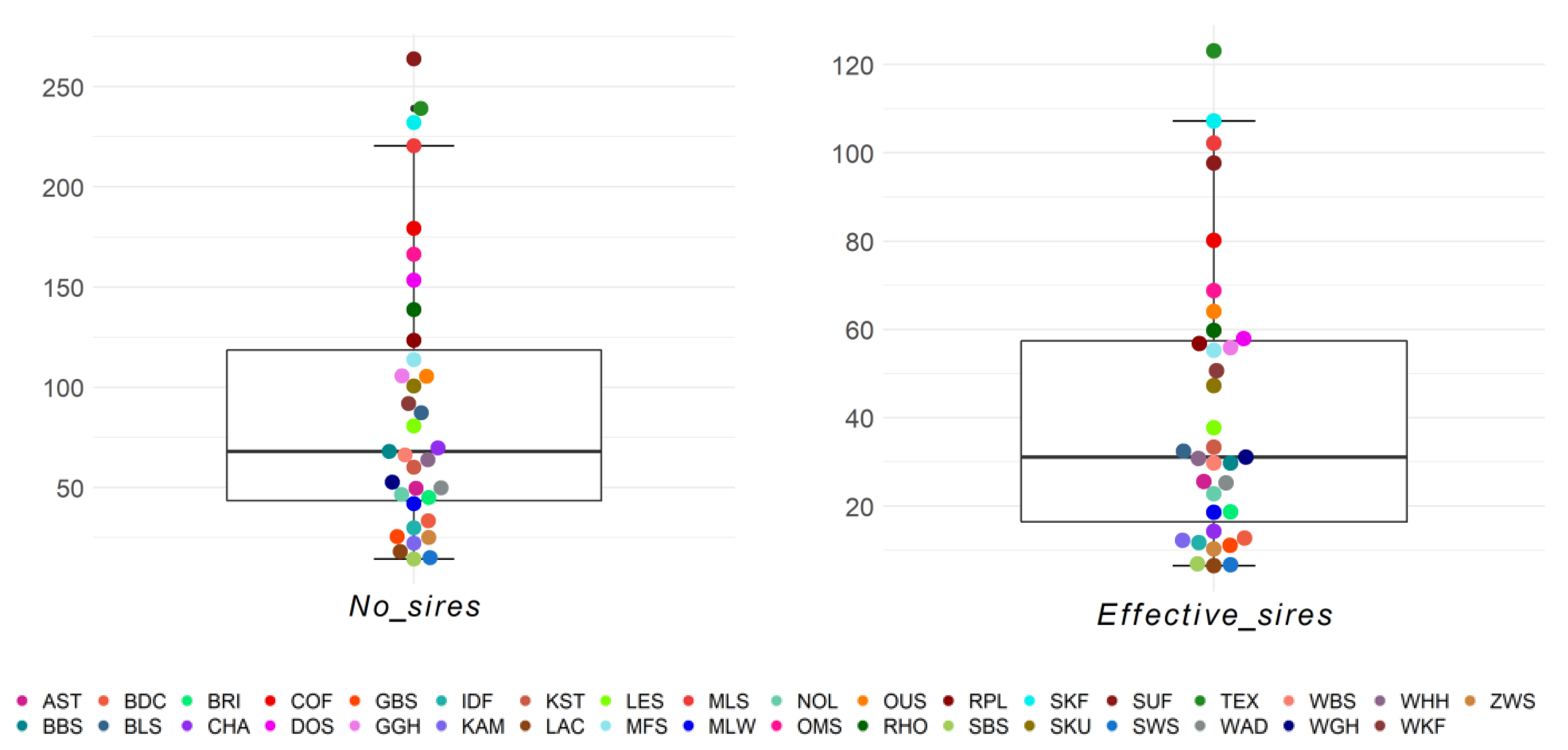{kind=link}
| Code | BD | Sheep Breed | NPed | NRef | GE | GI | f | fe | fg | fa |
|---|---|---|---|---|---|---|---|---|---|---|
| AST | MON | Alpine Steinschaf | 10,420 | 8729 | 4.71 | 3.70 | 286 | 24.1 | 10.7 | 17.0 |
| BBS | MON | Brown Mountain | 22,961 | 13,510 | 5.79 | 4.06 | 784 | 89.2 | 25.0 | 53.6 |
| BDC | EXO | Berrichon du Cher | 4680 | 3101 | 3.51 | 3.45 | 491 | 129.7 | 46.1 | 74.7 |
| BLS | CON | Bentheim | 46,173 | 29,973 | 7.83 | 3.76 | 1064 | 68.8 | 17.9 | 41.8 |
| BRI | CON | Carinthian | 10,669 | 6386 | 4.07 | 3.79 | 669 | 99.8 | 32.7 | 60.7 |
| CHA | MEA | Charollais | 11,237 | 8044 | 3.08 | 3.21 | 1039 | 150.5 | 64.6 | 81.2 |
| COF | CON | Coburg | 70,156 | 43,072 | 7.30 | 3.82 | 2828 | 177.4 | 31.9 | 76.6 |
| DOS | MEA | Dorper | 36,057 | 28,347 | 5.72 | 3.43 | 1380 | 41.1 | 17.8 | 28.4 |
| GBS | MON | Baraka | 3938 | 3334 | 2.55 | 3.54 | 272 | 45.4 | 27.2 | 43.1 |
| GGH | HEA | German Grey Heath | 69,369 | 41,982 | 7.39 | 3.92 | 3226 | 156.8 | 25.9 | 60.0 |
| IDF | MEA | Ile-de-France | 14,021 | 9066 | 3.77 | 3.94 | 696 | 140.9 | 49.5 | 75.0 |
| KAM | EXO | Kamerun | 7404 | 3313 | 4.59 | 3.99 | 281 | 46.6 | 18.0 | 35.2 |
| KST | MON | Krainer Steinschaf | 9671 | 7234 | 5.41 | 3.87 | 399 | 52.9 | 18.1 | 29.4 |
| LAC | MIL | Lacaune | 4652 | 3632 | 3.09 | 3.17 | 444 | 58.4 | 24.0 | 36.6 |
| LES | CON | Leine | 42,949 | 37,108 | 7.44 | 3.83 | 1348 | 104.4 | 24.8 | 56.4 |
| MFS | MER | German Mutton Merino | 132,413 | 85020 | 6.20 | 4.07 | 8500 | 324.5 | 42.5 | 91.1 |
| MLS | MER | German Merino | 204,494 | 172,172 | 8.30 | 4.02 | 5546 | 172.8 | 29.9 | 67.4 |
| MLW | MER | Merino Longwool | 61,216 | 30,546 | 6.42 | 3.82 | 1959 | 109.3 | 26.8 | 61.0 |
| NOL | EXO | Nolana | 11,920 | 8073 | 3.68 | 3.57 | 821 | 46.6 | 24.0 | 40.9 |
| OMS | MIL | East Friesian | 71,159 | 42,427 | 7.98 | 3.08 | 2644 | 253.3 | 42.1 | 102.6 |
| OUS | EXO | Ouessant | 10,051 | 4927 | 6.43 | 3.67 | 638 | 57.9 | 25.6 | 44.6 |
| RHO | CON | Rhön | 78,095 | 45,228 | 6.20 | 4.20 | 3618 | 183.2 | 49.5 | 121.3 |
| RPL | CON | Pomeranian Coarsewool | 56,965 | 31,288 | 6.78 | 4.43 | 940 | 74.9 | 22.8 | 46.5 |
| SBS | MON | Black Mountain | 3903 | 2110 | 3.42 | 3.73 | 161 | 25.5 | 10.7 | 15.3 |
| SKF | MEA | German Blackhead Mutton | 128,839 | 104,178 | 7.71 | 3.92 | 4554 | 186.4 | 49.5 | 104.4 |
| SKU | HEA | Skudde | 32,747 | 16,496 | 6.03 | 4.34 | 1431 | 156.6 | 40.7 | 85.2 |
| SUF | MEA | Suffolk | 68,136 | 54,635 | 5.17 | 3.54 | 4260 | 477.7 | 125.6 | 225.6 |
| SWS | MEA | Swifter | 4608 | 3309 | 3.78 | 3.03 | 235 | 69.6 | 21.1 | 31.9 |
| TEX | MEA | Texel | 58,223 | 45,535 | 5.76 | 6.42 | 2860 | 26.3 | 106.2 | 184.0 |
| WAD | CON | Wald | 17,172 | 9801 | 5.49 | 4.96 | 470 | 40.5 | 15.2 | 29.4 |
| WBS | MON | White Mountain | 30,188 | 19,491 | 8.05 | 3.77 | 1103 | 90.6 | 16.0 | 41.7 |
| WGH | HEA | German White Heath | 18,158 | 12,160 | 7.42 | 3.74 | 667 | 48.7 | 10.8 | 24.4 |
| WHH | HEA | White Polled Heath | 41,306 | 23,289 | 8.68 | 3.85 | 1525 | 65.5 | 14.3 | 40.2 |
| WKF | MEA | German Whitehead Mutton | 38,390 | 21,464 | 7.52 | 3.60 | 871 | 113.4 | 26.6 | 57.8 |
| ZWS | EXO | Zwartbles | 3222 | 2113 | 4.52 | 3.01 | 402 | 102 | 20.6 | 38.9 |
| Breed | Lossfounder (%) | Lossdrift (%) |
|---|---|---|
| AST | 44.56 | 55.44 |
| BBS | 27.99 | 72.01 |
| BDC | 35.51 | 64.49 |
| BLS | 26.02 | 73.98 |
| BRI | 32.73 | 67.27 |
| CHA | 42.92 | 57.08 |
| COF | 17.99 | 82.01 |
| DOS | 43.41 | 56.59 |
| GBS | 59.80 | 40.20 |
| GGH | 16.52 | 83.48 |
| IDF | 35.15 | 64.85 |
| KAM | 38.67 | 61.33 |
| KST | 34.12 | 65.88 |
| LAC | 41.11 | 58.89 |
| LES | 23.74 | 76.26 |
| MFS | 13.11 | 86.89 |
| MLS | 17.27 | 82.73 |
| MLW | 24.49 | 75.51 |
| NOL | 51.42 | 48.58 |
| OMS | 16.61 | 83.39 |
| OUS | 44.25 | 55.75 |
| RHO | 27.04 | 72.96 |
| RPL | 30.47 | 69.53 |
| SBS | 41.96 | 58.04 |
| SKF | 26.53 | 73.47 |
| SKU | 25.96 | 74.04 |
| SUF | 26.29 | 73.71 |
| SWS | 30.29 | 69.71 |
| TEX | 32.55 | 67.45 |
| WAD | 37.41 | 62.59 |
| WBS | 17.67 | 82.33 |
| WGH | 22.24 | 77.76 |
| WHH | 21.80 | 78.20 |
| WKF | 23.44 | 76.56 |
| ZWS | 20.15 | 79.85 |
| Breed | F | Finbred | Inbred | Φ | α |
|---|---|---|---|---|---|
| AST | 0.058 | 0.073 | 0.792 | 0.139 | 0.025 |
| BBS | 0.040 | 0.051 | 0.791 | 0.087 | 0.010 |
| BDC | 0.011 | 0.048 | 0.217 | 0.008 | 0.000 |
| BLS | 0.030 | 0.034 | 0.880 | 0.104 | 0.009 |
| BRI | 0.024 | 0.045 | 0.533 | 0.044 | 0.003 |
| CHA | 0.008 | 0.070 | 0.113 | 0.005 | 0.000 |
| COF | 0.032 | 0.041 | 0.786 | 0.115 | 0.010 |
| DOS | 0.038 | 0.052 | 0.736 | 0.114 | 0.015 |
| GBS | 0.027 | 0.087 | 0.305 | 0.027 | 0.002 |
| GGH | 0.024 | 0.031 | 0.771 | 0.075 | 0.006 |
| IDF | 0.020 | 0.100 | 0.196 | 0.024 | 0.001 |
| KAM | 0.027 | 0.045 | 0.599 | 0.103 | 0.007 |
| KST | 0.041 | 0.059 | 0.698 | 0.107 | 0.015 |
| LAC | 0.013 | 0.056 | 0.228 | 0.009 | 0.001 |
| LES | 0.036 | 0.042 | 0.860 | 0.124 | 0.012 |
| MFS | 0.017 | 0.027 | 0.614 | 0.055 | 0.003 |
| MLS | 0.029 | 0.031 | 0.921 | 0.099 | 0.008 |
| MLW | 0.016 | 0.018 | 0.854 | 0.032 | 0.002 |
| NOL | 0.035 | 0.062 | 0.561 | 0.057 | 0.008 |
| OMS | 0.029 | 0.036 | 0.795 | 0.090 | 0.008 |
| OUS | 0.047 | 0.058 | 0.809 | 0.190 | 0.019 |
| RHO | 0.020 | 0.026 | 0.767 | 0.062 | 0.004 |
| RPL | 0.041 | 0.043 | 0.925 | 0.123 | 0.014 |
| SBS | 0.049 | 0.096 | 0.510 | 0.052 | 0.008 |
| SKF | 0.019 | 0.020 | 0.932 | 0.073 | 0.003 |
| SKU | 0.037 | 0.048 | 0.765 | 0.132 | 0.012 |
| SUF | 0.016 | 0.029 | 0.535 | 0.034 | 0.002 |
| SWS | 0.015 | 0.052 | 0.276 | 0.013 | 0.001 |
| TEX | 0.014 | 0.021 | 0.645 | 0.034 | 0.002 |
| WAD | 0.079 | 0.097 | 0.819 | 0.159 | 0.033 |
| WBS | 0.037 | 0.046 | 0.803 | 0.124 | 0.014 |
| WGH | 0.054 | 0.065 | 0.826 | 0.178 | 0.026 |
| WHH | 0.042 | 0.050 | 0.834 | 0.157 | 0.017 |
| WKF | 0.030 | 0.033 | 0.910 | 0.074 | 0.006 |
| ZWS | 0.032 | 0.078 | 0.408 | 0.051 | 0.008 |
| Parameter | F | Fa_Bal | Fa_Kal | Fa_New | AHC | Lossdrift | Lossfounder |
|---|---|---|---|---|---|---|---|
| F | - | 0.795 * | 0.943 * | 0.941 * | 0.790 * | 0.676 * | 0.645 * |
| Inbred | 0.492 * | 0.733 * | 0.544 * | 0.424 * | 0.697 * | 0.366 * | −0.025 |
| Finbred | 0.496 * | 0.735 * | 0.548 * | 0.425 * | 0.700 * | 0.374 * | −0.020 |
| 0.649 * | 0.181 | 0.440 * | 0.755 * | 0.192 | 0.287 | 0.799 * | |
| Ne | −0.699 * | −0.367 * | −0.539 * | −0.756 * | −0.379 * | −0.401 * | −0.716 * |
| NeffS | −0.144 | 0.151 | −0.059 | −0.177 | 0.144 | −0.367 * | −0.515 * |
| NeffD | −0.213 | 0.070 | −0.139 | −0.254 | 0.044 | −0.189 | −0.490 * |
| Lossdrift | - | 0.614 * | 0.694 * | 0.580 * | 0.594 * | - | - |
| Lossfounder | - | 0.290 | 0.525 * | 0.681 * | 0.300 | 0.600* | - |
| Parameter | f | fe | fg | fa |
|---|---|---|---|---|
| F | −0.349 * | −0.513 * | −0.560 * | −0.523 * |
| Inbred | 0.271 | −0.012 | −0.238 | −0.030 |
| Finbred | 0.268 | −0.021 | −0.249 | −0.041 |
| −0.500 * | −0.531 * | −0.393 * | −0.480 * | |
| Ne | 0.687 * | 0.652 * | 0.519 * | 0.595 * |
| Fa_Bal | −0.092 | −0.327 | −0.461 * | −0.349 * |
| Fa_Kal | −0.268 | −0.439 * | −0.501 * | −0.456 * |
| Fa_New | −0.389 * | −0.521 * | −0.543 * | −0.518 * |
| AHC | −0.112 | −0.333 | −0.449 * | −0.350 * |
| Lossdrift (%) | 0.553 * | 0.416 * | 0.038 | 0.229 |
| Lossfounder (%) | −0.553 * | −0.416 * | −0.038 | −0.229 |
| Lossdrift | −0.331 * | −0.595 * | −0.726 * | −0.682 * |
| Lossfounder | −0.567 * | −0.702 * | −0.581 * | −0.667 * |
| NeffS | 0.682 * | 0.697 * | 0.583 * | 0.705 * |
| NeffD | 0.870 * | 0.536 * | 0.273 | 0.419 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Justinski, C.; Wilkens, J.; Distl, O. Genetic Diversity and Trends of Ancestral and New Inbreeding in German Sheep Breeds by Pedigree Data. Animals 2023, 13, 623. https://doi.org/10.3390/ani13040623
Justinski C, Wilkens J, Distl O. Genetic Diversity and Trends of Ancestral and New Inbreeding in German Sheep Breeds by Pedigree Data. Animals. 2023; 13(4):623. https://doi.org/10.3390/ani13040623
Chicago/Turabian StyleJustinski, Cathrin, Jens Wilkens, and Ottmar Distl. 2023. "Genetic Diversity and Trends of Ancestral and New Inbreeding in German Sheep Breeds by Pedigree Data" Animals 13, no. 4: 623. https://doi.org/10.3390/ani13040623
APA StyleJustinski, C., Wilkens, J., & Distl, O. (2023). Genetic Diversity and Trends of Ancestral and New Inbreeding in German Sheep Breeds by Pedigree Data. Animals, 13(4), 623. https://doi.org/10.3390/ani13040623