


Susceptibility to SARS-CoV-2 and MERS-CoV in Beagle Dogs
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Animal Studies
2.3. Blood Biochemistry and Hematological Examination
2.4. Histopathology
2.5. Quantitative Real-Time PCR
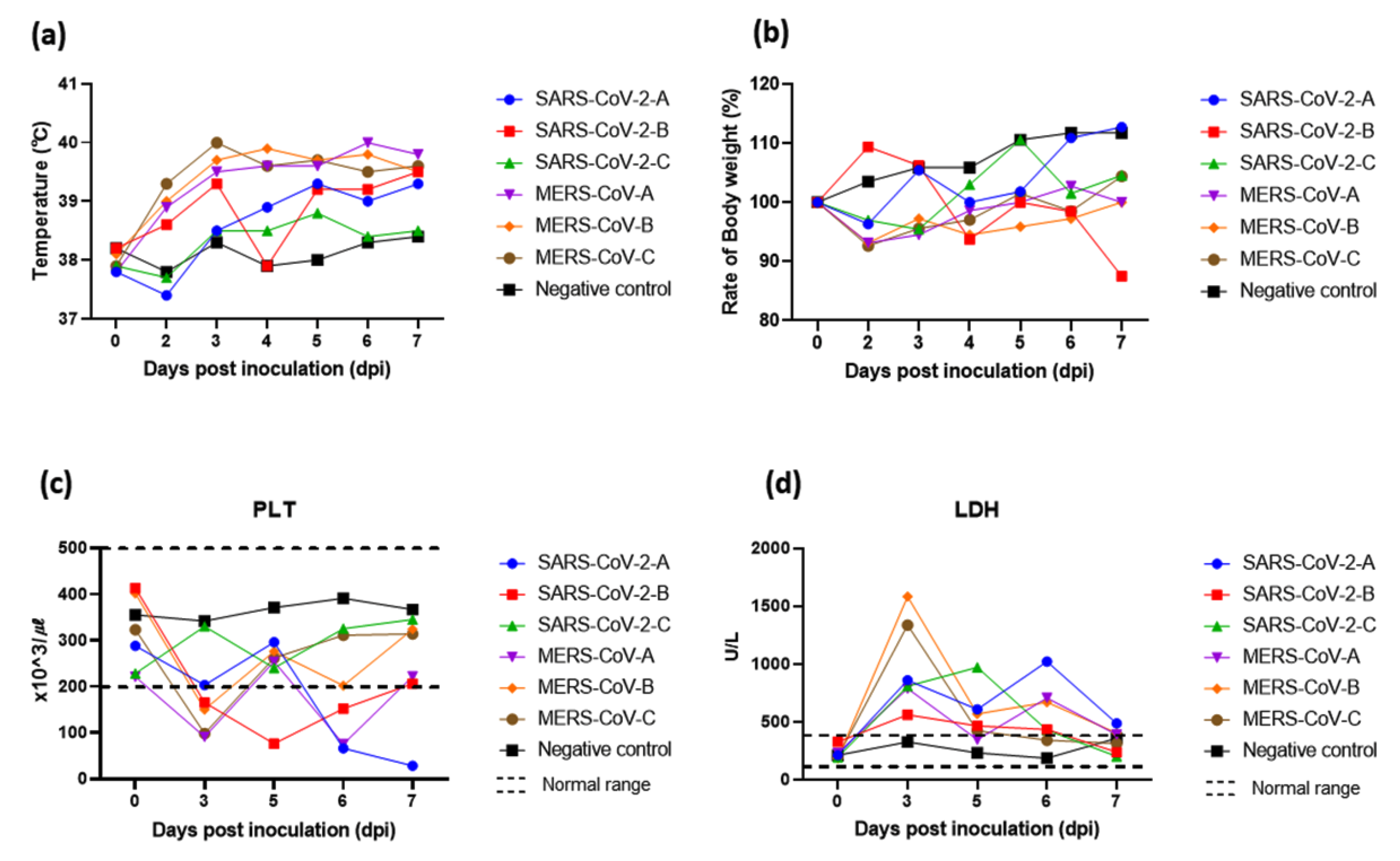
3. Results
3.1. Clinical Observations
3.2. Blood Biochemistry and Hematological Examination
3.3. Viral Loads by Quantitative Real-Time PCR
3.4. Lung Pathology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Sit, T.H.C.; Brackman, C.J.; Ip, S.M.; Tam, K.W.S.; Law, P.Y.T.; To, E.M.W.; Yu, V.Y.T.; Sims, L.D.; Tsang, D.N.C.; Chu, D.K.W.; et al. Infection of dogs with SARS-CoV-2. Nature 2020, 586, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Rabaan, A.A.; Mutair, A.A.; Alawi, Z.A.; Alhumaid, S.; Mohaini, M.A.; Aldali, J.; Tirupathi, R.; Sule, A.A.; Koritala, T.; Adhikari, R.; et al. Comparative pathology, molecular pathogenicity, immunological features, and genetic characterization of three highly pathogenic human coronaviruses (MERS-CoV, SARS-CoV, and SARS-CoV-2). Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 7162–7184. [Google Scholar]
- Memish, Z.A.; Perlman, S.; Van Kerkhove, M.D.; Zumla, A. Middle East respiratory syndrome. Lancet 2020, 395, 1063–1077. [Google Scholar] [CrossRef]
- Wang, N.; Shi, X.; Jiang, L.; Zhang, S.; Wang, D.; Tong, P.; Guo, D.; Fu, L.; Cui, Y.; Liu, X.; et al. Structure of MERS-CoV spike receptor-binding domain complexed with human receptor DPP4. Cell Res. 2013, 23, 986–993. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Munster, V.J. Animal models of Middle East respiratory syndrome coronavirus infection. Antivir. Res. 2015, 122, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Bao, L.; Deng, W.; Xu, L.; Li, F.; Lv, Q.; Yu, P.; Chen, T.; Xu, Y.; Zhu, H.; et al. An animal model of MERS produced by infection of rhesus macaques with MERS coronavirus. J. Infect. Dis. 2014, 209, 236–242. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.S.; Abdelwhab, E.M. Evidence for SARS-CoV-2 Infection of Animal Hosts. Pathogens 2020, 9, 529. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-Coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef]
- Pascucci, I.; Paniccià, M.; Giammarioli, M.; Biagetti, M.; Duranti, A.; Campomori, P.; Smilari, V.; Ancora, M.; Scialabba, S.; Secondini, B.; et al. SARS-CoV-2 Delta VOC in a Paucisymptomatic Dog, Italy. Pathogens 2022, 11, 514. [Google Scholar] [CrossRef]
- Rivero, R.; Garay, E.; Botero, Y.; Serrano-Coll, H.; Gastelbondo, B.; Muñoz, M.; Ballesteros, N.; Castañeda, S.; Patiño, L.H.; Ramirez, J.D.; et al. Human-to-dog transmission of SARS-CoV-2, Colombia. Sci. Rep. 2022, 12, 7880. [Google Scholar] [CrossRef]
- Kim, J.M.; Chung, Y.S.; Jo, H.J.; Lee, N.J.; Kim, M.S.; Woo, S.H.; Park, S.; Kim, J.W.; Kim, H.M.; Han, M.G. Identification of Coronavirus Isolated from a Patient in Korea with COVID-19. Osong Public Health Res. Perspect. 2020, 11, 3–7. [Google Scholar] [CrossRef]
- Noh, J.Y.; Yoon, S.W.; Kim, D.J.; Lee, M.S.; Kim, J.H.; Na, W.; Song, D.; Jeong, D.G.; Kim, H.K. Simultaneous detection of severe acute respiratory syndrome, Middle East respiratory syndrome, and related bat coronaviruses by real-time reverse transcription PCR. Arch. Virol. 2017, 162, 1617–1623. [Google Scholar] [CrossRef]
- Sia, S.F.; Yan, L.M.; Chin, A.W.H.; Fung, K.; Choy, K.T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef]
- Lee, C.Y.; Lowen, A.C. Animal models for SARS-CoV-2. Curr. Opin. Virol. 2021, 48, 73–81. [Google Scholar] [CrossRef]
- Opriessnig, T.; Huang, Y.W. Third update on possible animal sources for human COVID-19. Xenotransplantation 2021, 28, e12671. [Google Scholar] [CrossRef]
- Jurgiel, J.; Filipiak, K.J.; Szarpak, Ł.; Jaguszewski, M.; Smereka, J.; Dzieciątkowski, T. Do pets protect their owners in the COVID-19 era? Med. Hypotheses 2020, 142, e109831. [Google Scholar] [CrossRef]
- Watson, K.M.; Zhang, Y.; Towns, K.; Kahe, K. Owner concerns that pets have COVID-19. Vet. Rec. 2020, 186, 608–609. [Google Scholar] [CrossRef]
- Goryoka, G.W.; Cossaboom, C.M.; Gharpure, R.; Dawson, P.; Tansey, C.; Rossow, J.; Mrotz, V.; Rooney, J.; Torchetti, M.; Loiacono, C.M.; et al. One Health Investigation of SARS-CoV-2 Infection and Seropositivity among Pets in Households with Confirmed Human COVID-19 Cases-Utah and Wisconsin, 2020. Viruses 2021, 13, 1813. [Google Scholar] [CrossRef]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Porter, S.M.; Gordy, P.W.; Nehring, M.; Byas, A.D.; VandeWoude, S.; Ragan, I.K.; Maison, R.M.; Bowen, R.A. Experimental infection of domestic dogs and cats with SARS-CoV-2: Pathogenesis, transmission, and response to reexposure in cats. Proc. Natl. Acad. Sci. USA 2020, 117, 26382–26388. [Google Scholar] [CrossRef]
- Sun, J.; Zhu, A.; Li, H.; Zheng, K.; Zhuang, Z.; Chen, Z.; Shi, Y.; Zhang, Z.; Chen, S.B.; Liu, X.; et al. Isolation of infectious SARS-CoV-2 from urine of a COVID-19 patient. Emerg. Microbes Infect. 2020, 9, 991–993. [Google Scholar] [CrossRef] [PubMed]
- Felsburg, P.J. Overview of immune system development in the dog: Comparison with humans. Hum. Exp. Toxicol. 2002, 21, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Yuki, K.; Fujiogi, M.; Koutsogiannaki, S. COVID-19 pathophysiology: A review. Clin. Immunol. 2020, 215, 108427. [Google Scholar] [CrossRef] [PubMed]
- Fialkowski, A.; Gernez, Y.; Arya, P.; Weinacht, K.G.; Kinane, T.B.; Yonker, L.M. Insight into the pediatric and adult dichotomy of COVID-19: Age-related differences in the immune response to SARS-CoV-2 infection. Pediatr. Pulmonol. 2020, 55, 2556–2564. [Google Scholar] [CrossRef] [PubMed]
- Darif, D.; Hammi, I.; Kihel, A.; El Idrissi Saik, I.; Guessous, F.; Akarid, K. The pro-inflammatory cytokines in COVID-19 pathogenesis: What goes wrong? Microb. Pathog. 2021, 153, 104799. [Google Scholar] [CrossRef]
- Assinger, A. Platelets and infection—An emerging role of platelets in viral infection. Front. Immunol. 2014, 5, 649. [Google Scholar] [CrossRef]
- Henry, B.M.; Aggarwal, G.; Wong, J.; Benoit, S.; Vikse, J.; Plebani, M.; Lippi, G. Lactate dehydrogenase levels predict coronavirus disease 2019 (COVID-19) severity and mortality: A pooled analysis. Am. J. Emerg. Med. 2020, 38, 1722–1726. [Google Scholar] [CrossRef]
- Wu, M.Y.; Yao, L.; Wang, Y.; Zhu, X.Y.; Wang, X.F.; Tang, P.J.; Chen, C. Clinical evaluation of potential usefulness of serum lactate dehydrogenase (LDH) in 2019 novel coronavirus (COVID-19) pneumonia. Respir. Res. 2020, 21, 171. [Google Scholar] [CrossRef]
- Yan, L.; Zhang, H.-T.; Goncalves, J.; Xiao, Y.; Wang, M.; Guo, Y.; Sun, C.; Tang, X.; Jing, L.; Zhang, M. An interpretable mortality prediction model for COVID-19 patients. Nat. Mach. Intel. 2020, 2, 283–288. [Google Scholar] [CrossRef]
- Di Martino, G.; Stefani, A.L.; Lippi, G.; Gagliazzo, L.; McCormick, W.; Gabai, G.; Bonfanti, L. The degree of acceptability of swine blood values at increasing levels of hemolysis evaluated through visual inspection versus automated quantification. J. Vet. Diagn. Investig. 2015, 27, 306–312. [Google Scholar] [CrossRef]
- Friedrichs, K.R.; Harr, K.E.; Freeman, K.P.; Szladovits, B.; Walton, R.M.; Barnhart, K.F.; Blanco-Chavez, J. ASVCP reference interval guidelines: Determination of de novo reference intervals in veterinary species and other related topics. Vet. Clin. Pathol. 2012, 41, 441–453. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Lung | Tonsil | Serum | Nasal | Anal | Urethra | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Days Post-Inoculation | Days Post-Inoculation | Days Post-Inoculation | Days Post-Inoculation | ||||||||||||||||||||
| 0 | 3 | 5 | 6 | 7 | 0 | 3 | 5 | 6 | 7 | 0 | 3 | 5 | 6 | 7 | 0 | 3 | 5 | 6 | 7 | ||||
| SARS- | A | - | - | - | - | - | - | - | - | 2.5 * (+) | 3.4 (+) | 4.1 (+) | 4.5 | - | 3.7 | 4.6 | 4.1 | 4.9 | - | 2.7 | 2.9 | 3.0 | 2.4 |
| CoV-2 | B | - | - | - | - | - | - | - | - | 3.2 (+) | 3.6 | 2.8 (+) | 3.4 | - | 2.2 | 4.3 | 4.5 | 3.7 | - | 1.4 | 1.8 | 2.7 | 2.3 (+) |
| C | - | - | - | - | - | - | - | - | 3.4 (+) | 3.1 (+) | 4.4 | 4.7 | - | 3.5 | 3.9 | 4.0 | 3.4 | - | 2.1 (+) | 3.0 (+) | 2.4 (+) | 1.5 (+) | |
| MERS- | A | - | - | - | - | - | - | - | - | 1.6 | - | - | - | - | 2.3 | - | - | - | - | 1.5 | - | - | - |
| CoV | B | - | - | - | - | - | - | - | - | 2.4 | 2.1 | 2.5 | - | - | 2.1 | - | - | - | - | 1.7 | - | - | - |
| C | - | - | - | - | - | - | - | - | 1.9 | 2.2 | 1.8 | - | - | - | - | - | - | - | - | - | - | - | |
| Cont. | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyoo, K.-S.; Yeo, Y.-H.; Lee, S.-G.; Yeom, M.; Lee, J.-Y.; Kim, K.-C.; Song, D. Susceptibility to SARS-CoV-2 and MERS-CoV in Beagle Dogs. Animals 2023, 13, 624. https://doi.org/10.3390/ani13040624
Lyoo K-S, Yeo Y-H, Lee S-G, Yeom M, Lee J-Y, Kim K-C, Song D. Susceptibility to SARS-CoV-2 and MERS-CoV in Beagle Dogs. Animals. 2023; 13(4):624. https://doi.org/10.3390/ani13040624
Chicago/Turabian StyleLyoo, Kwang-Soo, Yoon-Hwan Yeo, Sung-Geun Lee, Minjoo Yeom, Joo-Yeon Lee, Kyung-Chang Kim, and Daesub Song. 2023. "Susceptibility to SARS-CoV-2 and MERS-CoV in Beagle Dogs" Animals 13, no. 4: 624. https://doi.org/10.3390/ani13040624
APA StyleLyoo, K. -S., Yeo, Y. -H., Lee, S. -G., Yeom, M., Lee, J. -Y., Kim, K. -C., & Song, D. (2023). Susceptibility to SARS-CoV-2 and MERS-CoV in Beagle Dogs. Animals, 13(4), 624. https://doi.org/10.3390/ani13040624