
Black Soldier Fly Larva Fat in Broiler Chicken Diets Affects Breast Meat Quality

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Birds and Housing
2.3. Broiler Diets
2.4. Data and Sample Collection
2.5. Physico-Chemical Analysis
2.6. Statistical Analyses
3. Results
3.1. Hermetia illucens Fatty Acid Profile
3.2. Selected Physico-Chemical Meat Traits
3.3. Growth Performance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kierończyk, B.; Rawski, M.; Mikołajczak, Z.; Homska, N.; Jankowski, J.; Ognik, K.; Józefiak, A.; Mazurkiewicz, J.; Józefiak, D. Available for millions of years but discovered through the last decade: Insects as a source of nutrients and energy in animal diets. Anim. Nutr. 2022, 11, 60–79. [Google Scholar] [CrossRef] [PubMed]
- Gasco, L.; Finke, M.; van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Benzertiha, A.; Kierończyk, B.; Rawski, M.; Mikołajczak, Z.; Urbański, A.; Nogowski, L.; Józefiak, D. Insect fat in animal nutrition: A review. Ann. Anim. Sci. 2020, 20, 1217–1240. [Google Scholar] [CrossRef]
- Kierończyk, B.; Sypniewski, J.; Mikołajczak, Z.; Rawski, M.; Pruszyńska-Oszmałek, E.; Sassek, M.; Kołodziejski, P.; Józefiak, D. Replacement of soybean oil with cold-extracted fat from Hermetia illucens in young turkey diets: Effects on performance, nutrient digestibility, selected organ measurements, meat and liver tissue traits, intestinal microbiota modulation, and physiological. Anim. Feed Sci. Technol. 2022, 286, 115210. [Google Scholar] [CrossRef]
- Kierończyk, B.; Rawski, M.; Józefiak, A.; Mazurkiewicz, J.; Świątkiewicz, S.; Siwek, M.; Bednarczyk, M.; Szumacher-Strabel, M.; Cieślak, A.; Benzertiha, A.; et al. Effects of replacing soybean oil with selected insect fats on broilers. Anim. Feed Sci. Technol. 2018, 240, 170–183. [Google Scholar] [CrossRef]
- Benzertiha, A.; Kierończyk, B.; Rawski, M.; Kołodziejski, P.; Bryszak, M.; Józefiak, D. Insect oil as an alternative to palm oil and poultry fat in broiler chicken nutrition. Animals 2019, 9, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kierończyk, B.; Rawski, M.; Stuper-Szablewska, K.; Józefiak, D. First report of the apparent metabolisable energy value of black soldier fly larvae fat used in broiler chicken diets. Animal 2022, 16, 100656. [Google Scholar] [CrossRef]
- Sauvant, D.; Perez, J.-M.; Tran, G. Tables of Composition and Nutritional Value of Feed Materials: Pigs, Poultry, Cattle, Sheep, Goats, Rabbits, Horses and Fish; Wageningen Academic Publishers: Wageningen, The Netherlands; INRA: Paris, France, 2004; ISBN 9086866689. [Google Scholar]
- Schiavone, A.; Dezzutto, D.; Bergagna, S.; Gai, F.; De Marco, M.; Meneguz, M.; Gasco, L.; Dalle Zotte, A.; Biasato, I.; Cullere, M.; et al. Partial or total replacement of soybean oil by black soldier fly larvae (Hermetia illucens L.) fat in broiler diets: Effect on growth performances, feed-choice, blood traits, carcass characteristics and meat quality. Ital. J. Anim. Sci. 2016, 16, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.; Kim, M.; Jeong, J.Y.; Kim, H.R.; Ji, S.Y.; Jung, H.; Park, S.H. Black soldier fly (Hermetia illucens) larvae oil as an alternative fat ingredient to soybean oil in laying hen diets. Anim. Biosci. 2022, 35, 1408–1417. [Google Scholar] [CrossRef]
- Gasco, L.; Józefiak, A.; Henry, M. Beyond the protein concept: Health aspects of using edible insects on animals. J. Insects Food Feed 2020, 7, 715–741. [Google Scholar] [CrossRef]
- Sypniewski, J.; Kierończyk, B.; Benzertiha, A.; Mikołajczak, Z.; Pruszyńska-Oszmałek, E.; Kołodziejski, P.; Sassek, M.; Rawski, M.; Czekała, W.; Józefiak, D. Replacement of soybean oil by Hermetia illucens fat in turkey nutrition: Effect on performance, digestibility, microbial community, immune and physiological status and final product quality. Br. Poult. Sci. 2020, 61, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Kierończyk, B.; Rawski, M.; Mikołajczak, Z.; Leciejewska, N.; Józefiak, D. Hermetia iilucens fat affects the gastrointestinal tract selected microbial populations, their activity, and the immune status of broiler chickens. Ann. Anim. Sci. 2021, 22, 663–675. [Google Scholar] [CrossRef]
- Kierończyk, B.; Sypniewski, J.; Rawski, M.; Czekała, W.; Swiatkiewicz, S.; Józefiak, D. From waste to sustainable feed material: The effect of Hermetia illucens oil on the growth performance, nutrient digestibility, and gastrointestinal tract morphometry of broiler chickens. Ann. Anim. Sci. 2020, 20, 157–177. [Google Scholar] [CrossRef] [Green Version]
- Riekkinen, K.; Väkeväinen, K.; Korhonen, J. The effect of substrate on the nutrient content and fatty acid composition of edible insects. Insects 2022, 13, 590. [Google Scholar] [CrossRef]
- Hoc, B.; Genva, M.; Fauconnier, M.L.; Lognay, G.; Francis, F.; Caparros Megido, R. About lipid metabolism in Hermetia illucens (L. 1758): On the origin of fatty acids in prepupae. Sci. Rep. 2020, 10, 11916. [Google Scholar] [CrossRef] [PubMed]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Dalle Zotte, A.; Cullere, M.; Martins, C.; Alves, S.P.; Freire, J.P.B.; Falcão-e-Cunha, L.; Bessa, R.J.B. Incorporation of black soldier fly (Hermetia illucens L.) larvae fat or extruded linseed in diets of growing rabbits and their effects on meat quality traits including detailed fatty acid composition. Meat Sci. 2018, 146, 50–58. [Google Scholar] [CrossRef]
- Fawole, F.J.; Labh, S.N.; Hossain, M.S.; Overturf, K.; Small, B.C.; Welker, T.L.; Hardy, R.W.; Kumar, V. Insect (black soldier fly larvae) oil as a potential substitute for fish or soy oil in the fish meal-based diet of juvenile rainbow trout (Oncorhynchus mykiss). Anim. Nutr. 2021, 7, 1360–1370. [Google Scholar] [CrossRef]
- Li, S.; Zhou, J.; Tian, J.; Yu, H.; Zhang, B.; Ji, H. Influence of black soldier fly (Hermetia illucens) larvae oil on growth performance, body composition, tissue fatty acid composition and lipid deposition in juvenile jian carp (Cyprinus carpio var. jian). Aquaculture 2016, 465, 43–52. [Google Scholar] [CrossRef]
- Cullere, M.; Schiavone, A.; Dabbou, S.; Gasco, L.; Zotte, A.D. Meat quality and sensory traits of finisher broiler chickens fed with black soldier fly (Hermetia illucens L.) larvae fat as alternative fat source. Animals 2019, 9, 140. [Google Scholar] [CrossRef] [Green Version]
- Aviagen Ross Broiler Management Handbook. 2018. Available online: https://en.aviagen.com/assets/Tech_Center/Ross_PS/RossPSHandBook2018.pdf (accessed on 15 February 2023).
- CIE. Official recommendations on uniform color space, Color difference equations and metric color terms. CIE Publ. 1976, 15, 1976. [Google Scholar]
- Petracci, M.; Baéza, E. Harmonization of methodologies for the assessment of poultry meat quality features. World’s Poult. Sci. J. 2011, 67, 137–151. [Google Scholar] [CrossRef]
- Bowker, B.; Hawkins, S.; Zhuang, H. Measurement of water-holding capacity in raw and freeze-dried broiler breast meat with visible and near-infrared spectroscopy. Poult. Sci. 2014, 93, 1834–1841. [Google Scholar] [CrossRef]
- Stuper-Szablewska, K.; Buśko, M.; Góral, T.; Perkowski, J. The fatty acid profile in different wheat cultivars depending on the level of contamination with microscopic fungi. Food Chem. 2014, 153, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Cieślak, A.; Kierończyk, B.; Huang, H.; Yanza, Y.R.; Zaworska-Zakrzewska, A.; Józefiak, D.; Szumacher-Strabel, M. Effects of raw and fermented rapeseed cake on growth performance, methane production, and breast meat fatty acid composition in broiler chickens. Animals 2020, 10, 2250. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Bang, H.T.; Kim, K.H.; Kim, M.J.; Jeong, J.Y.; Chun, J.L.; Ji, S.Y. Evaluation of black soldier fly larvae oil as a dietary fat source in broiler chicken diets. J. Anim. Sci. Technol. 2020, 62, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Józefiak, D.; Kierończyk, B.; Rawski, M.; Hejdysz, M.; Rutkowski, A.; Engberg, R.M.; Højberg, O. Clostridium perfringens challenge and dietary fat type affect broiler chicken performance and fermentation in the gastrointestinal tract. Animal 2014, 8, 912–922. [Google Scholar] [CrossRef]
- Sosa, D.A.T.; Fogliano, V. Potential of insect-derived ingredients for food applications. In Insectect Physioliology Ecology; IntechOpen: London, UK, 2017; pp. 215–231. [Google Scholar]
- Nekrasov, R.V.; Ivanov, G.A.; Chabaev, M.G.; Zelenchenkova, A.A.; Bogolyubova, N.V.; Nikanova, D.A.; Sermyagin, A.A.; Bibikov, S.O.; Shapovalov, S.O. Effect of black soldier fy (Hermetia illucens L.) fat on health and productivity performance of dairy cows. Animals 2022, 12, 2118. [Google Scholar] [CrossRef]
- Danieli, P.P.; Lussiana, C.; Gasco, L.; Amici, A.; Ronchi, B. The effects of diet formulation on the yield, proximate composition, and fatty acid profile of the black soldier fly (Hermetia illucens L.) prepupae intended for animal feed. Animals 2019, 9, 178. [Google Scholar] [CrossRef] [Green Version]
- Belghit, I.; Waagbø, R.; Lock, E.-J.; Liland, N.S. Insect-based diets high in lauric acid reduce liver lipids in freshwater atlantic salmon. Aquac. Nutr. 2019, 25, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Borrelli, L.; Varriale, L.; Dipineto, L.; Pace, A.; Menna, L.F.; Fioretti, A. Insect derived lauric acid as promising alternative strategy to antibiotics in the antimicrobial resistance scenario. Front. Microbiol. 2021, 12, 620798. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Li, Q.; Zhang, J.; Yu, Z. Double the biodiesel yield: Rearing black soldier fly larvae, Hermetia illucens, on solid residual fraction of restaurant waste after grease extraction for biodiesel production. Renew. Energy 2012, 41, 75–79. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Gort, G.; Dicke, M.; van Loon, J.J.A. Nutritional plasticity of the black soldier fly (Hermetia illucens) in response to artificial diets varying in protein and carbohydrate concentrations. J. Insects Food Feed 2021, 7, 51–61. [Google Scholar] [CrossRef]
- Palma, L.; Fernandez-Bayo, J.; Niemeier, D.; Pitesky, M.; VanderGheynst, J.S. Managing high fiber food waste for the cultivation of black soldier fly larvae. NPJ Sci. Food 2019, 3, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschirner, M.; Simon, A. Influence of different growing substrates and processing on the nutrient composition of black soldier fly larvae destined for animal feed. J. Insects Food Feed 2015, 1, 249–259. [Google Scholar] [CrossRef]
- Schiavone, A.; Dabbou, S.; Petracci, M.; Zampiga, M.; Sirri, F.; Biasato, I.; Gai, F.; Gasco, L. Black soldier fly defatted meal as a dietary protein source for broiler chickens: Effects on carcass traits, breast meat quality and safety. Animal 2019, 13, 2397–2405. [Google Scholar] [CrossRef] [PubMed]
- Fabrikov, D.; Barroso, F.G.; Sánchez-Muros, M.J.; Hidalgo, M.C.; Cardenete, G.; Tomás-Almenar, C.; Melenchón, F.; Guil-Guerrero, J.L. Effect of feeding with insect meal diet on the fatty Acid compositions of sea bream (Sparus aurata), tench (Tinca tinca) and rainbow trout (Oncorhynchus mykiss) fillets. Aquaculture 2021, 545, 737170. [Google Scholar] [CrossRef]
- Mašek, T.; Starčević, K.; Filipović, N.; Stojević, Z.; Brozić, D.; Gottstein, Ž.; Severin, K. Tissue fatty acid composition and estimated ∆ desaturase activity after castration in chicken broilers fed with linseed or sunflower oil. J. Anim. Physiol. Anim. Nutr. 2014, 98, 384–392. [Google Scholar] [CrossRef]
- Tor, M.; Estany, J.; Francesch, A.; Cubiló, M.D. Comparison of fatty acid profiles of edible meat, adipose tissues and muscles between cocks and capons. Anim. Res. 2005, 54, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Milićević, D.; Vranić, D.; Mašić, Z.; Parunović, N.; Trbović, D.; Nedeljković-Trailović, J.; Petrović, Z. The role of total fats, saturated/unsaturated fatty acids and cholesterol content in chicken meat as cardiovascular risk factors. Lipids Health Dis. 2014, 13, 42. [Google Scholar] [CrossRef] [Green Version]
- Turley, J.; Thompson, J. Nutrition: Your Life Science; Cengage Learning: Bostona, MA, USA, 2015; ISBN 1305480619. [Google Scholar]
- Bovera, F.; Loponte, R.; Marono, S.; Piccolo, G.; Parisi, G.; Iaconisi, V.; Gasco, L.; Nizza, A. Use of Tenebrio molitor larvae meal as protein source in broiler diet: Effect on growth performance, nutrient digestibility, and carcass and meat traits. J. Anim. Sci. 2016, 94, 639–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullere, M.; Tasoniero, G.; Giaccone, V.; Acuti, G.; Marangon, A.; Dalle Zotte, A. Black soldier fly as dietary protein source for broiler quails: Meat proximate composition, fatty acid and amino acid profile, oxidative status and sensory traits. Animal 2018, 12, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Leiber, F.; Gelencsér, T.; Stamer, A.; Amsler, Z.; Wohlfahrt, J.; Früh, B.; Maurer, V. Insect and legume-based protein sources to replace soybean cake in an organic broiler diet: Effects on growth performance and physical meat quality. Renew. Agric. Food Syst. 2017, 32, 21–27. [Google Scholar] [CrossRef]
- Altmann, B.A.; Neumann, C.; Velten, S.; Liebert, F.; Mörlein, D. Meat quality derived from high inclusion of a micro-alga or insect meal as an alternative protein source in poultry diets: A pilot study. Foods 2018, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, E.; Pretorius, Q.; Hoffman, L.C.; Drew, D.W. The carcass quality, meat quality and sensory characteristics of broilers raised on diets containing either Musca domestica larvae meal, fish meal or soya bean meal as the main protein source. Anim. Prod. Sci. 2013, 54, 622–628. [Google Scholar] [CrossRef]
- Pieterse, E.; Erasmus, S.W.; Uushona, T.; Hoffman, L.C. Black soldier fly (Hermetia illucens) pre-pupae meal as a dietary protein source for broiler production ensures a tasty chicken with standard meat quality for every pot. J. Sci. Food Agric. 2019, 99, 893–903. [Google Scholar] [CrossRef]
- Beauclercq, S.; Mignon-Grasteau, S.; Petit, A.; Berger, Q.; Lefèvre, A.; Métayer-Coustard, S.; Tesseraud, S.; Emond, P.; Berri, C.; Le Bihan-Duval, E. A divergent selection on breast meat ultimate pH, a key factor for chicken meat quality, is associated with different circulating lipid profiles. Front. Physiol. 2022, 13, 935868. [Google Scholar] [CrossRef]
- Mir, N.A.; Rafiq, A.; Kumar, F.; Singh, V.; Shukla, V. Determinants of broiler chicken meat quality and factors affecting them: A review. J. Food Sci. Technol. 2017, 54, 2997–3009. [Google Scholar] [CrossRef]
- Bianchi, M.; Ferioli, F.; Petracci, M.; Caboni, M.F.; Cavani, C. The influence of dietary lipid source on quality characteristics of raw and processed chicken meat. Eur. Food Res. Technol. 2009, 229, 339–348. [Google Scholar] [CrossRef]


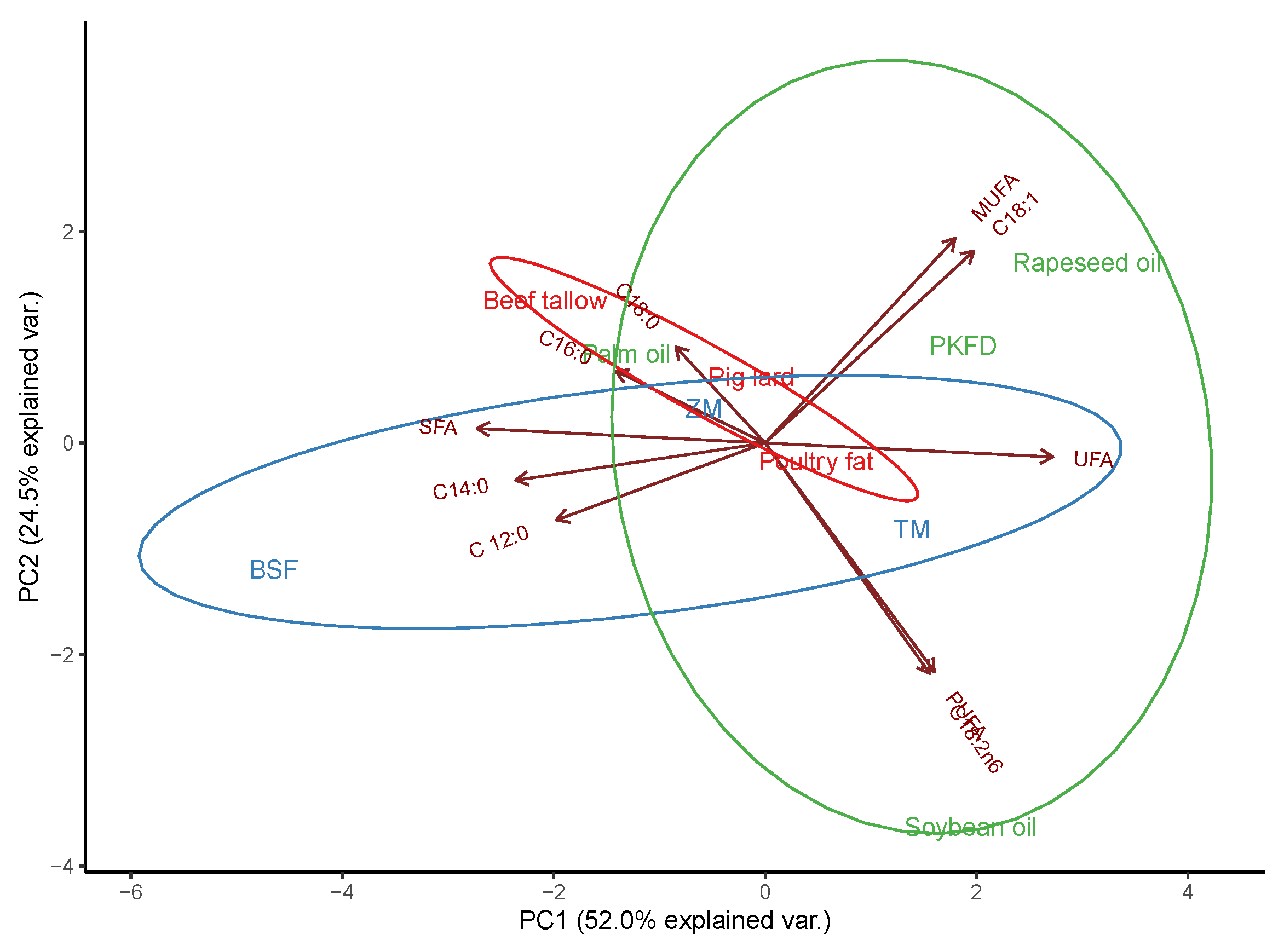{kind=link}
| Ingredients, g kg−1 | 1 HI0 | 2 HI03 | 3 HI06 | 4 HI09 |
|---|---|---|---|---|
| Maize | 657 | 657 | 657 | 657 |
| Soybean meal, 468 g CP/kg | 306 | 306 | 306 | 305 |
| Monocalcium phosphate | 11.9 | 11.9 | 11.9 | 11.9 |
| Limestone | 10.5 | 10.5 | 10.5 | 10.5 |
| Hermetia illucens larvae fat | 0.00 | 30.0 | 60.0 | 90.0 |
| NaCl | 2.60 | 2.60 | 2.60 | 2.60 |
| Na2SO4 | 1.20 | 1.20 | 1.20 | 1.20 |
| 5 Mineral-vitamin premix | 3.00 | 3.00 | 3.00 | 3.00 |
| L-Lysine | 1.80 | 1.80 | 1.80 | 1.80 |
| L-Methionine | 2.30 | 2.30 | 2.30 | 2.30 |
| L-Threonine | 0.60 | 0.60 | 0.60 | 0.60 |
| L-Valine | 0.10 | 0.10 | 0.10 | 0.10 |
| TiO2 | 3.00 | 3.00 | 3.00 | 3.00 |
| Analyzed nutritive value, g kg−1 | ||||
| Dry matter | 884 | 888 | 893 | 896 |
| Gross energy, MJ/kg | 16.4 | 17.0 | 18.0 | 18.5 |
| Crude protein | 208 | 222 | 209 | 207 |
| Crude fat | 2.65 | 4.31 | 8.2 | 10.2 |
| Item | 1 HI | 2 HI0 | 3 HI03 | 4 HI06 | 5 HI09 |
|---|---|---|---|---|---|
| Saturated | |||||
| C8:0 | 0.12 | 0.00 | 0.02 | 0.05 | 0.1 |
| C10:0 | 0.04 | 0.00 | 0.01 | 0.02 | 0.02 |
| C12:0 | 20.3 | 0.00 | 3.86 | 6.1 | 8.4 |
| C14:0 | 6.74 | 0.00 | 1.6 | 2.2 | 3.0 |
| C16:0 | 26.09 | 11.5 | 13.2 | 15.5 | 16.9 |
| C18:0 | 2.46 | 2.80 | 2.60 | 2.70 | 2.68 |
| C21:0 | 0.00 | 0.00 | 0.1 | 0.1 | 0.00 |
| C24:0 | 0.00 | 0.00 | 0.1 | 0.1 | 0.00 |
| Monosaturated | |||||
| C16:1n7 | 24.1 | 0.00 | 1.48 | 1.9 | 2.3 |
| C18:1 n9 | 3.45 | 34.8 | 32.9 | 29.9 | 26.4 |
| Polysaturated | |||||
| C18:2n6 (linoleic acid) | 16.7 | 50.8 | 44.0 | 41.33 | 40.1 |
| C18:3n3 (linolenic acid) | 0.00 | 0.10 | 0.10 | 0.10 | 0.10 |
| Total | 100 | 100 | 100 | 100 | 100 |
| 6 SFA | 55.75 | 14.3 | 21.52 | 26.77 | 31.1 |
| 7 UFA | 44.25 | 85.7 | 78.48 | 73.23 | 68.9 |
| 8 MUFA | 27.55 | 34.8 | 34.38 | 31.8 | 28.7 |
| 9 PUFA | 16.70 | 50.9 | 44.1 | 41.43 | 40.2 |
| PUFA/SFA | 0.30 | 3.56 | 2.05 | 1.55 | 1.29 |
| Item | Treatment | 5 SEM | p | Linear Effect | Quadratic Effect | |||
|---|---|---|---|---|---|---|---|---|
| 1 HI0 | 2 HI03 | 3 HI06 | 4 HI09 | |||||
| Saturated | ||||||||
| C12:0 | 6 ND c | 0.14 b | 0.21 b | 0.30 a | 0.02 | <0.001 | <0.001 | 0.378 |
| C14:0 | 0.54 d | 0.86 c | 0.96 b | 1.08 a | 0.03 | <0.001 | <0.001 | <0.001 |
| C15:0 | ND d | 0.06 c | 0.13 b | 0.24 a | 0.02 | <0.001 | <0.001 | 0.069 |
| C16:0 | 15.3 d | 14.6 c | 14.2 b | 13.9 a | 0.09 | <0.001 | <0.001 | <0.001 |
| C17:0 | 0.28 c | 0.19 b | 0.11 a | 0.09 a | 0.01 | <0.001 | <0.001 | <0.001 |
| C18:0 | 4.73 ab | 4.88 a | 4.71 ab | 4.66 b | 0.03 | 0.026 | 0.082 | 0.033 |
| C20:0 | 0.15 c | 0.21 a | 0.18 ab | 0.16 bc | 0.01 | <0.001 | 0.751 | <0.001 |
| Unsaturated | ||||||||
| C16:1 | ND c | 1.24 b | 1.29 b | 1.35 a | 0.09 | <0.001 | <0.001 | <0.001 |
| C17:1 | ND c | 1.55 a | 1.42 b | 1.50 ab | 0.11 | <0.001 | <0.001 | <0.001 |
| C18:1 | 39.3 a | 37.0 b | 36.1 c | 35.7 c | 0.24 | <0.001 | <0.001 | <0.001 |
| C18:2 | 36.0 b | 36.4 b | 38.1 a | 38.6 a | 0.20 | <0.001 | <0.001 | 0.887 |
| C18:3n6 | 0.18 b | 0.29 a | 0.30 a | 0.39 a | 0.02 | <0.001 | <0.001 | 0.728 |
| C18:3n3 | 2.75 a | 2.39 b | 2.17 c | 1.95 d | 0.05 | <0.001 | <0.001 | 0.178 |
| C20:1 | 0.78 a | 0.09 b | 0.07 bc | 0.05 c | 0.05 | <0.001 | <0.001 | <0.001 |
| C20:2 | ND c | 0.12 a | 0.09 b | 0.08 b | 0.01 | <0.001 | <0.001 | <0.001 |
| Summarized | ||||||||
| 7 SFA | 21.0 a | 20.9 a | 20.5 b | 20.4 b | 0.06 | <0.001 | <0.001 | 0.980 |
| 8 UFA | 79.0 b | 79.1 b | 79.5 a | 79.6 a | 0.06 | <0.001 | <0.001 | 0.980 |
| 9 MUFA | 40.1 a | 39.8 a | 38.9 b | 38.6 b | 0.14 | <0.001 | <0.001 | 0.925 |
| 10 PUFA | 38.9 b | 39.1 b | 40.6 a | 40.9 a | 0.17 | <0.001 | <0.001 | 0.689 |
| PUFA/SFA | 1.85 b | 1.87 b | 1.98 a | 2.01 a | 0.01 | <0.001 | <0.001 | 0.857 |
| n6 | 36.2 b | 36.8 b | 38.5 a | 39.1 a | 0.21 | <0.001 | <0.001 | 0.824 |
| n3 | 2.75 a | 2.69 b | 2.17 c | 1.95 d | 0.05 | <0.001 | <0.001 | 0.178 |
| n6/n3 | 13.2 d | 15.4 c | 18.1 b | 20.1 a | 0.46 | <0.001 | <0.001 | 0.948 |
| Atherogenic index | 0.221 b | 0.230 a | 0.229 a | 0.232 a | <0.01 | <0.001 | <0.001 | 0.037 |
| Thrombogenic index | 0.443 | 0.446 | 0.439 | 0.438 | <0.01 | 0.128 | 0.058 | 0.472 |
| Item | Treatment | 5 SEM | p | Linear Effect | Quadratic Effect | |||
|---|---|---|---|---|---|---|---|---|
| 1 HI0 | 2 HI03 | 3 HI06 | 4 HI09 | |||||
| Color | ||||||||
| 6 L* | 64.1 b | 70.4 a | 71.3 a | 69.8 ab | 0.83 | 0.002 | 0.009 | 0.010 |
| 7 a* | 5.84 a | 6.30 a | 5.86 a | 2.54 b | 0.46 | 0.002 | 0.007 | 0.025 |
| 8 b* | 13.3 b | 20.3 a | 20.6 a | 21.6 a | 0.81 | <0.001 | <0.001 | 0.024 |
| Chroma (C*) | 14.6 b | 21.4 a | 21.5 a | 22.0 a | 0.82 | <0.001 | 0.001 | 0.027 |
| Hue angle (H°) | 67.9 b | 72.8 a | 74.1 a | 82.8 a | 1.23 | <0.001 | <0.001 | 0.305 |
| pH | ||||||||
| 20 min | 6.41 | 6.33 | 6.36 | 6.47 | 0.29 | 0.198 | 0.472 | 0.100 |
| 60 min | 5.86 | 5.98 | 6.00 | 5.97 | 0.03 | 0.131 | 0.124 | 0.185 |
| 24 h | 5.81 | 5.77 | 5.75 | 5.76 | 0.02 | 0.407 | 0.295 | 0.402 |
| Drip loss, % | ||||||||
| 24 h | 3.31 a | 2.80 b | 3.00 ab | 2.73 b | 0.07 | 0.023 | 0.010 | 0.343 |
| 7 days | 5.97 a | 4.71 b | 4.86 b | 4.55 b | 0.13 | <0.001 | <0.001 | 0.035 |
| 9 Salt, % | ||||||||
| 24 h | 15.3 a | 12.9 ab | 10.6 ab | 8.97 b | 0.77 | 0.006 | 0.002 | 0.788 |
| 48 h | 6.39 b | 6.93 ab | 7.88 ab | 9.64 a | 0.43 | 0.012 | 0.004 | 0.438 |
| Item | Treatment | 5 SEM | p | Linear Effect | Quadratic Effect | |||
|---|---|---|---|---|---|---|---|---|
| 1 HI0 | 2 HI03 | 3 HI06 | 4 HI09 | |||||
| 6 BWG, g | ||||||||
| 0–14 days | 325 c | 352 ab | 356 a | 340 b | 2.32 | <0.001 | 0.005 | <0.001 |
| 15–21 days | 344 c | 380 ab | 393 a | 369 b | 2.89 | <0.001 | <0.001 | <0.001 |
| 22–28 days | 432 b | 470 a | 486 a | 486 a | 3.69 | <0.001 | <0.001 | <0.001 |
| 29–35 days | 638 c | 689 ab | 702 a | 673 b | 3.95 | <0.001 | <0.001 | <0.001 |
| 1–35 days | 1739 c | 1891 ab | 1938 a | 1868 b | 10.23 | <0.001 | <0.001 | <0.001 |
| 7 FI, g | ||||||||
| 0–14 days | 447 a | 446 a | 439 a | 417 b | 2.07 | <0.001 | <0.001 | <0.001 |
| 15–21 days | 520 a | 532 a | 528 a | 492 b | 2.91 | <0.001 | <0.001 | <0.001 |
| 22–28 days | 633 b | 662 a | 667 a | 654 a | 2.82 | <0.001 | 0.003 | <0.001 |
| 29–35 days | 1057 bc | 1089 a | 1080 ab | 1051 c | 4.07 | <0.001 | 0.421 | <0.001 |
| 1–35 days | 2656 bc | 2729 a | 2714 ab | 2613 c | 9.62 | <0.001 | 0.054 | <0.001 |
| 8 FCR, g:g | ||||||||
| 0–14 days | 1.38 a | 1.27 b | 1.23 c | 1.23 c | 0.01 | <0.001 | <0.001 | <0.001 |
| 15–21 days | 1.51 a | 1.40 b | 1.34 c | 1.33 c | 0.01 | <0.001 | <0.001 | <0.001 |
| 22–28 days | 1.47 a | 1.41 b | 1.37 bc | 1.35 c | 0.01 | <0.001 | <0.001 | 0.194 |
| 29–35 days | 1.66 a | 1.58 b | 1.54 c | 1.56 ab | 0.01 | <0.001 | <0.001 | <0.001 |
| 1–35 days | 1.53 a | 1.44 b | 1.40 c | 1.40 c | 0.01 | <0.001 | <0.001 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kierończyk, B.; Rawski, M.; Mikołajczak, Z.; Szymkowiak, P.; Stuper-Szablewska, K.; Józefiak, D. Black Soldier Fly Larva Fat in Broiler Chicken Diets Affects Breast Meat Quality. Animals 2023, 13, 1137. https://doi.org/10.3390/ani13071137
Kierończyk B, Rawski M, Mikołajczak Z, Szymkowiak P, Stuper-Szablewska K, Józefiak D. Black Soldier Fly Larva Fat in Broiler Chicken Diets Affects Breast Meat Quality. Animals. 2023; 13(7):1137. https://doi.org/10.3390/ani13071137
Chicago/Turabian StyleKierończyk, Bartosz, Mateusz Rawski, Zuzanna Mikołajczak, Piotr Szymkowiak, Kinga Stuper-Szablewska, and Damian Józefiak. 2023. "Black Soldier Fly Larva Fat in Broiler Chicken Diets Affects Breast Meat Quality" Animals 13, no. 7: 1137. https://doi.org/10.3390/ani13071137
APA StyleKierończyk, B., Rawski, M., Mikołajczak, Z., Szymkowiak, P., Stuper-Szablewska, K., & Józefiak, D. (2023). Black Soldier Fly Larva Fat in Broiler Chicken Diets Affects Breast Meat Quality. Animals, 13(7), 1137. https://doi.org/10.3390/ani13071137