
Transcriptome Profile in Dairy Cows Resistant or Sensitive to Milk Fat Depression

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Sampling and Analysis
3. Results and Discussion
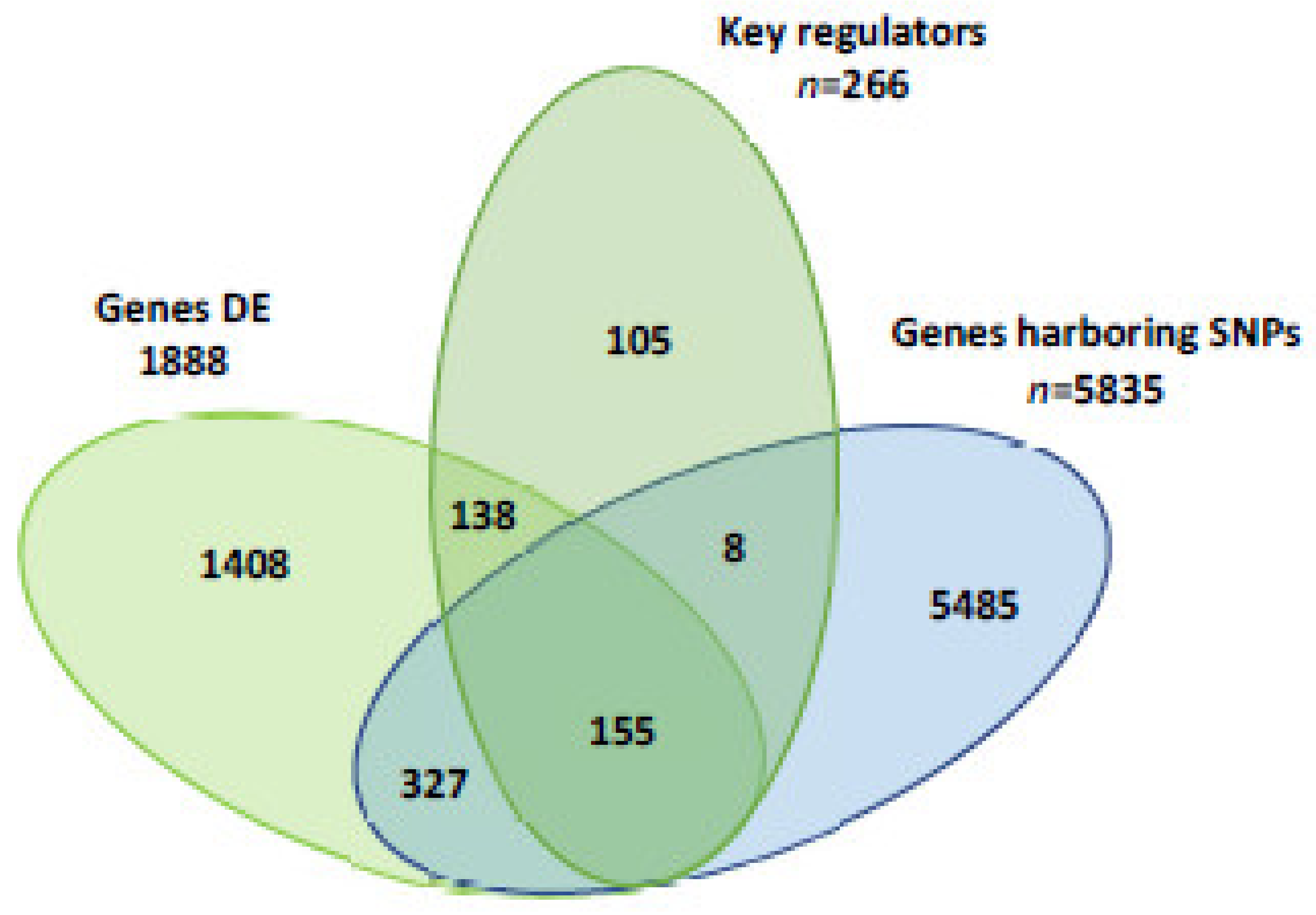
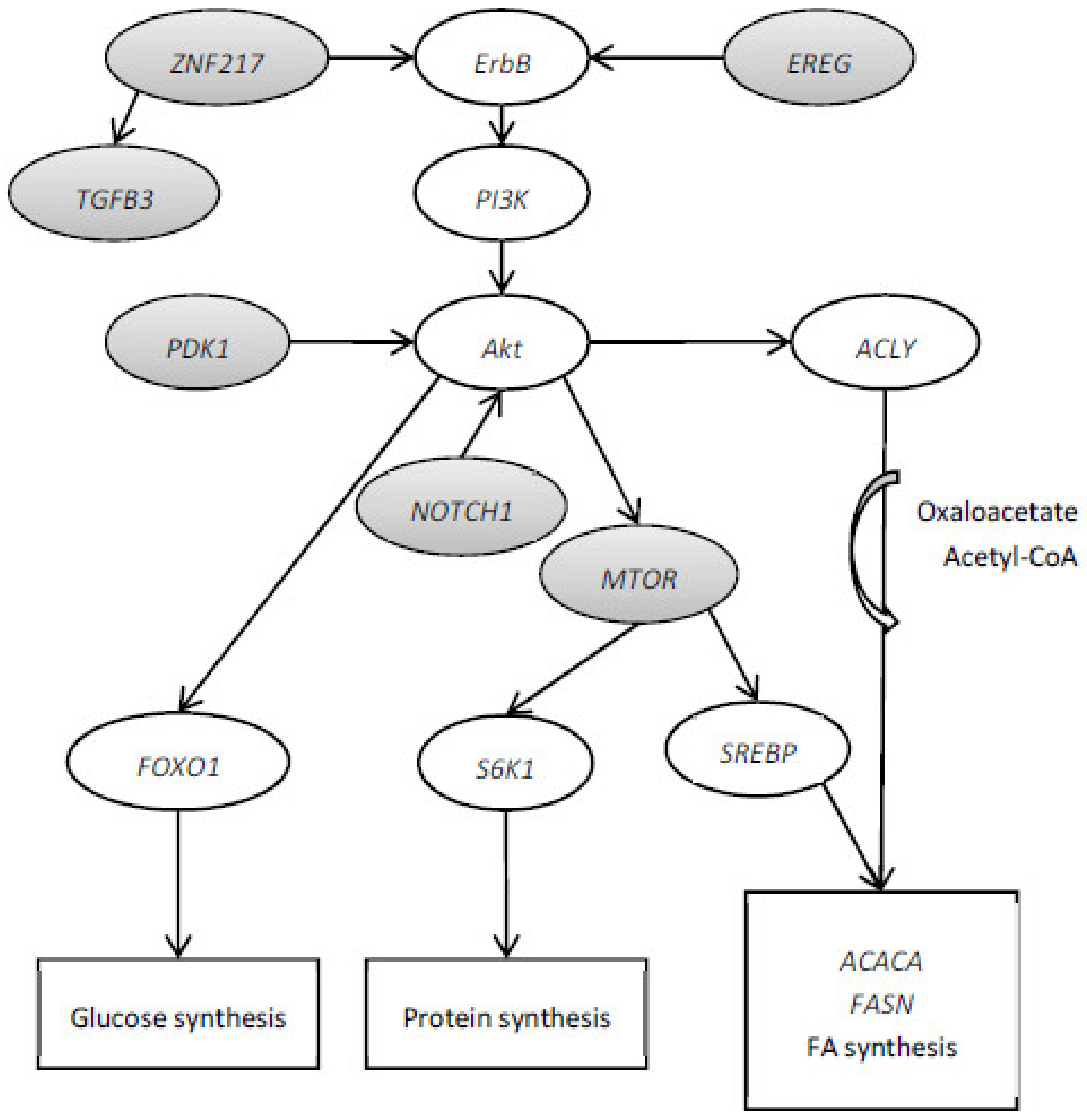
3.1. RNA-Sequencing Expression Analysis, Pathway Analysis and Identification of Key Gene Regulators
3.2. Identification of Genes Harboring SNP
3.3. Identification of Target Genes Differentially Expressed, Key Genes Regulators and Contained SNP
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siurana, A.; Calsamiglia, S. A metaanalysis of feeding strategies to increase the content of conjugated linoleic acid (CLA) in dairy cattle milk and the impact on daily human consumption. Anim. Feed Sci. Technol. 2016, 217, 13–26. [Google Scholar] [CrossRef]
- Bauman, D.E.; Griinari, J.M. Regulation and nutritional manipulation of milk fat: Low-fat milk syndrome. Livest. Prod. Sci. 2001, 70, 15–29. [Google Scholar] [CrossRef]
- Peterson, D.G.; Matitashvili, E.A.; Bauman, D.E. Diet-induced milk fat depression in dairy cows results in increased trans-10, cis-12 CLA in milk fat and coordinated suppression of mRNA abundance for mammary enzymes involved in milk fat synthesis. J. Nutr. 2013, 133, 3098–3102. [Google Scholar] [CrossRef] [Green Version]
- Harvatine, K.J.; Bauman, D.E. SREBP1 and thyroid hormone responsive spot 14 (S14) are involved in the regulation of bovine mammary lipid synthesis during diet-induced milk fat depression and treatment with CLA. J. Nutr. 2006, 136, 2468–2474. [Google Scholar] [CrossRef] [Green Version]
- Kadegowda, A.K.; Bionaz, M.; Piperova, L.S.; Erdman, R.A.; Loor, J.J. Peroxisome proliferator-activated receptor-gamma activation and long-chain fatty acids alter lipogenic gene networks in bovine mammary epithelial cells to various extents. J. Dairy Sci. 2009, 92, 4276–4289. [Google Scholar] [CrossRef] [Green Version]
- McFadden, J.W.; Corl, B.A. Activation of liver X receptor (LXR) enhances de novo fatty acid synthesis in bovine mammary epithelial cells. J. Dairy Sci. 2010, 93, 4651–4658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medrano, J.F.; Rincon, G.; Islas-Trejo, A. Comparative analysis of bovine milk and mammary gland transcriptome using RNA-Seq. In Proceedings of the 9th World Congress on Genetics Applied to Livestock Production, Leipzig, Germany, 1–6 August 2010; Volume 852. [Google Scholar]
- Cánovas, A.; Rincon, G.; Bevilacqua, C.; Islas-Trejo, A.; Brenaut, P.; Hovey, R.C.; Boutinaud, M.; Morgenthaler, C.; VanKlompenberg, M.K.; Martin, P.; et al. Comparison of five different RNA sources to examine the lactating bovine mammary gland transcriptome using RNA- Sequencing. Sci. Rep. 2014, 4, 5297. [Google Scholar] [CrossRef] [Green Version]
- Wickramasinghe, S.; Rincon, G.; Islas-Trejo, A.; Medrano, J.F. Transcriptional profiling of bovine milk using RNA sequencing. BMC Genom. 2012, 13, 45. [Google Scholar] [CrossRef] [Green Version]
- Cánovas, A.; Rincon, G.; Islas-Trejo, A.; Wickramasinghe, S.; Medrano, J.F. SNP discovery in the bovine milk transcriptome using RNA-Seq technology. Mamm. Genome 2010, 21, 592–598. [Google Scholar] [CrossRef] [Green Version]
- Cánovas, A.; Rincón, G.; Islas-Trejo, A.; Jimenez-Flores, R.; Laubscher, A.; Medrano, J.F. RNA sequencing to study gene expression and single nucleotide polymorphism variation associated with citrate content in cow milk. J. Dairy Sci. 2013, 96, 2637–2648. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B. Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinform. 2010, 11, 11–17. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Petea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Lam, S.; Miglior, F.; Fonseca, P.A.S.; Gómez-Redondo, I.; Zeidan, J.; Suárez-Vega, A.; Schenkel, F.; Guan, L.L.; Waters, S.; Stothard, P.; et al. Identification of functional candidate variants and genes for feed efficiency in Holstein and Jersey cattle breeds using RNA-sequencing. J. Dairy Sci. 2021, 104, 1928–1950. [Google Scholar] [CrossRef]
- Asselstine, V.; Miglior, F.; Suárez-Vega, A.; Fonseca, P.A.S.; Mallard, B.; Karrow, N.; Islas-Trejo, A.; Medrano, J.F.; Cánovas, A. Genetic mechanisms regulating the host response during mastitis. J. Dairy Sci. 2019, 102, 9043–9059. [Google Scholar] [CrossRef]
- Cánovas, A.; Pena, R.N.; Gallardo, D.; Ramírez, O.; Amills, M.; Quintanilla, R. Segregation of regulatory polymorphisms with effects on the gluteus medius transcriptome in a purebred pig population. PLoS ONE 2012, 7, e35583. [Google Scholar] [CrossRef] [Green Version]
- Asselstine, V.; Medrano, J.F.; Cánovas, A. Identification of novel alternative splicing associated with mastitis disease in Holstein dairy cows using large gap read mapping. BMC Genom. 2022, 23, 222. [Google Scholar] [CrossRef]
- Lam, S.; Zeidan, J.; Miglior, F.; Suárez-Vega, A.; Gómez-Redondo, I.; Fonseca, P.A.S.; Guan, L.L.; Waters, S.; Cánovas, A. Development and comparison of RNA-sequencing pipelines for more accurate SNP identification: Practical example of functional SNP detection associated with feed efficiency in Nellore beef cattle. BMC Genom. 2020, 21, 703. [Google Scholar] [CrossRef]
- Cui, X.; Hou, Y.; Yang, S.; Xie, Y.; Zhang, S.; Zhang, Y.; Zhang, Q.; Lu, X.; Liu, G.E.; Sun, D. Transcriptional profiling of mammary gland in Holstein cows with extremely different milk protein and fat percentage using RNA sequencing. BMC Genom. 2014, 15, 226. [Google Scholar] [CrossRef] [Green Version]
- Hart, S.N.; Thereneau, T.M.; Zhang, Y.; Poland, G.A.; Kocher, J.P. Calculating sample size estimates for RA sequencing data. J. Comput. Biol. 2013, 20, 970–978. [Google Scholar] [CrossRef] [Green Version]
- McCulloch, C.E. Maximum likelihood algorithms for generalized linear mixed models. J. Am. Stat. Assoc. 1997, 92, 162–170. [Google Scholar] [CrossRef]
- Aban, I.B.; Cutter, G.R.; Mavinga, N. Inferences and power analysis concerning two negative binomial distributions with an application to MRI lesion count data. Comput. Stat. Data Anal. 2008, 53, 820–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, T.V.; Jimenez, C.R. An accurate paired sample test for count data. Bioinformatics 2012, 28, i596–i602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ching, T.; Huang, S.; Garmire, K.X. Power analysis and sample size estimation for RNA-Seq differential expression. RNA 2014, 20, 1684–1696. [Google Scholar] [CrossRef] [Green Version]
- Poplawski, A.; Binder, H. Feasibility of sample size calculation for RNA-seq studies. Brief. Bioinform. 2017, 19, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Rapaport, F.; Khanin, R.; Liang, Y.; Pirun, M.; Krek, A.; Zumbo, P.; Mason, C.E.; Socci, N.D.; Betel, D. Comprehensive evaluation of differential gene expression analysis methods for RNA-seq data. Genome Biol. 2013, 14, R95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yecies, J.L.; Zhang, H.H.; Menon, S.; Liu, S.; Yecies, D.; Lipovsky, A.I.; Gorgun, C.; Kwiatkowski, D.J.; Hotamisligil, G.S.; Lee, C.H.; et al. Akt stimulates hepatic SREBP1c and lipogenesis through parallel mTORC1-dependent and independent pathways. Cell Metab. 2011, 14, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Brown, M.S.; Goldstein, J.L. Bifurcation of insulin signaling pathway in rat liver: mTORC1 required for stimulation of lipogenesis, but not inhibition of gluconeogenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 3441–3446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Zhao, F.; Wei, C.; Liang, M.; Zhang, N.; Wang, C.; Li, Q.Z.; Gao, X.J. Function of SREBP1 in the milk fat synthesis of dairy cow mammary epithelial cells. Int. J. Mol. Sci. 2014, 15, 16998–17013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BCM Genom. 2008, 9, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porstmann, T.; Santos, C.R.; Griffiths, B.; Cully, M.; Wu, M.; Leevers, S.; Griffiths, J.F.; Chung, Y.L.; Schulze, A. SREBP activity is regulated by mTORC1 and contributes to Akt-dependent cell growth. Cell Metab. 2008, 8, 224–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarbassov, D.D.; Ali, S.M.; Sabatini, D.M. Growing roles for the mTOR pathway. Curr. Opin. Cell Biol. 2005, 17, 596–603. [Google Scholar] [CrossRef]
- Riese, D.J.; Cullum, R.L. Epiregulin: Roles in normal physiology and cancer. Semin. Cell Dev. Biol. 2014, 28, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; He, H.W.; Sun, H.X.; Ren, K.H.; Shao, R.G. Dual knockdown of N-ras and epiregulin synergistically suppressed the growth of human hepatoma cells. Biochem. Biophys. Res. Commun. 2009, 387, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Pajvani, U.B.; Qiang, L.; Kangsamaksin, T.; Kitajewski, J.; Ginsberg, H.N.; Accili, D. Inhibition of Notch uncouples Akt activation from hepatic lipid accumulation by decreasing mTorc1 stability. Nat. Med. 2013, 19, 1054–1060. [Google Scholar] [CrossRef] [Green Version]
- Mullen, M.P.; Berry, D.P.; Howard, D.J.; Diskin, M.G.; Lynch, C.O.; Giblin, L.; Kenny, D.A.; Magee, D.A.; Meade, K.G.; Waters, S.M. Single nucleotide polymorphisms in the Insulin-like Growth Factor 1 (IGF-1) gene are associated with performance in Holstein-Friesian dairy cattle. Front. Genet. 2011, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Krig, S.R.; Miller, J.K.; Frietze, S.; Beckett, L.A.; Neve, R.M.; Farnham, P.J.; Yaswen, P.I.; Sweeney, C.A. ZNF217, a candidate breast cancer oncogene amplified at 20q13, regulates expression of the ErbB3 receptor tyrosine kinase in breast cancer cells. Oncogene 2010, 29, 5500–5510. [Google Scholar] [CrossRef] [Green Version]
- Vendrell, J.A.; Thollet, A.; Nguyen, N.T.; Ghayad, S.E.; Vinot, S.; Bieche, I.; Grisard, E.; Josserand, V.; Coll, J.L.; Roux, P.; et al. ZNF217 is a marker of poor prognosis in breast cancer that drives epithelial-mesenchymal transition and invasion. Cancer Res. 2012, 72, 3593–3606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolgacheva, L.P.; Turovskaya, M.V.; Dynnik, V.V.; Zinchenko, V.P.; Goncharov, N.V.; Davletov, B.; Turovsky, E.A. Angiotensin II activates different calcium signaling pathways in adipocytes. Arch. Biochem. Biophys. 2016, 593, 38–49. [Google Scholar] [CrossRef]
- Johansson, J.U.; Woodling, N.S.; Wang, Q.; Panchal, M.; Liang, X.; Trueba-Saiz, A.; Brown, H.D.; Mhatre, S.D.; Loui, T.; Andreasson, K.I. Prostaglandin signaling suppresses beneficial microglial function in Alzheimer’s disease models. J. Clin. Investig. 2015, 125, 350–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossiter, H.; Barresi, C.; Ghannadan, M.; Gruber, F.; Mildner, M.; Födinger, D.; Tschachler, E. Inactivation of VEGF in mammary gland epithelium severely compromises mammary gland development and function. FASEB J. 2016, 21, 3994–4004. [Google Scholar] [CrossRef] [PubMed]
- Adini, A.; Adini, I.; Ghosh, K.; Benny, O.; Pravda, E.; Hu, R.; Luyindula, D.; D’Amato, R.J. The stem cell marker prominin-1/CD133 interacts with vascular endothelial growth factor and potentiates its action. Angiogenesis 2013, 16, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Ibeagha-Awemu, E.M.; Peters, S.O.; Akwanji, K.A.; Imumorin, I.G.; Zhao, X. High density genome wide genotyping-by-sequencing and association identifies common and low frequency SNPs, and novel candidate genes influencing cow milk traits. Sci. Rep. 2016, 6, 31109. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Cow 1 | Gene | p-Value | q-Value | FC 2 | Chr. 3 | Chr. Position | SNP ID | Allele Change |
|---|---|---|---|---|---|---|---|---|
| S | APBB1 | 0.01 | 0.04 | −3.36 | 15 | 45561080 | rs207812421 | C/T |
| S | APBB1 | 0.01 | 0.04 | −3.36 | 15 | 45561104 | rs209491693 | T/C |
| S | APBB1 | 0.01 | 0.04 | −3.36 | 15 | 45569813 | rs210165374 | G/C |
| S | APBB1 | 0.01 | 0.04 | −3.36 | 15 | 45569330 | rs210595896 | C/T |
| S | APBB1 | 0.01 | 0.04 | −3.36 | 15 | 45569819 | rs211146069 | T/C |
| S | APBB1 | 0.01 | 0.04 | −3.36 | 15 | 45576807 | rs41255144 | C/T |
| S | CD38 | 0.01 | 0.02 | −2.31 | 6 | 115589806 | rs109641719 | C/T |
| S | CD38 | 0.01 | 0.02 | −2.31 | 6 | 115591914 | rs134955750 | G/A |
| R | CD38 | 0.01 | 0.02 | −2.31 | 6 | 115595101 | rs136147162 | G/C |
| S | CD38 | 0.01 | 0.02 | −2.31 | 6 | 115592674 | rs43434904 | C/T |
| S | CD38 | 0.01 | 0.02 | −2.31 | 6 | 115589816 | rs43434912 | G/A |
| S | EREG | 0.01 | 0.01 | −9.50 | 6 | 92346668 | rs42580620 | G/A |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31554406 | rs109247749 | A/G |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31489476 | rs111027111 | C/T |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31483700 | rs133983660 | A/G |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31485999 | rs136560138 | G/C |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31528552 | rs137508649 | A/G |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31484135 | rs207631114 | G/C |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31483177 | rs209090694 | T/C |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31483898 | rs209547908 | A/G |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31483715 | rs210883339 | G/C |
| R | FLT1 | 0.01 | 0.01 | 3.55 | 12 | 31483473 | rs211512991 | T/C |
| S | ITGB4 | 0.01 | 0.01 | −4.12 | 19 | 57146058 | rs41926899 | G/A |
| S | ITGB4 | 0.01 | 0.01 | −4.12 | 19 | 57157390 | rs41927658 | C/G |
| S | MTOR | 0.01 | 0.02 | −2.18 | 16 | 39224720 | rs208757293 | T/A |
| S | MTOR | 0.01 | 0.02 | −2.18 | 16 | 39235739 | rs211448695 | G/A |
| S | MTOR | 0.01 | 0.02 | −2.18 | 16 | 39231288 | rs211677647 | C/T |
| S | NFATC2 | 0.01 | 0.01 | −6.51 | 13 | 80208242 | rs137043317 | T/C |
| S | NOTCH1 | 0.01 | 0.02 | −2.47 | 11 | 107708189 | rs110163085 | C/G |
| R | NOTCH1 | 0.01 | 0.02 | −2.47 | 11 | 107686385 | rs133307736 | A/G |
| S | NOTCH1 | 0.01 | 0.02 | −2.47 | 11 | 107685263 | rs207760072 | A/G |
| S | NOTCH1 | 0.01 | 0.02 | −2.47 | 11 | 107698629 | rs211580903 | G/A |
| S | NOTCH1 | 0.01 | 0.02 | −2.47 | 11 | 107679366 | rs378232535 | C/T |
| R | NOTCH2 | 0.01 | 0.03 | −2.51 | 3 | 25061073 | rs135438495 | T/C |
| S | PDPK1 | 0.01 | 0.02 | −3.58 | 25 | 2633350 | rs208965123 | T/C |
| S | PROM1 | 0.01 | 0.01 | −17.9 | 6 | 115755844 | rs110069470 | A/G |
| S | PROM1 | 0.01 | 0.01 | −17.9 | 6 | 115732932 | rs42165955 | A/C |
| S | RICTOR | 0.01 | 0.01 | −2.90 | 20 | 37609145 | rs41940570 | T/C |
| S | RICTOR | 0.01 | 0.01 | −2.90 | 20 | 37610024 | rs41940571 | A/G |
| R | TGFBR3 | 0.01 | 0.01 | −3.90 | 3 | 54795884 | rs110491344 | A/G |
| R | TGFBR3 | 0.01 | 0.01 | −3.90 | 3 | 54796526 | rs134330950 | T/C |
| R | TGFBR3 | 0.01 | 0.01 | −3.90 | 3 | 54798328 | rs379514543 | A/C |
| R | WWC1 | 0.01 | 0.01 | −7.19 | 7 | 82262781 | rs108980081 | C/T |
| S | ZNF217 | 0.01 | 0.05 | −2.06 | 13 | 82061022 | rs134599263 | A/G |
| Comparisons (FC) 1 | ||||
|---|---|---|---|---|
| Gene | 1 | 2 | 3 | 4 |
| APBB1 | −3.36 | −6.42 | - 2 | - |
| CD38 | −2.31 | - | - | - |
| EREG | −9.50 | −3.13 | - | - |
| FLT1 | 3.55 | 5.95 | - | - |
| ITGB4 | −4.12 | −4.97 | - | - |
| MTOR | −2.17 | RG 3 1.025 | - | GDE 4 −2.40 |
| NFATC2 | −6.51 | RG −2.17 | - | GDE −3.58 |
| NOTCH1 | −2.47 | −3.52 | - | - |
| NOTCH2 | −2.51 | - | - | - |
| PDPK1 | −3.58 | - | - | GDE −3.88 |
| PROM1 | −17.89 | GDE −12.72 | - | - |
| RICTOR | −2.90 | GDE −2.47 | - | - |
| TGFBR3 | −3.90 | GDE −2.36 | - | - |
| WWC1 | −7.19 | GDE −6.74 | - | - |
| ZNF217 | −2.06 | −2.77 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siurana, A.; Cánovas, A.; Casellas, J.; Calsamiglia, S. Transcriptome Profile in Dairy Cows Resistant or Sensitive to Milk Fat Depression. Animals 2023, 13, 1199. https://doi.org/10.3390/ani13071199
Siurana A, Cánovas A, Casellas J, Calsamiglia S. Transcriptome Profile in Dairy Cows Resistant or Sensitive to Milk Fat Depression. Animals. 2023; 13(7):1199. https://doi.org/10.3390/ani13071199
Chicago/Turabian StyleSiurana, Adriana, Angela Cánovas, Joaquim Casellas, and Sergio Calsamiglia. 2023. "Transcriptome Profile in Dairy Cows Resistant or Sensitive to Milk Fat Depression" Animals 13, no. 7: 1199. https://doi.org/10.3390/ani13071199
APA StyleSiurana, A., Cánovas, A., Casellas, J., & Calsamiglia, S. (2023). Transcriptome Profile in Dairy Cows Resistant or Sensitive to Milk Fat Depression. Animals, 13(7), 1199. https://doi.org/10.3390/ani13071199