
The Significance of a Multilocus Analysis for Assessing the Biodiversity of the Romanov Sheep Breed in a Comparative Aspect
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
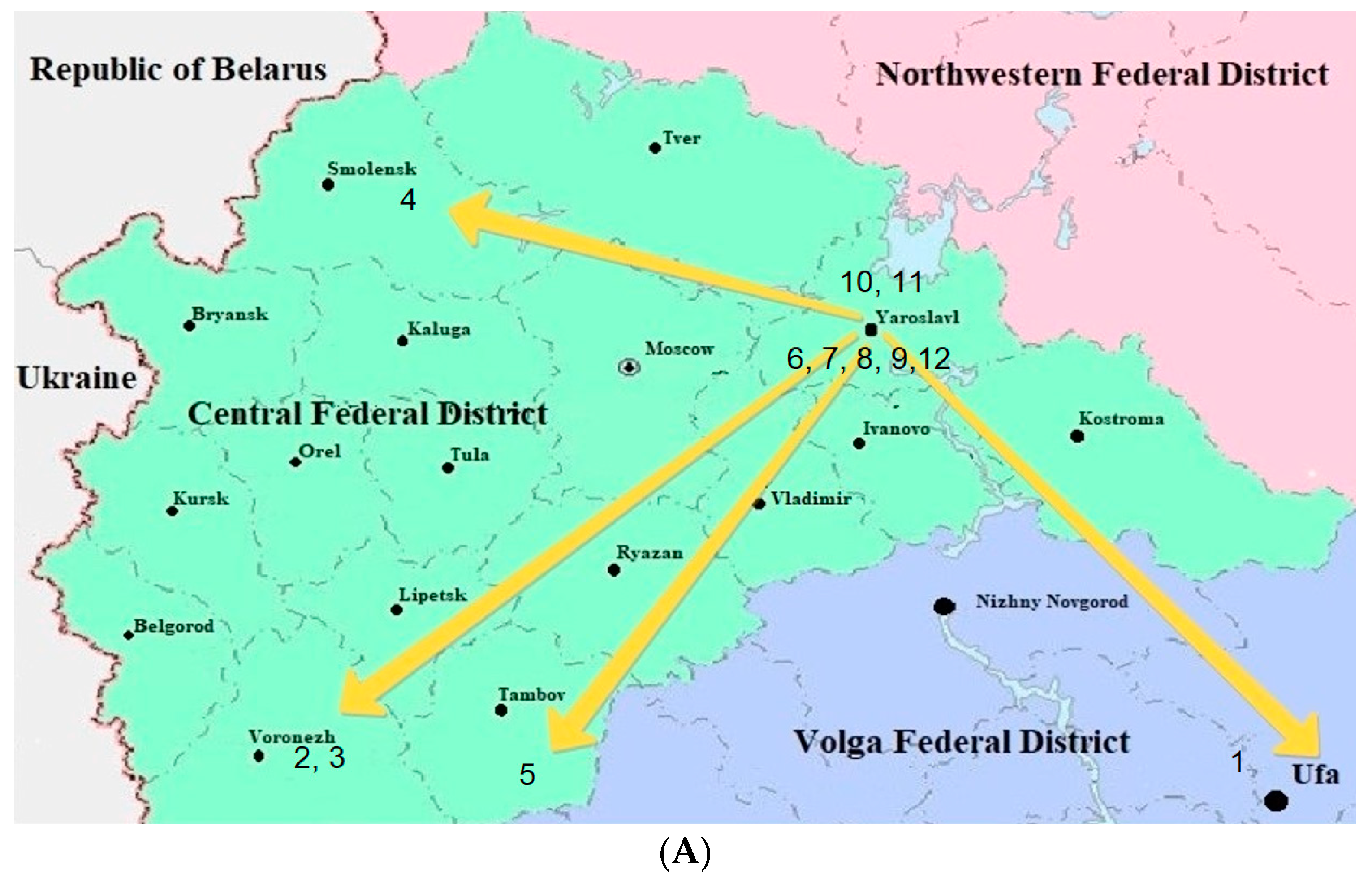
2. Materials and Methods
3. Results and Discussion
3.1. Characteristics of the Romanov Breed According to Seven Systems of Blood Groups
3.2. Diagnosis of High Ovulation Rate Genes in Sheep of the Romanov Breed
3.3. Characteristics of the b Hemoglobin (HB) Locus in Sheep of Various Breeds
3.4. Characterization of the Transferrin (TF) Locus in Sheep of Various Breeds
3.5. Genetic Structure of the Albumin (ALB) Locus in Sheep of Different Breeds
3.6. Characterization of the Prealbumin (PRE) Locus in Sheep of Different Breeds
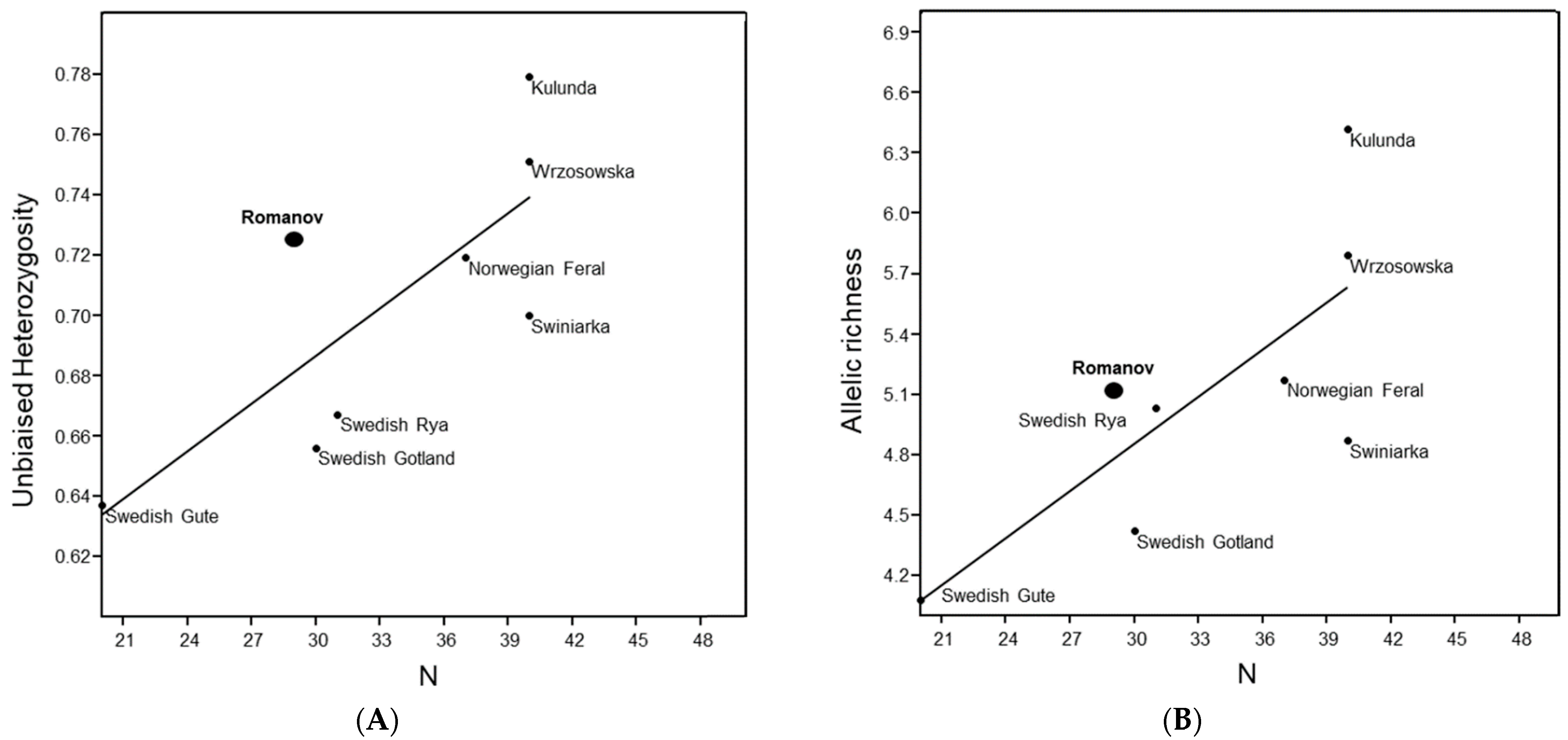
3.7. Comparative Genetic Diversity
3.8. Inter-Population Analysis from Seven Blood Group Systems
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Erokhin, A.I.; Karasev, E.A.; Erokhin, S.A. Romanov Breed of Sheep: State, Improvement, Use of the Gene Pool; Federal State Rosinformagroteh: Moscow, Russia, 2005; p. 329. [Google Scholar]
- Korenev, M.M.; Furaeva, N.S.; Khrustaleva, V.I.; Sokolova, S.I.; Grigoryan, L.N.; Marzanov, N.S. Valuable world gene pool of sheep—Romanov breed. Sheep Goats Wool. Bus. 2017, 3, 2–4. [Google Scholar]
- Ryder, M.L. A survey of European primitive breeds of sheep. Ann. Gén. Sél. Anim. 1981, 13, 381–418. [Google Scholar] [CrossRef]
- Egorov, E.A. Genetic Systems of Sheep Blood Proteins; Publishing House “Fan”: Tashkent, Uzbekistan, 1973; p. 223. [Google Scholar]
- Marzanov, N.S. Immunology and Immunogenetics of Sheep and Goats; Shtiintsa: Chisinau, Moldova, 1991; p. 238. [Google Scholar]
- Marzanov, N.S.; Nasibov, M.G.; Ozerov, M.Y.; Kantanen, J. Allelofond of Various Sheep Breeds by Microsatellites; Publishing House: Dubrovitsy, Russia, 2004; p. 119. [Google Scholar]
- Lyutskanov, P.I. Creation of New Types of Tsigai and Karakul Sheep in the Republic of Moldova Using Genetic Markers; Diss. Doct. Biol. Scie.: Dubrovitsy, Russia, 2009; p. 220. [Google Scholar]
- Tapio, M.; Ozerov, M.; Tapio, I.; Toro, M.A.; Marzanov, N.; Cinkulov, M.; Goncharenko, G.; Kiselyova, T.; Murawski, M.; Kantanen, J. Microsatellite-based genetic diversity and population structure of domestic sheep in northern Eurasia. BMC Genet. 2010, 11, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzanov, N.S.; Komkova, E.A.; Malyuchenko, O.P.; Alekseev, Y.I.; Ozerov, M.Y.; Kantanen, Y.; Lobkov, V.Y.; Marzanova, L.K.; Astafieva, E.E.; Petrov, S.N.; et al. Characteristics of the allele pool of the Romanov breed of sheep according to various types of genetic markers. Probl. Biol. Prod. Anim. 2015, 2, 23–40. (In Russian) [Google Scholar]
- Bernstein, F. Ergebnisse Einer Biostatistischen Zusam-Menfassenden Betrachtung über die Erblichen Blutstrukturen des Menschen. Klin. Wschr. 1924, 3, 1495–1497. [Google Scholar] [CrossRef]
- Polley, S.; De, S.; Brahma, B.; Mukherjee, A.; Vinesh, P.V.; Batabyal, S.; Arora, J.S.; Pan, S.; Samanta, A.K.; Datta, T.K.; et al. Polymorphism of BMPR1B, BMP15 and GDF9 fecundity genes in prolific Garole sheep. Trop. Anim. Health Prod. 2010, 42, 985–993. [Google Scholar] [CrossRef]
- Malyuchenko, O.P.; Alekseev, Y.I.; Monakhova, Y.A.; Marzanova, S.N.; Marzanov, N.S. Study of molecular variability of BMP-15 and GDF9 fertility genes in the Romanov breed of sheep. Izvestiya TAA 2011, 6, 167–169. [Google Scholar]
- Niu, Z.G.; Qin, J.; Jiang, Y.; Ding, X.D.; Ding, Y.G.; Tang, S.; Shi, H.C. The identification of mutation in BMP15 gene associated with litter size in Xinjiang Cele black sheep. Animals 2021, 11, 668. [Google Scholar] [CrossRef] [PubMed]
- Marzanov, N.S.; Maluchenko, O.P.; Koreckaya, E.A.; Marzanova, S.N.; Marzanova, L.K.; Timoshenko, Y.I.; Feizullaev, F.R. Characteristics of the Romanov Breed by the BMP-15 Locus Responsible for Prolificacy in Sheep. Russ. Agric. Sci. 2019, 45, 374–377. [Google Scholar] [CrossRef]
- Petrov, S.N. The Use of Genetic Technology to Characterize the Biological Features and Origin of Sheep Breeds; Diss. Cand. Biol. Sciences: Dubrovitsy, Russia, 2008; p. 126. [Google Scholar]
- Cavalli-Sforza, L.L.; Edwards, A.W. Phylogenetic Analysis Models and Estimation Procedures. Am. J. Hum. Genet. 1967, 19, 233–257. [Google Scholar] [PubMed]
- Zhivotovsky, L.A. Population Biometrics; Nauka: Moscow, Russia, 1991; p. 271. [Google Scholar]
- Glantz, S. Medico-Biological Statistics; Translation from English; Publishing House “Practice”: Moscow, Russia, 1999; p. 459. ISBN 5-89816-009-4. [Google Scholar]
- Iovenko, V.N. Genetic diversity of protein markers in sheep populations from Ukraine. Russ. J. Genet. 2002, 38, 1417–1423. [Google Scholar] [CrossRef]
- Rendel, J. Further studies on some antigenic characters of sheep blood determined by epistatic action of genes. Acta Agric. Scand. 1957, 7, 224–259. [Google Scholar] [CrossRef]
- Nasibov, M.G.; Marzanova, L.K.; Kanatbaev, S.G.; Chmirkov, E.V.; Marzanov, N.S. Identification of antigens and systems of blood groups in various animal species. Agric. Biol. 2005, 6, 119–125. [Google Scholar]
- Davis, G.H. Major genes affecting ovulation rate in sheep. Genet. Sel. Evol. 2005, 37, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Piper, L.R.; Bindon, B.M.; Davis, G.H. The single gene inheritance of the high litter size of the Booroola Merino. In Genetics of Reproduction in Sheep; Land, R.B., Robinson, D.W., Eds.; Butterworths: London, UK, 1985; pp. 115–125. [Google Scholar]
- Chu, M.X.; Liu, Z.H.; Jiao, C.L.; He, Y.Q.; Fang, L.; Ye, S.C.; Chen, G.H.; Wang, J.Y. Mutations in BMPR-IB and BMP-15 genes are associated with litter size in Small Tailed Han sheep (Ovis aries). J. Anim. Sci. 2007, 85, 598–603. [Google Scholar] [CrossRef]
- Zhou, M.; Pan, Z.; Cao, X.; Guo, X.; He, X.; Sun, Q.; Di, R.; Hu, W.; Wang, X.; Zhang, X.; et al. Single Nucleotide Polymorphisms in the HIRA Gene Affect Litter Size in Small Tail Han Sheep. Animals 2018, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Guan, F.; Liu, S.R.; Shi, G.Q.; Yang, L.G. Polymorphism of FecB gene in nine sheep breeds or strains and its effects on litter size, lamb growth and development. Anim. Reprod. Sci. 2007, 99, 44–52. [Google Scholar] [CrossRef]
- Vavilov, N.I. The Origin, Variation, Immunity and Breeding of Cultivated Plants; Chester, K.S., Translator; Chronica Botanica Company: Waltham, MA, USA, 1951; p. 364. [Google Scholar]
- Mullen, M.P.; Hanrahan, J.P.; Howard, D.J.; Powell, R. Investigation of Prolific Sheep from UK and Ireland for Evidence on Origin of the Mutations in BMP15 (FecXG, FecXB) and GDF9 (FecGH) in Belclare and Cambridge Sheep. PLoS ONE 2020, 8, e53172. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Wang, X.; Kijas, J.W.; Dalrymple, B.P. Beta-globin gene evolution in the ruminants: Evidence for an ancient origin of sheep haplotype B. Anim. Genet. 2015, 46, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamenek, V.M. The Role of Biochemical Polymorphism in the Ecological Genetic Differentiation of Animals; Diss. Doct. Biol. Scie.: St. Petersburg-Pushkin, Russia, 1996; p. 272. [Google Scholar]
- Zaitsev, S.Y.; Konopatov, Y.V. Biochemistry of Animals; Publishing House “Lan”: St. Petersburg, Russia, 2004; p. 384. [Google Scholar]
- Ozerov, M.Y.; Tapio, M.; Kantanen, J.; Marzanova, S.N.; Koreckaya, E.A.; Lushnikov, V.P.; Marzanov, N.S. Environmental factors affecting genetic variation in coarse-wool sheep. Russ. Agric. Sci. 2020, 46, 65–70. [Google Scholar] [CrossRef]
- Plakhov, A. Note on the feeding and improvement of sheep. In Proceedings of the Free Economic Society; Publishing House at the Imperial Academy of Sciences: St. Petersburg, Russia, 1802; p. 115. [Google Scholar]
- Ben Jemaa, S.; Ruesche, J.; Sarry, J.; Woloszyn, F.; Lassoued, N.; Fabre, S. The high prolificacy of D’man sheep is associated with the segregation of the FecLL mutation 2 in the B4GALNT2 gene. Reprod. Domest. Anim. 2019, 54, 531–537. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| System | Genotype | Frequency Occurrence | Allele | Frequency Occurrence | Phenotypic Classes | Frequency Occurrence |
|---|---|---|---|---|---|---|
| A (n = 52) | A-/- | 0.3654 | A- | 0.5884 | Aa | 0.1346 |
| Ab/b | 0.0798 | Ab | 0.2899 | Aab | 0.0962 | |
| Aa/a | 0.0126 | Aa | 0.1217 | Ab | 0.4038 | |
| Aa/b | 0.0962 | A- | 0.3654 | |||
| Ab/- | 0.324 | |||||
| Aa/- | 0.1220 | |||||
| C (n = 52) | C-/- | 0.3462 | C- | 0.5827 | C- | 0.3462 |
| Cb/b | 0.0918 | Cb | 0.3055 | Cb | 0.4423 | |
| Ca/a | 0.0118 | Ca | 0.1118 | Cab | 0.0769 | |
| Ca/b | 0.0769 | Ca | 0.1346 | |||
| Cb/- | 0.3504 | |||||
| Ca/- | 0.1229 | |||||
| M (n = 52) | M-/- | 0.0192 | M- | 0.0734 | M- | 0.0192 |
| Mc/c | 0.1225 | Ma | 0.0481 | Mac | 0.0962 | |
| Mc/- | 0.1082 | Mc | 0.5516 | Mc | 0.2308 | |
| Ma/c | 0.0963 | Mb | 0.2596 | Mbc | 0.5192 | |
| Mb/c | 0.5192 | Mab | 0.0673 | Mabc | 0.1346 | |
| Mab/c | 0.1346 | |||||
| D (n = 52) | Da/a | 0.0192 | Da | 0.1340 | Da | 0.2500 |
| D-/- | 0.7500 | D- | 0.8660 | D- | 0.7500 | |
| Da/- | 0.2308 | |||||
| R (n = 37) | RR/R | 0.0541 | R | 0.2466 | R | 0.4324 |
| RR/r | 0.3784 | r | 0.7534 | O | 0.5676 | |
| Rr/r | 0.5675 | |||||
| I (R = 52) | II/I | 0.2115 | I | 0.4629 | I | 0.7115 |
| II/i | 0.5000 | i | 0.5371 | i | 0.2885 | |
| Ii/i | 0.2885 |
| System | Genotype | Frequency | Allele | Frequency | Phenotypic Classes | Frequency |
|---|---|---|---|---|---|---|
| Occurrence | Occurrence | Occurrence | ||||
| B-/- | 0.099 | B- | 0.2533 | Bbdg | 0.1923 | |
| Bb/- | 0.0186 | Bb | 0.0412 | Bbeig | 0.0769 | |
| Bg/g | 0.0007 | Bd | 0.0585 | Bdg | 0.0769 | |
| Bb/g | 0.0385 | Be | 0.0075 | B- | 0.1154 | |
| Bd/g | 0.0769 | Bi | 0.0249 | Bdeg | 0.0576 | |
| Bbdg/- | 0.1923 | Bg | 0.0928 | Bbdig | 0.2885 | |
| Bbdg/b | 0.0107 | Bbde | 0.0033 | Bdeig | 0.0192 | |
| B | Bbdg/d | 0.0178 | Bbdi | 0.056 | Bg | 0.0192 |
| Bbdg/g | 0.0113 | Bbdg | 0.1808 | Bbdeig | 0.0385 | |
| Bbdg/bdg | 0.009 | Bbei | 0.0405 | Bbei | 0.0385 | |
| Bbei/- | 0.0306 | Bbeg | 0.0179 | Bbg | 0.0385 | |
| Bbei/b | 0.0022 | Bbig | 0.0961 | Bbig | 0.0385 | |
| Bbei/e | 0.0022 | Bdei | 0.0078 | |||
| Bbei/i | 0.0022 | Bdeg | 0.0379 | |||
| Bbei/bei | 0.0012 | Bdig | 0.0591 | |||
| Bbig/- | 0.0305 | Beig | 0.0224 | |||
| Bbig/b | 0.0022 | |||||
| Bbig/i | 0.0022 | |||||
| Bbig/g | 0.0023 | |||||
| Bbig/big | 0.0012 | |||||
| Bdeg/- | 0.0457 | |||||
| Bdeg/d | 0.0033 | |||||
| Bdeg/e | 0.0033 | |||||
| Bdeg/g | 0.0034 | |||||
| Bdeg/deg | 0.0019 | |||||
| Bbdi/g | 0.0037 | |||||
| Bbdg/i | 0.0363 | |||||
| Bbig/d | 0.0242 | |||||
| Bdig/b | 0.0218 | |||||
| Bbdi/bdg | 0.0363 | |||||
| Bbdi/big | 0.0242 | |||||
| Bbdg/eig | 0.0403 | |||||
| Bbdi/dig | 0.0218 | |||||
| Bbdg/dig | 0.0363 | |||||
| Bbig/dig | 0.0242 | |||||
| Bbei/g | 0.0032 | |||||
| Bbeg/i | 0.007 | |||||
| Bbig/e | 0.0078 | |||||
| Beig/b | 0.007 | |||||
| Bbei/beg | 0.0078 | |||||
| Bbei/big | 0.0086 | |||||
| Bbeg/big | 0.0078 | |||||
| Bbei/eig | 0.0078 | |||||
| Bbeg/big | 0.007 | |||||
| Bbig/eig | 0.0078 | |||||
| Bdei/g | 0.0018 | |||||
| Bdeg/I | 0.002 | |||||
| Bdig/e | 0.0018 | |||||
| Beig/d | 0.0018 | |||||
| Bdei/deg | 0.002 | |||||
| Bdei/dig | 0.0018 | |||||
| Bdeg/dig | 0.002 | |||||
| Bdei/eig | 0.0018 | |||||
| Bdeg/eig | 0.002 | |||||
| Bdig/eig | 0.0018 | |||||
| Bbde/big | 0.0023 | |||||
| Bbdi/beg | 0.0021 | |||||
| Bbdg/bei | 0.0038 | |||||
| Bbdi/dig | 0.0021 | |||||
| Bbdi/deg | 0.0025 | |||||
| Bbdg/dei | 0.0035 | |||||
| Bbde/eig | 0.0021 | |||||
| Bbei/deg | 0.0027 | |||||
| Bbeg/dei | 0.0021 | |||||
| Bbdi/eig | 0.0021 | |||||
| Bbei/dig | 0.0023 | |||||
| Bbig/dei | 0.0023 | |||||
| Bbdg/eig | 0.0035 | |||||
| Bbeg/dig | 0.0021 | |||||
| Bbig/deg | 0.0027 |
| Indicators | Genotype at the BMP-15 Locus | ||
|---|---|---|---|
| WW | WM | MM | |
| Number of identified ewes with genotypes (n = 50) | 17 | 26 | 7 |
| Frequency of genotypes | 0.34 | 0.52 | 0.14 |
| Number of barren ewes in 3 years (%) | 4 (23.5%) | 5 (19.2%) | 1 (14.2%) |
| The number of non-productive ewes for 3 years (%) | 4 (23.5%) | 3 (11.2%) | 0 (0%) |
| Number of productive ewes (n = 43) | 13 | 23 | 7 |
| Obtained lambs for 3 years | 92 | 181 | 63 |
| Weaning lambs for 3 years | 85 | 170 | 56 |
| Obtained lambs per ewe on average per year | 2.3 ± 0.4 | 2.6 ± 0.4 | 3.0 ± 0.3 |
| Weaning lambs per ewe on average per year | 2.2 ± 0.4 | 2.5 ± 0.4 | 2.7 ± 0.3 |
| Genotype | Allele Frequency | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Locus | Ewe Numbers | WW | WM | MM | W | M | χ2 | Df | p | |
| BMP-15 | 48 | Obs | 35 | 13 | 0 | 0.865 | 0.135 | 1.18 | 1 | 0.6 |
| FecXG | Exp | 35.88 | 11.24 | 0.88 | ||||||
| BMPR1B | 48 | Obs | 34 | 14 | 0 | 0.854 | 0.146 | 1.4 | 1 | 0.497 |
| FecB | Exp | 35.02 | 11.96 | 1.02 | ||||||
| Breeds | Occurrence of Genotypes at the Hemoglobin Locus | Allele Frequency | χ2 | Df | p | |||
|---|---|---|---|---|---|---|---|---|
| AA | AB | BB | A | B | ||||
| Populations of sheep of the Romanov breed from various regions of Russia | ||||||||
| I, n = 50 | 24 | 19 | 7 | 0.67 | 0.33 | 1 | 1 | >0.05 |
| II, n = 47 | 25 | 17 | 5 | 0.713 | 0.287 | 0.64 | 1 | >0.05 |
| III, n = 40 | 14 | 20 | 6 | 0.6 | 0.4 | 0.168 | 1 | >0.05 |
| Coarse wool breeds | ||||||||
| Bozakh, n = 40 | 2 | 4 | 34 | 0.1 | 0.9 | 7.9 | 1 | <0.01 ** |
| Karabakh, n = 40 | 0 | 0 | 40 | 0 | 1 | 0 | 1 | >0.05 |
| Karakul, n = 40 | 3 | 12 | 25 | 0.225 | 0.775 | 0.782 | 1 | >0.05 |
| Semi-fine wool breeds | ||||||||
| Kuibyshev, n = 40 | 6 | 10 | 24 | 0.275 | 0.725 | 5.55 | 1 | <0.05 * |
| North Caucasian meat-wool, n = 40 | 2 | 11 | 27 | 0.188 | 0.812 | 0.39 | 1 | >0.05 |
| Tsigay, n = 40 | 1 | 7 | 32 | 0.1125 | 0.8875 | 0.602 | 1 | >0.05 |
| Fine wool breeds | ||||||||
| Azerbaijan merino, n = 40 | 2 | 3 | 35 | 0.0875 | 0.9125 | 11.09 | 1 | <0.001 *** |
| Volgogradsk, n = 40 | 5 | 18 | 17 | 0.35 | 0.65 | 0.005 | 1 | >0.05 |
| Caucasian, n = 40 | 1 | 18 | 21 | 0.25 | 0.75 | 1.6 | 1 | >0.05 |
| Breeds | Occurrence of Alleles at the Transferrin Locus | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TFA | TFB | TFC | TFD | TFE | TFI | TFP | Df | p | He | |
| Populations of sheep of the Romanov breed from various regions of Russia | ||||||||||
| I, n = 46 | 0.033 | 0.348 | 0.587 | 0.033 | 12 | 0.452 NS | 0.65 | |||
| II, n = 48 | 0.063 | 0.25 | 0.656 | 0.031 | 12 | 0.374 NS | 0.63 | |||
| III, n = 46 | 0.120 | 0.424 | 0.38 | 0.08 | 12 | 0.0001 *** | 0.93 | |||
| Coarse wool breeds | ||||||||||
| Bozakh, n = 40 | 0.075 | 0.225 | 0.475 | 0.2 | 0.025 | 12 | 0.513 NS | 0.70 | ||
| Karabakh, n = 40 | 0.113 | 0.237 | 0.325 | 0.237 | 0.088 | 17 | 0.45 NS | 0.70 | ||
| Karakul, n = 40 | 0.138 | 0.475 | 0.275 | 0.11 | 12 | 0.0092 ** | 0.73 | |||
| Semi-fine wool breeds | ||||||||||
| Kuibyshev, n = 39 | 0.4231 | 0.1026 | 0.321 | 0.013 | 0.01 | 20 | 0.446 NS | 0.74 | ||
| North Caucasian meat-wool, n = 40 | 0.225 | 0.2375 | 0.425 | 0.063 | 0.025 | 0.013 | 0.013 | 21 | 0.607 NS | 0.80 |
| Tsigay, n = 39 | 0.2308 | 0.1538 | 0.564 | 0.0513 | 21 | 0.459 NS | 0.51 | |||
| Fine wool breeds | ||||||||||
| Azerbaijan merino, n = 40 | 0.063 | 0.300 | 0.413 | 0.013 | 0.025 | 0.063 | 0.012 | 21 | 0.087 NS | 0.65 |
| Volgogradsk, n = 39 | 0.3333 | 0.359 | 0.179 | 0.090 | 0.013 | 0.003 | 16 | 0.098 NS | 0.56 | |
| Caucasian, n = 40 | 0.275 | 0.188 | 0.488 | 0.013 | 0.025 | 0.013 | 21 | 0.012 * | 0.48 | |
| Breeds | Albumin Locus Genotypes | Allele Frequency | He | Df | p | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AA | AB | AC | AD | BB | BC | BD | CC | CD | DD | A | B | C | D | ||||
| Populations of the Romanov sheep from various regions of Russia | |||||||||||||||||
| I, n = 46 | 3 | 2 | 20 | 12 | 9 | 0.055 | 0.38 | 0.315 | 0.25 | 1 | 8 | 0.009 ** | |||||
| II, n = 48 | 4 | 1 | 16 | 12 | 15 | 0.042 | 0.354 | 0.323 | 0.28 | 0.979 | 8 | 0.006 ** | |||||
| III, n = 50 | 2 | 18 | 9 | 1 | 8 | 1 | 1 | 0.31 | 0.28 | 0.4 | 0.01 | 0.72 | 8 | 0.036 * | |||
| Coarse wool breeds | |||||||||||||||||
| Bozakh, n = 40 | 4 | 14 | 8 | 1 | 6 | 6 | 1 | 0.325 | 0.063 | 0.412 | 0.2 | 0.825 | 8 | 0.403 NS | |||
| Karabakh, n = 40 | 4 | 9 | 4 | 12 | 11 | 0 | 0.163 | 0.3 | 0.537 | 0.625 | 4 | 0.994 NS | |||||
| Karakul, n = 40 | 1 | 1 | 12 | 19 | 2 | 4 | 1 | 0.025 | 0.575 | 0.35 | 0.05 | 0.575 | 8 | 0.992 NS | |||
| Semi-fine wool breeds | |||||||||||||||||
| Kuibyshev, n = 40 | 1 | 1 | 6 | 20 | 6 | 5 | 1 | 0.025 | 0.488 | 0.4 | 0.087 | 0.725 | 8 | 0.700 NS | |||
| North Caucasian meat-wool, n = 40 | 2 | 15 | 9 | 11 | 3 | 0 | 0.35 | 0.325 | 0.325 | 0.6 | 4 | 0.012 * | |||||
| Tsigay, n = 40 | 6 | 30 | 1 | 3 | 0 | 0.538 | 0.45 | 0.012 | 0.775 | 4 | 0.046 * | ||||||
| Fine-wool breeds | |||||||||||||||||
| Azerbaijan merino, n = 40 | 6 | 13 | 19 | 1 | 1 | 0 | 0.55 | 0.2 | 0.25 | 0.825 | 4 | 0.084 NS | |||||
| Volgogradsk, n = 40 | 5 | 15 | 20 | 0 | 0.5625 | 0.1875 | 0.25 | 0.875 | 4 | 0.009 ** | |||||||
| Caucasian, n = 40 | 1 | 2 | 6 | 19 | 2 | 10 | 0.038 | 0.425 | 0.512 | 0.025 | 0.6 | 8 | 0.893 NS | ||||
| Breeds | Prealbumin Locus Genotypes | Allele Frequency | Df | p | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FF | FS | FO | SS | SO | OO | F | S | O | He | |||
| Populations of sheep of the Romanov breed from various regions of Russia | ||||||||||||
| I, n = 46 | 19 | 6 | 21 | 0.272 | 0.435 | 0.293 | 1 | 4 | 0.0011 ** | |||
| II, n = 48 | 11 | 7 | 30 | 0.188 | 0.427 | 0.385 | 1 | 4 | 0.0032 ** | |||
| III, n = 50 | 13 | 29 | 8 | 0.42 | 0.21 | 0.37 | 1 | 4 | 0.0025 ** | |||
| Coarse wool breeds | ||||||||||||
| Bozakh, n = 40 | 1 | 20 | 6 | 4 | 9 | 0.35 | 0.463 | 0.188 | 0.88 | 4 | 0.25 NS | |
| Karabakh, n = 40 | 1 | 23 | 7 | 3 | 6 | 0.4 | 0.438 | 0.163 | 0.90 | 4 | 0.11 NS | |
| Karakul, n = 40 | 1 | 20 | 10 | 1 | 8 | 0.4 | 0.375 | 0.225 | 0.95 | 4 | 0.07 NS | |
| Semi-fine wool breeds | ||||||||||||
| Kuibyshev, n = 40 | 4 | 26 | 4 | 3 | 3 | 0.475 | 0.438 | 0.088 | 0.83 | 4 | 0.19 NS | |
| North Caucasian meat-wool, n = 40 | 6 | 15 | 14 | 2 | 3 | 0.513 | 0.275 | 0.213 | 0.80 | 4 | 0.49 NS | |
| Tsigay, n = 40 | 5 | 23 | 4 | 3 | 5 | 0.463 | 0.425 | 0.113 | 0.80 | 4 | 0.63 NS | |
| Fine-wool breeds | ||||||||||||
| Azerbaijan merino, n = 40 | 4 | 19 | 12 | 1 | 4 | 0.488 | 0.313 | 0.2 | 0.88 | 4 | 0.23 NS | |
| Volgogradsk, n = 40 | 5 | 17 | 15 | 1 | 2 | 0.525 | 0.263 | 0.213 | 0.85 | 4 | 0.15 NS | |
| Caucasian, n = 40 | 1 | 21 | 14 | 2 | 2 | 0.463 | 0.338 | 0.2 | 0.93 | 4 | 0.014 * | |
| Number Farms | n | HO | HE | AR | AM | F |
|---|---|---|---|---|---|---|
| 1 | 47 | 0.52 | 0.581 | 4.61 | 4.71 | 0.107 |
| 2 | 40 | 0.636 | 0.585 | 4.14 | 4.14 | −0.087 |
| 3 | 52 | 0.335 | 0.561 | 4.66 | 4.71 | 0.405 |
| 4 | 50 | 0.44 | 0.424 | 2.56 | 2.57 | −0.039 |
| 5 | 50 | 0.574 | 0.569 | 3.96 | 4.00 | −0.01 |
| 6 | 40 | 0.614 | 0.579 | 4.27 | 4.29 | −0.061 |
| 7 | 39 | 0.553 | 0.571 | 4.14 | 4.14 | 0.032 |
| 8 | 40 | 0.596 | 0.526 | 4.11 | 4.14 | −0.135 |
| 9 | 40 | 0.579 | 0.558 | 4.41 | 4.43 | −0.037 |
| 10 | 50 | 0.272 | 0.348 | 2.85 | 2.86 | 0.221 |
| 11 | 50 | 0.411 | 0.412 | 2.71 | 2.71 | 0.001 |
| 12 | 38 | 0.541 | 0.548 | 3.86 | 3.86 | 0.012 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marzanov, N.S.; Devrishov, D.A.; Ozerov, M.Y.; Maluchenko, O.P.; Marzanova, S.N.; Shukurova, E.B.; Koreckaya, E.A.; Kantanen, J.; Petit, D. The Significance of a Multilocus Analysis for Assessing the Biodiversity of the Romanov Sheep Breed in a Comparative Aspect. Animals 2023, 13, 1320. https://doi.org/10.3390/ani13081320
Marzanov NS, Devrishov DA, Ozerov MY, Maluchenko OP, Marzanova SN, Shukurova EB, Koreckaya EA, Kantanen J, Petit D. The Significance of a Multilocus Analysis for Assessing the Biodiversity of the Romanov Sheep Breed in a Comparative Aspect. Animals. 2023; 13(8):1320. https://doi.org/10.3390/ani13081320
Chicago/Turabian StyleMarzanov, Nurbiy S., Davud A. Devrishov, Mikhail Y. Ozerov, Oleg P. Maluchenko, Saida N. Marzanova, Elena B. Shukurova, Elena A. Koreckaya, Juha Kantanen, and Daniel Petit. 2023. "The Significance of a Multilocus Analysis for Assessing the Biodiversity of the Romanov Sheep Breed in a Comparative Aspect" Animals 13, no. 8: 1320. https://doi.org/10.3390/ani13081320
APA StyleMarzanov, N. S., Devrishov, D. A., Ozerov, M. Y., Maluchenko, O. P., Marzanova, S. N., Shukurova, E. B., Koreckaya, E. A., Kantanen, J., & Petit, D. (2023). The Significance of a Multilocus Analysis for Assessing the Biodiversity of the Romanov Sheep Breed in a Comparative Aspect. Animals, 13(8), 1320. https://doi.org/10.3390/ani13081320