
Evaluation of Immunodiagnostic Performances of Neospora caninum Peroxiredoxin 2 (NcPrx2), Microneme 4 (NcMIC4), and Surface Antigen 1 (NcSAG1) Recombinant Proteins for Bovine Neosporosis

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of N. caninum Tachyzoites
2.2. Production of GST-Fused Recombinant Proteins NcPrx2, NcMIC4, and NcSAG1
2.3. Protein Identification by Mass Spectrometry
2.4. Serum Samples
2.4.1. Control Sera
2.4.2. Field Dairy Cattle Serum Samples
2.5. Indirect ELISA (iELISA)
2.6. IFAT
2.7. Statistical Analysis
3. Results
3.1. Identification of NcPrx2, NcMIC4, and NcSAG1 Recombinant Proteins
3.2. Assessment of Recombinant Proteins Using Mouse and Bovine Infected Sera
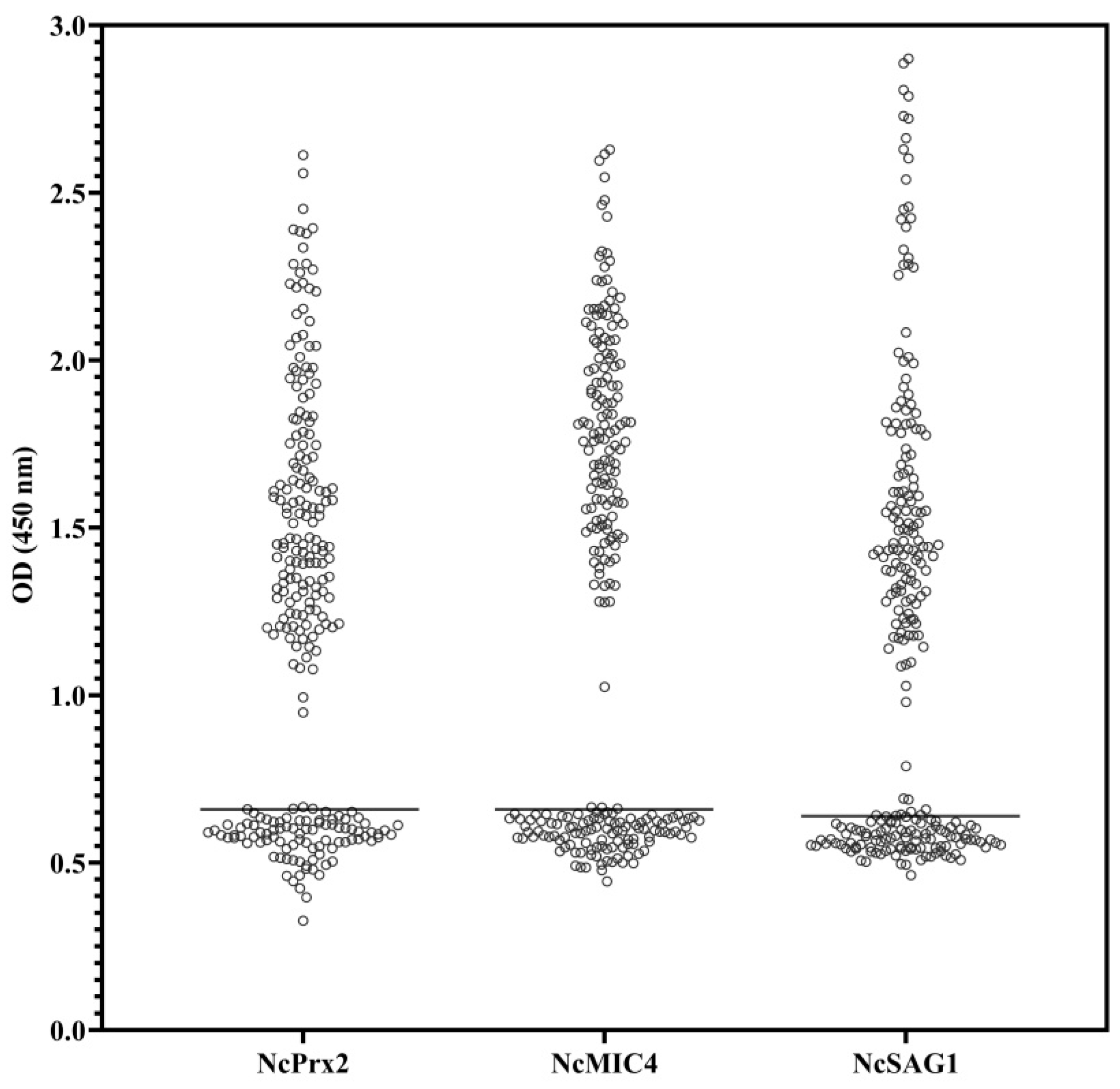
3.3. Comparison of ELISAs Using NcPrx2, NcMIC4, and NcSAG1 with IFAT in Field Cattle Serum Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P.; Schares, G.; Ortega-Mora, L.M. Epidemiology and control of neosporosis and Neospora caninum. Clin. Microbiol. Rev. 2007, 20, 323–367. [Google Scholar] [CrossRef] [PubMed]
- Innes, E.A.; Bartley, P.M.; Maley, S.W.; Wright, S.E.; Buxton, D. Comparative host-parasite relationships in ovine toxoplasmosis and bovine neosporosis and strategies for vaccination. Vaccine 2007, 25, 5495–5503. [Google Scholar] [CrossRef] [PubMed]
- Reichel, M.P.; Ayanegui-Alcérreca, M.A.; Gondim, L.F.; Ellis, J.T. What is the global economic impact of Neospora caninum in cattle–the billion dollar question. Int. J. Parasitol. 2013, 43, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Review of Neospora caninum and neosporosis in animals. Korean J. Parasitol. 2003, 41, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Horcajo, P.; Regidor-Cerrillo, J.; Aguado-Martínez, A.; Hemphill, A.; Ortega-Mora, L. Vaccines for bovine neosporosis: Current status and key aspects for development. Parasite Immunol. 2016, 38, 709–723. [Google Scholar] [CrossRef] [PubMed]
- Davison, H.C.; Otter, A.; Trees, A.J. Significance of Neospora caninum in British dairy cattle determined by estimation of seroprevalence in normally calving cattle and aborting cattle. Int. J. Parasitol. 1999, 29, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, N.; Fecteau, G.; Pare, J.; Martineau, R.; Villeneuve, A. Vertical and horizontal transmission of Neospora caninum in dairy herds in Quebec. Can. Vet. J. 2000, 41, 464–467. [Google Scholar] [PubMed]
- Mazuz, M.L.; Fish, L.; Reznikov, D.; Wolkomirsky, R.; Leibovitz, B.; Savitzky, I.; Golenser, J.; Shkap, V. Neosporosis in naturally infected pregnant dairy cattle. Vet. Parasitol. 2014, 205, 85–91. [Google Scholar] [CrossRef]
- Spilovska, S.; Reiterova, K.; Antolova, D. Neospora caninum—Associated Abortions in Slovak Dairy Farm. Iran. J. Parasitol. 2015, 10, 96–101. [Google Scholar]
- Lefkaditis, M.; Mpairamoglou, R.; Sossidou, A.; Spanoudis, K.; Tsakiroglou, M. Neospora caninum, A potential cause of reproductive failure in dairy cows from Northern Greece. Vet. Parasitol. Reg. Stud. Rep. 2020, 19, 100365. [Google Scholar] [CrossRef]
- Wang, X.; Song, X.; Yang, J.; Liu, Q.; Liu, J. Characterization of Neospora caninum microneme protein 26 and its potential use as a diagnostic marker for neosporosis in cattle. Front. Vet. Sci. 2020, 7, 357. [Google Scholar] [CrossRef]
- Dubey, J.P.; Schares, G. Diagnosis of bovine neosporosis. Vet. Parasitol. 2006, 140, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Ghalmi, F.; China, B.; Jenkins, M.; Azzag, N.; Losson, B. Comparison of different serological methods to detect antibodies specific to Neospora caninum in bovine and canine sera. J. Vet. Diagn. Investig. 2014, 26, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Otsuki, T.; Kato, T.; Park, E.Y. Development of a diagnostic method for neosporosis in cattle using recombinant Neospora caninum proteins. BMC Biotechnol. 2012, 12, 19. [Google Scholar] [CrossRef]
- Sinnott, F.A.; Monte, L.G.; Collares, T.F.; Silveira, R.M.; Borsuk, S. Review on the immunological and molecular diagnosis of neosporosis (years 2011–2016). Vet. Parasitol. 2017, 239, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Novoa, M.B.; Valentini, B.S.; Sarli, M.; Torioni-de-Echaide, S.M.; Primo, M.E.; Echaide, I.E. Evaluation of a competitive inhibition ELISA based on the recombinant protein tSAG1 to detect anti-Neospora caninum antibodies in cattle. J. Vet. Diagn. Invest. 2020, 32, 401–408. [Google Scholar] [CrossRef]
- Lee, E.G.; Kim, J.H.; Shin, Y.S.; Shin, G.W.; Kim, Y.H.; Kim, G.S.; Kim, D.Y.; Jung, T.S.; Suh, M.D. Two-dimensional gel electrophoresis and immunoblot analysis of Neospora caninum tachyzoites. J. Vet. Sci. 2004, 5, 139–145. [Google Scholar] [CrossRef]
- Shin, Y.S.; Lee, E.G.; Shin, G.W.; Kim, Y.R.; Lee, E.Y.; Kim, J.H.; Jang, H.; Gershwin, L.J.; Kim, D.Y.; Kim, Y.H.; et al. Identification of antigenic proteins from Neospora caninum recognized by bovine immunoglobulins M, E, A and G using immunoproteomics. Proteomics 2004, 4, 3600–3609. [Google Scholar] [CrossRef]
- Lee, E.G.; Kim, J.H.; Shin, Y.S.; Shin, G.W.; Suh, M.D.; Kim, D.Y.; Kim, Y.H.; Kim, G.S.; Jung, T.S. Establishment of a two-dimensional electrophoresis map for Neospora caninum tachyzoites by proteomics. Proteomics 2003, 3, 2339–2350. [Google Scholar] [CrossRef]
- Udonsom, R.; Reamtong, O.; Adisakwattana, P.; Popruk, S.; Jirapattharasate, C.; Nishikawa, Y.; Inpankaew, T.; Toompong, J.; Kotepui, M.; Mahittikorn, A. Immunoproteomics to identify species-specific antigens in Neospora caninum recognised by infected bovine sera. Parasite 2022, 29, 60. [Google Scholar] [CrossRef]
- Venancio-Brochi, J.C.; Pereira, L.M.; Baroni, L.; Abreu-Filho, P.G.; Yatsuda, A.P. Characterization of the Neospora caninum peroxiredoxin: A novel peroxidase and antioxidant enzyme. Parasitol. Res. 2022, 121, 1735–1748. [Google Scholar] [CrossRef]
- Fereig, R.M.; Kuroda, Y.; Terkawi, M.A.; Mahmoud, M.E.; Nishikawa, Y. Immunization with Toxoplasma gondii peroxiredoxin 1 induces protective immunity against toxoplasmosis in mice. PLoS ONE 2017, 12, e0176324. [Google Scholar] [CrossRef]
- Fereig, R.M.; Nishikawa, Y. Peroxiredoxin 3 promotes IL-12 production from macrophages and partially protects mice against infection with Toxoplasma gondii. Parasitol. Int. 2016, 65, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Bayih, A.G.; Daifalla, N.S.; Gedamu, L. DNA-protein immunization using Leishmania peroxidoxin-1 induces a strong CD4+ T cell response and partially protects mice from cutaneous leishmaniasis: Role of fusion murine granulocyte-macrophage colony-stimulating factor DNA adjuvant. PLoS Negl. Trop. Dis. 2014, 8, e3391. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.; Riesen, M.; Naguleswaran, A.; Vonlaufen, N.; Stettler, R.; Leepin, A.; Wastling, J.M.; Hemphill, A. Identification and characterization of a Neospora caninum microneme-associated protein (NcMIC4) that exhibits unique lactose-binding properties. Infect. Immun. 2004, 72, 4791–4800. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Mueller, J.; Suana, A.; Hemphill, A. Vaccination with microneme protein NcMIC4 increases mortality in mice inoculated with Neospora caninum. J. Parasitol. 2007, 93, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, E.V.; Bernardes, E.S.; Silva, N.M.; Mineo, J.R.; Panunto-Castelo, A.; Roque-Barreira, M.C. Immunization with MIC1 and MIC4 induces protective immunity against Toxoplasma gondii. Microbes Infect. 2006, 8, 1244–1251. [Google Scholar] [CrossRef] [PubMed]
- Pinzan, C.F.; Sardinha-Silva, A.; Almeida, F.; Lai, L.; Lopes, C.D.; Lourenco, E.V.; Panunto-Castelo, A.; Matthews, S.; Roque-Barreira, M.C. Vaccination with Recombinant Microneme Proteins Confers Protection against Experimental Toxoplasmosis in Mice. PLoS ONE 2015, 10, e0143087. [Google Scholar] [CrossRef]
- Takashima, Y.; Takasu, M.; Yanagimoto, I.; Hattori, N.; Batanova, T.; Nishikawa, Y.; Kitoh, K. Prevalence and dynamics of antibodies against NcSAG1 and NcGRA7 antigens of Neospora caninum in cattle during the gestation period. J. Vet. Med. Sci. 2013, 75, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Abdelbaky, H.H.; Nishimura, M.; Shimoda, N.; Hiasa, J.; Fereig, R.M.; Tokimitsu, H.; Inokuma, H.; Nishikawa, Y. Evaluation of Neospora caninum serodiagnostic antigens for bovine neosporosis. Parasitol. Int. 2020, 75, 102045. [Google Scholar] [CrossRef]
- Fereig, R.M.; Abdelbaky, H.H.; Nishikawa, Y. Comparative Evaluation of Four Potent Neospora caninum Diagnostic Antigens Using Immunochromatographic Assay for Detection of Specific Antibody in Cattle. Microorganisms 2021, 9, 2133. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Kohara, J.; Kuroda, Y.; Hiasa, J.; Tanaka, S.; Muroi, Y.; Kojima, N.; Furuoka, H.; Nishikawa, Y. Oligomannose-coated liposome-entrapped dense granule protein 7 induces protective immune response to Neospora caninum in cattle. Vaccine 2013, 31, 3528–3535. [Google Scholar] [CrossRef] [PubMed]
- Wiengcharoen, J.; Thompson, R.C.; Nakthong, C.; Rattanakorn, P.; Sukthana, Y. Transplacental transmission in cattle: Is Toxoplasma gondii less potent than Neospora caninum? Parasitol. Res. 2011, 108, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Wiengcharoen, J.; Nakthong, C.; Mitchaothai, J.; Udonsom, R.; Sukthana, Y. Toxoplasmosis and neosporosis among beef cattle slaughtered for food in Western Thailand. Southeast. Asian J. Trop. Med. Public. Health 2012, 43, 1087. [Google Scholar] [PubMed]
- McHugh, M.L. Interrater reliability: The kappa statistic. Biochem Med. (Zagreb) 2012, 22, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Gondim, L.F.; Mineo, J.R.; Schares, G. Importance of serological cross-reactivity among Toxoplasma gondii, Hammondia spp., Neospora spp., Sarcocystis spp. and Besnoitia besnoiti. Parasitology 2017, 144, 851–868. [Google Scholar] [CrossRef]
- Chahan, B.; Gaturaga, I.; Huang, X.; Liao, M.; Fukumoto, S.; Hirata, H.; Nishikawa, Y.; Suzuki, H.; Sugimoto, C.; Nagasawa, H.; et al. Serodiagnosis of Neospora caninum infection in cattle by enzyme-linked immunosorbent assay with recombinant truncated NcSAG1. Vet. Parasitol. 2003, 118, 177–185. [Google Scholar] [CrossRef]
- Kubota, N.; Sakata, Y.; Miyazaki, N.; Itamoto, K.; Bannai, H.; Nishikawa, Y.; Xuan, X.; Inokuma, H. Serological survey of Neospora caninum infection among dogs in Japan through species-specific ELISA. J. Vet. Med. Sci. 2008, 70, 869–872. [Google Scholar] [CrossRef]
- Udonsom, R.; Sukthana, Y.; Nishikawa, Y.; Fereig, R.M.; Jirapattharasate, C. Current situation of Neospora caninum and Toxoplasma gondii infection among beef cattle in Kanchanaburi, Ratchaburi and Nakhon Patom provinces, Thailand. Thai J. Vet. Med. 2018, 48, 403–409. [Google Scholar] [CrossRef]
- Howe, D.K.; Tang, K.; Conrad, P.A.; Sverlow, K.; Dubey, J.P.; Sibley, L.D. Sensitive and specific identification of Neospora caninum infection of cattle based on detection of serum antibodies to recombinant Ncp29. Clin. Diagn. Lab. Immunol. 2002, 9, 611–615. [Google Scholar] [CrossRef]
- Alves Sinnott, F.; da Silva Leal, K.; de Oliveira Silva, M.T.; Barros de Pinho, R.; Pappen, F.; da Rosa Farias, N.A.; Bedoya Llano, H.A.; Pereira Garcia Melo, D.; Borsuk, S. An indirect ELISA for Neosporosis: Associating recombinant Neospora caninum proteins NcSRS2 and NcSAG1. Vet. Parasitol. 2020, 281, 109101. [Google Scholar] [CrossRef]
- McGonigle, S.; Dalton, J.P.; James, E.R. Peroxidoxins: A new antioxidant family. Parasitol. Today 1998, 14, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Lindsay, D.S. A review of Neospora caninum and neosporosis. Vet. Parasitol. 1996, 67, 1–59. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Neosporosis in cattle: Biology and economic impact. J. Am. Vet. Med. Assoc. 1999, 214, 1160–1163. [Google Scholar] [CrossRef]
- Ghalmi, F.; China, B.; Ghalmi, A.; Hammitouche, D.; Losson, B. Study of the risk factors associated with Neospora caninum seroprevalence in Algerian cattle populations. Res. Vet. Sci. 2012, 93, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, C.; Uggla, A. Serological diagnosis of Neospora caninum infection. Int. J. Parasitol. 1999, 29, 1497–1507. [Google Scholar] [CrossRef]
- Atkinson, R.; Harper, P.; Reichel, M.; Ellis, J. Progress in the serodiagnosis of Neospora caninum infections of cattle. Parasitol. Today 2000, 16, 110–114. [Google Scholar] [CrossRef]
- Alvarez-Garcia, G.; Collantes-Fernandez, E.; Costas, E.; Rebordosa, X.; Ortega-Mora, L.M. Influence of age and purpose for testing on the cut-off selection of serological methods in bovine neosporosis. Vet. Res. 2003, 34, 341–352. [Google Scholar] [CrossRef]
- Dubey, J.P.; Jenkins, M.C.; Adams, D.S.; McAllister, M.M.; Anderson-Sprecher, R.; Baszler, T.V.; Kwok, O.C.; Lally, N.C.; Bjorkman, C.; Uggla, A. Antibody responses of cows during an outbreak of neosporosis evaluated by indirect fluorescent antibody test and different enzyme-linked immunosorbent assays. J. Parasitol. 1997, 83, 1063–1069. [Google Scholar] [CrossRef]
- von Blumroder, D.; Schares, G.; Norton, R.; Williams, D.J.; Esteban-Redondo, I.; Wright, S.; Bjorkman, C.; Frossling, J.; Risco-Castillo, V.; Fernandez-Garcia, A.; et al. Comparison and standardisation of serological methods for the diagnosis of Neospora caninum infection in bovines. Vet. Parasitol. 2004, 120, 11–22. [Google Scholar] [CrossRef]
- Silva, D.A.; Lobato, J.; Mineo, T.W.; Mineo, J.R. Evaluation of serological tests for the diagnosis of Neospora caninum infection in dogs: Optimization of cut off titers and inhibition studies of cross-reactivity with Toxoplasma gondii. Vet. Parasitol. 2007, 143, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Benetti, A.H.; Schein, F.B.; dos Santos, T.R.; Toniollo, G.H.; da Costa, A.J.; Mineo, J.R.; Lobato, J.; de Oliveira Silva, D.A.; Gennari, S.M. [Inquiry of antibodies anti-Neospora caninum in dairy cattle, dogs and rural workers of the south-west region of Mato Grosso State]. Rev. Bras. Parasitol. Vet. 2009, 18 (Suppl. S1), 29–33. [Google Scholar] [CrossRef] [PubMed]
- Campero, L.M.; Minke, L.; More, G.; Rambeaud, M.; Bacigalupe, D.; Moore, D.P.; Hecker, Y.; Campero, C.M.; Schares, G.; Venturini, M.C. Evaluation and comparison of serological methods for the detection of bovine neosporosis in Argentina. Rev. Argent. Microbiol. 2015, 47, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Sondgen, P.; Peters, M.; Barwald, A.; Wurm, R.; Holling, F.; Conraths, F.J.; Schares, G. Bovine neosporosis: Immunoblot improves foetal serology. Vet. Parasitol. 2001, 102, 279–290. [Google Scholar] [CrossRef]
- Staubli, D.; Nunez, S.; Sager, H.; Schares, G.; Gottstein, B. Neospora caninum immunoblotting improves serodiagnosis of bovine neosporosis. Parasitol. Res. 2006, 99, 648–658. [Google Scholar] [CrossRef]
- Ortega-Mora, L.; Fernández-García, A.; Gómez-Bautista, M. Diagnosis of bovine neosporosis: Recent advances and perspectives. Acta Parasitol. 2006, 51, 1–14. [Google Scholar] [CrossRef]
- Chanlun, A.; Emanuelson, U.; Frossling, J.; Aiumlamai, S.; Bjorkman, C. A longitudinal study of seroprevalence and seroconversion of Neospora caninum infection in dairy cattle in northeast Thailand. Vet. Parasitol. 2007, 146, 242–248. [Google Scholar] [CrossRef]
- Inpankaew, T.; Jittapalapong, S.; Mitchell, T.J.; Sununta, C.; Igarashi, I.; Xuan, X. Seroprevalence of Neospora caninum infection in dairy cows in Northern provinces, Thailand. Acta Parasitol. 2014, 59, 305–309. [Google Scholar] [CrossRef]
- Arunvipas, P.; Inpankaew, T.; Jittapalapong, S. Seroprevalence and risk factors of Neospora caninum infection among dairy cows in the Western Provinces of Nakhon Pathom, Ratchaburi and Kanchanaburi, Thailand. Agric. Nat. Resour. 2012, 46, 64–70. [Google Scholar]
- Schares, G.; Rauser, M.; Zimmer, K.; Peters, M.; Wurm, R.; Dubey, J.P.; de Graaf, D.C.; Edelhofer, R.; Mertens, C.; Hess, G.; et al. Serological differences in Neospora caninum-associated epidemic and endemic abortions. J. Parasitol. 1999, 85, 688–694. [Google Scholar] [CrossRef]
- Jenkins, M.C.; Caver, J.A.; Bjorkman, C.; Anderson, T.C.; Romand, S.; Vinyard, B.; Uggla, A.; Thulliez, P.; Dubey, J.P. Serological investigation of an outbreak of Neospora caninum-associated abortion in a dairy herd in southeastern United States. Vet. Parasitol. 2000, 94, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Reichel, M.P.; Ellis, J.T. If control of Neospora caninum infection is technically feasible does it make economic sense? Vet. Parasitol. 2006, 142, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Wouda, W.; Bartels, C.J.; Moen, A.R. Characteristics of Neospora caninum-associated abortion storms in diary herds in The Netherlands (1995 to 1997). Theriogenology 1999, 52, 233–245. [Google Scholar] [CrossRef]
- Asmare, K.; Regassa, F.; Robertson, L.J.; Skjerve, E. Seroprevalence of Neospora caninum and associated risk factors in intensive or semi-intensively managed dairy and breeding cattle of Ethiopia. Vet. Parasitol. 2013, 193, 85–94. [Google Scholar] [CrossRef] [PubMed]
- McAllister, M.M. Diagnosis and Control of Bovine Neosporosis. Vet. Clin. N. Am. Food Anim. Pract. 2016, 32, 443–463. [Google Scholar] [CrossRef]
- Guimarães, J.S., Jr.; Souza, S.L.; Bergamaschi, D.P.; Gennari, S.M. Prevalence of Neospora caninum antibodies and factors associated with their presence in dairy cattle of the north of Paraná state, Brazil. Vet. Parasitol. 2004, 124, 1–8. [Google Scholar] [CrossRef]
- Fávero, J.F.; Da Silva, A.S.; Campigotto, G.; Machado, G.; Daniel de Barros, L.; Garcia, J.L.; Vogel, F.F.; Mendes, R.E.; Stefani, L.M. Risk factors for Neospora caninum infection in dairy cattle and their possible cause-effect relation for disease. Microb. Pathog. 2017, 110, 202–207. [Google Scholar] [CrossRef]
- Manca, R.; Ciccarese, G.; Scaltrito, D.; Chirizzi, D. Detection of Anti-Neospora caninum Antibodies on Dairy Cattle Farms in Southern Italy. Vet. Sci. 2022, 9, 87. [Google Scholar] [CrossRef]
- Selim, A.; Alshammari, A.; Gattan, H.S.; Marzok, M.; Salem, M.; Al-Jabr, O.A. Neospora caninum infection in dairy cattle in Egypt: A serosurvey and associated risk factors. Sci. Rep. 2023, 13, 15489. [Google Scholar] [CrossRef]
- Anderson, M.L.; Andrianarivo, A.G.; Conrad, P.A. Neosporosis in cattle. Anim. Reprod. Sci. 2000, 60–61, 417–431. [Google Scholar] [CrossRef]
- Bartels, C.J.; Huinink, I.; Beiboer, M.L.; van Schaik, G.; Wouda, W.; Dijkstra, T.; Stegeman, A. Quantification of vertical and horizontal transmission of Neospora caninum infection in Dutch dairy herds. Vet. Parasitol. 2007, 148, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Japa, O.; Nuangmek, A.; Prakhammin, K.; Flynn, R.J. Prevalence of vertically transmitted Neospora caninum amongst beef cattle in Phayao, Thailand. Parasitol. Int. 2019, 70, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Hall, C.A.; Reichel, M.P.; Ellis, J.T. Neospora abortions in dairy cattle: Diagnosis, mode of transmission and control. Vet. Parasitol. 2005, 128, 231–241. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Farm | No. of Dairy Cattle | No. of Positive (%) | |||
|---|---|---|---|---|---|
| IFAT | NcPrx2 | NcMIC4 | NcSAG1 | ||
| 1 | 61 | 32 (52.4) | 36 (59.0) | 34 (55.7) | 27 (44.2) |
| 2 | 7 | 6 (85.7) | 4 (57.1) | 6 (85.7) | 6 (85.7) |
| 3 | 29 | 14 (48.2) | 15 (51.7) | 16 (55.1) | 14 (48.2) |
| 4 | 29 | 15 (51.7) | 20 (68.9) | 16 (55.1) | 19 (65.5) |
| 5 | 31 | 20 (64.5) | 14 (45.1) | 21 (67.7) | 21 (67.7) |
| 6 | 36 | 21 (58.3) | 28 (77.7) | 16 (44.4) | 25 (69.4) |
| 7 | 30 | 17 (56.6) | 21 (70.0) | 14 (46.6) | 14 (46.6) |
| 8 | 24 | 13 (54.1) | 17 (70.8) | 15 (62.5) | 11 (45.8) |
| Total | 247 | 138 (55.8) | 155 (62.7) | 138 (55.8) | 137(55.4) |
| IFAT | NcPrx2 | NcMIC4 | NcSAG1 | |||
|---|---|---|---|---|---|---|
| (+) | (−) | (+) | (−) | (+) | (−) | |
| (+) | 120 | 18 | 116 | 22 | 120 | 18 |
| (−) | 35 | 74 | 22 | 87 | 17 | 92 |
| Total | 155 | 92 | 138 | 109 | 137 | 110 |
| Parameters | NcPrx2 | NcMIC4 | NcSAG1 |
|---|---|---|---|
| Sensitivity (%) | 87.0 | 84.1 | 88.4 |
| Specificity (%) | 67.0 | 78.9 | 80.7 |
| Kappa value | 0.558 | 0.64 | 0.713 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Udonsom, R.; Adisakwattana, P.; Popruk, S.; Reamtong, O.; Jirapattharasate, C.; Thiangtrongjit, T.; Rerkyusuke, S.; Chanlun, A.; Hasan, T.; Kotepui, M.; et al. Evaluation of Immunodiagnostic Performances of Neospora caninum Peroxiredoxin 2 (NcPrx2), Microneme 4 (NcMIC4), and Surface Antigen 1 (NcSAG1) Recombinant Proteins for Bovine Neosporosis. Animals 2024, 14, 531. https://doi.org/10.3390/ani14040531
Udonsom R, Adisakwattana P, Popruk S, Reamtong O, Jirapattharasate C, Thiangtrongjit T, Rerkyusuke S, Chanlun A, Hasan T, Kotepui M, et al. Evaluation of Immunodiagnostic Performances of Neospora caninum Peroxiredoxin 2 (NcPrx2), Microneme 4 (NcMIC4), and Surface Antigen 1 (NcSAG1) Recombinant Proteins for Bovine Neosporosis. Animals. 2024; 14(4):531. https://doi.org/10.3390/ani14040531
Chicago/Turabian StyleUdonsom, Ruenruetai, Poom Adisakwattana, Supaluk Popruk, Onrapak Reamtong, Charoonluk Jirapattharasate, Tipparat Thiangtrongjit, Sarinya Rerkyusuke, Aran Chanlun, Tanjila Hasan, Manas Kotepui, and et al. 2024. "Evaluation of Immunodiagnostic Performances of Neospora caninum Peroxiredoxin 2 (NcPrx2), Microneme 4 (NcMIC4), and Surface Antigen 1 (NcSAG1) Recombinant Proteins for Bovine Neosporosis" Animals 14, no. 4: 531. https://doi.org/10.3390/ani14040531
APA StyleUdonsom, R., Adisakwattana, P., Popruk, S., Reamtong, O., Jirapattharasate, C., Thiangtrongjit, T., Rerkyusuke, S., Chanlun, A., Hasan, T., Kotepui, M., Siri, S., Nishikawa, Y., & Mahittikorn, A. (2024). Evaluation of Immunodiagnostic Performances of Neospora caninum Peroxiredoxin 2 (NcPrx2), Microneme 4 (NcMIC4), and Surface Antigen 1 (NcSAG1) Recombinant Proteins for Bovine Neosporosis. Animals, 14(4), 531. https://doi.org/10.3390/ani14040531