
Dietary 25-Hydroxycholecalciferol Supplementation as a Vitamin D3 Substitute Improves Performance, Egg Quality, Blood Indexes, Jejunal Morphology, and Tibia Quality in Late-Phase Laying Hens
 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds and Housing
2.2. Experimental Design and Diets
2.3. Sample Collection and Measurements
2.4. Production Performance and Egg Quality Analysis
2.5. Blood and Serum Biochemical Parameters Analyses
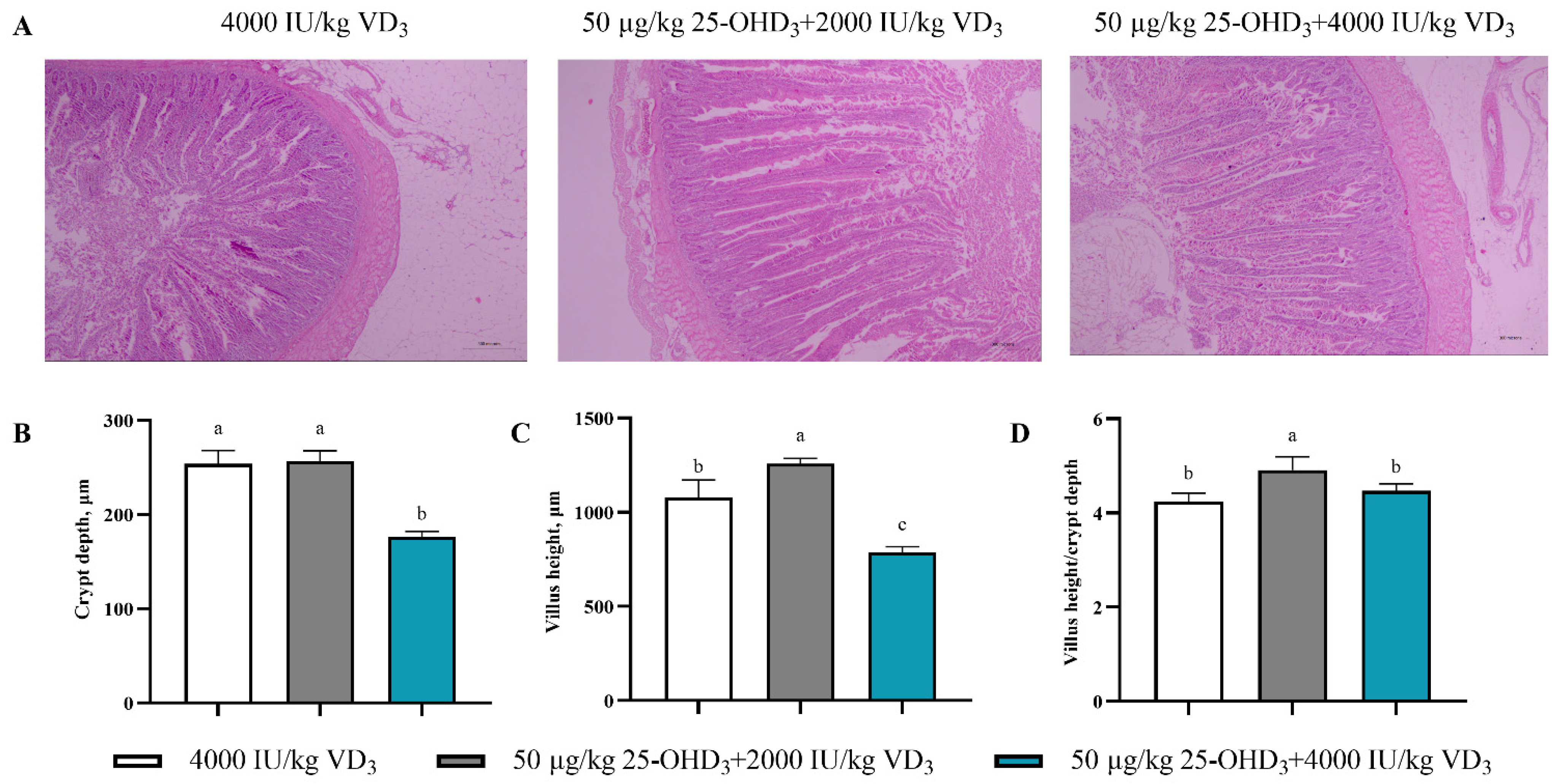
2.6. Intestinal Morphology Analysis
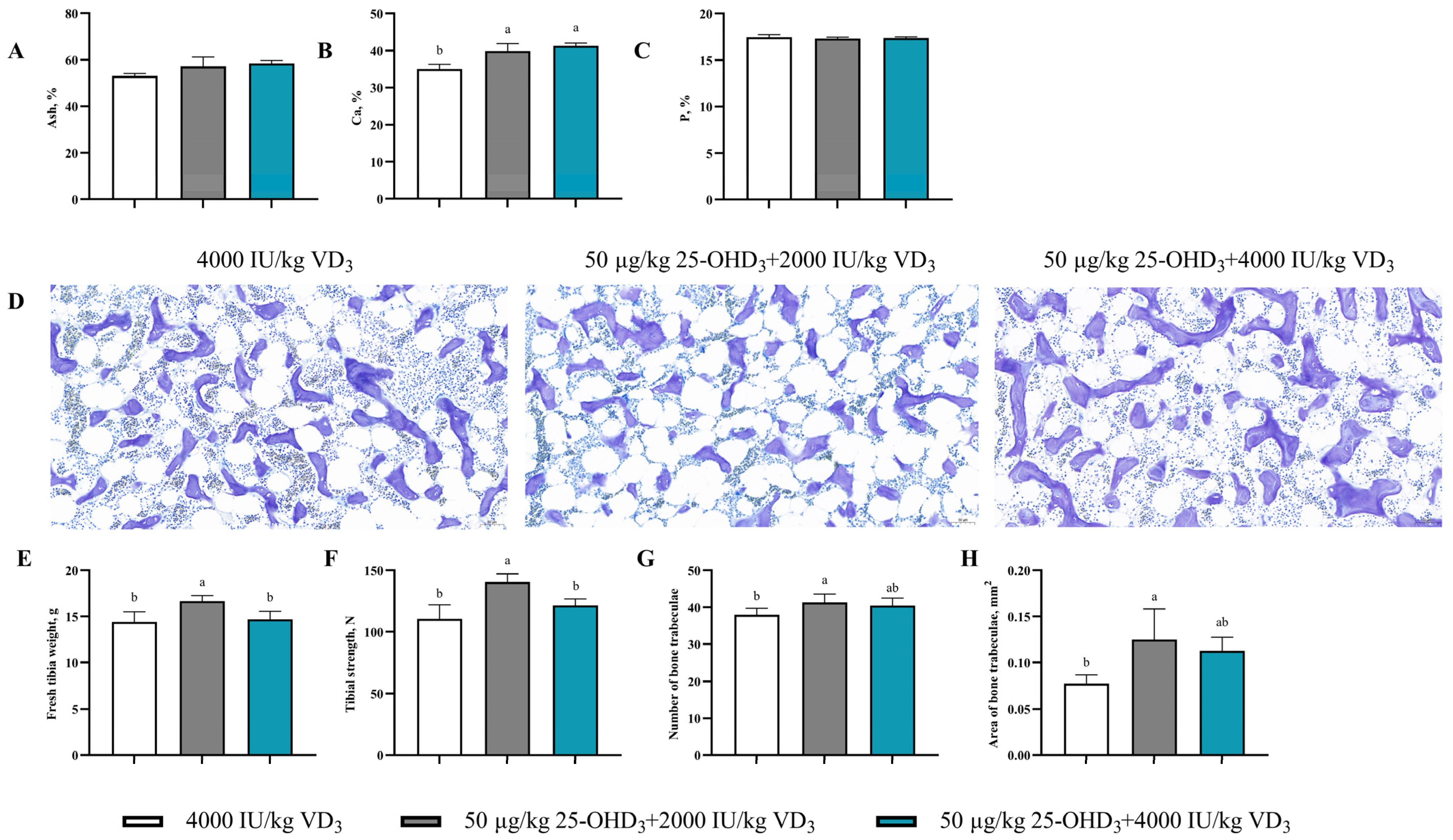
2.7. Tibial Quality Analysis
2.8. Statistical Analysis
3. Results
3.1. Production Performance
3.2. Egg Quality Assessment
3.3. Serum Biochemistry Analysis
3.4. Serum Hormone Concentration
3.5. Antioxidant Capacity
3.6. Intestinal Morphology
3.7. Tibia Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, A.C.; Wang, M.A.; Chen, L.; Long, C.; Guo, Y.; Sheng, X.H.; Wang, X.G.; Xing, K.; Xiao, L.F.; Ni, H.M.; et al. Effects of dietary pretreated Chinese herbal medicine supplementation on production performance, egg quality, uterine histopathological changes, and antioxidant capacity in late-phase laying hens. Front. Physiol. 2023, 14, 1110301. [Google Scholar] [CrossRef]
- Park, J.A.; Sohn, S.H. The influence of hen aging on eggshell ultrastructure and shell mineral components. Korean J. Food. Sci. Anim. Resour. 2018, 38, 1080–1091. [Google Scholar] [CrossRef]
- Kim, W.K.; Bloomfield, S.A.; Sugiyama, T.; Ricke, S.C. Concepts and methods for understanding bone metabolism in laying hens. World Poult. Sci. J. 2019, 68, 71–82. [Google Scholar] [CrossRef]
- Browning, L.C.; Cowieson, A.J. Interactive effects of vitamin D3 and strontium on performance, nutrient retention and bone mineral composition in laying hens. J. Sci. Food Agric. 2015, 95, 1080–1087. [Google Scholar] [CrossRef]
- Zhao, S.C.; Teng, X.Q.; Xu, D.L.; Chi, X.; Ge, M.; Xu, S.W. Influences of low level of dietary calcium on bone characters in laying hens. Poult. Sci. 2020, 99, 7084–7091. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, W.W.; Qi, G.H.; Cui, C.F.; Wu, S.G.; Zhang, H.J.; Xu, L.; Wang, J. Effects of dietary Bacillus subtilis supplementation and calcium levels on performance and eggshell quality of laying hens in the late phase of production. Poult. Sci. 2021, 100, 100970. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.K.; Coon, C. Sensitivity of various bone parameters of laying hens to different daily calcium intakes. Poult. Sci. 1990, 69, 2209–2213. [Google Scholar] [CrossRef] [PubMed]
- Olgun, O.; Aygun, A. Nutritional factors affecting the breaking strength of bone in laying hens. Proc. Nutr. Soc. 2016, 72, 821–832. [Google Scholar] [CrossRef]
- Nannoni, E.; Buonaiuto, G.; Martelli, G.; Lizzi, G.; Trevisani, G.; Garavini, G.; Sardi, L. Influence of increased freedom of movement on welfare and egg laying pattern of hens kept in aviaries. Animals 2022, 12, 2307. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.K.; Hester, P.Y.; Spurlock, D.M. Relationships of a transforming growth factor-β2 single nucleotide polymorphism and messenger ribonucleic acid abundance with bone and production traits in chickens. Poult. Sci. 2007, 86, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, C.C.; Fleming, R.H. Osteoporosis in cage layers. Poult. Sci. 2000, 79, 1033–1041. [Google Scholar] [CrossRef]
- Knowles, T.G.; Wilkins, L.J. The problem of broken bones during the handling of laying hens—A review. Poult. Sci. 1998, 77, 1798–1802. [Google Scholar] [CrossRef]
- Nascimento, G.D.; Murakami, A.; Guerra, A.; Ospinas-Rojas, I.; Ferreira, M.; Fanhani, J.C. Effect of different vitamin D sources and calcium levels in the diet of layers in the second laying cycle. Braz. J. Poult. Sci. 2014, 16, 37–42. [Google Scholar] [CrossRef]
- Geng, Y.; Ma, Q.; Wang, Z.; Guo, Y. Dietary vitamin D3 supplementation protects laying hens against lipopolysaccharide-induced immunological stress. Nutr. Metab. 2018, 15, 8. [Google Scholar] [CrossRef]
- Wen, J.; Livingston, K.A.; Persia, M.E. Effect of high concentrations of dietary vitamin D3 on pullet and laying hen performance, skeleton health, eggshell quality, and yolk vitamin D3 content when fed to W36 laying hens from day of hatch until 68 wk of age. Poult. Sci. 2019, 98, 6713–6720. [Google Scholar] [CrossRef]
- Mattila, P.; Valaja, J.; Rossow, L.; Venalainen, E.; Tupasela, T. Effect of vitamin D2- and D3-enriched diets on egg vitamin D content, production, and bird condition during an entire production period. Poult. Sci. 2004, 83, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, C.L.; Jensen, L.S.; Rowland, G.N. Effect of cimetidine on eggshell quality and plasma 25-hydroxycholecalciferol in laying hens. Poult. Sci. 1990, 69, 1892–1899. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.H.; Kerr, J.M.; Gray, R.W. 25-hydroxycholecalciferol in poultry nutrition. Poult. Sci. 1995, 74, 1919–1934. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Turner, B.; Applegate, T.J.; Litta, G.; Kim, W.K. Role of long-term supplementation of 25-hydroxyvitamin D3 on laying hen bone 3-dimensional structural development. Poult. Sci. 2020, 99, 5771–5782. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Wang, Y.; Song, F.; Xu, X.; Liu, M.; Wei, Y.; Zhu, H.; Liu, Y.; Wei, J.; Xu, X. A comparison between vitamin D3 and 25-hydroxyvitamin D3 on laying performance, eggshell quality and ultrastructure, and plasma calcium levels in late period laying hens. Animals 2022, 12, 2824. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Lu, M.; Wang, J.; Zhang, H.J.; Qiu, K.; Qi, G.H.; Wu, S.G. Dietary oregano essential oil supplementation improves intestinal functions and alters gut microbiota in late-phase laying hens. J. Anim. Sci. Biotechnol. 2021, 12, 265–279. [Google Scholar] [CrossRef]
- Gebhardt-Henrich, S.G.; Pfulg, A.; Fröhlich, E.K.F.; Käppeli, S.; Guggisberg, D.; Liesegang, A.; Stoffel, M.H. Limited associations between keel bone damage and bone properties measured with computer tomography, three-point bending test, and analysis of minerals in swiss laying hens. Front. Vet. Sci. 2017, 4, 128. [Google Scholar] [CrossRef]
- Wu, J.; Ma, X.; Liao, X.; Song, C.; Li, S.; Zhang, L.; Lu, L. Dietary calcium and nonphosphate phosphorus interaction influences tibiotarsus development and related gene expression of broilers from 1 to 21 days of age. Poult. Sci. 2023, 102, 102851. [Google Scholar] [CrossRef]
- Dijkslag, M.A.; Kwakkel, R.P.; Martin-Chaves, E.; Alfonso-Carrillo, C.; Navarro-Villa, A. Long-term effects of dietary calcium and phosphorus level, and feed form during rearing on egg production, eggshell quality, and bone traits in brown laying hens from 30 to 89 wk of age. Poult. Sci. 2023, 102, 102618. [Google Scholar] [CrossRef]
- Qiu, J.L.; Zhou, Q.; Zhu, J.M.; Lu, X.T.; Xu, J.M. Organic trace minerals improve eggshell quality by improving the eggshell ultrastructure of laying hens during the late laying period. Poult. Sci. 2019, 99, 1483–1490. [Google Scholar] [CrossRef]
- Akbari Moghaddam Kakhki, R.; Heuthorst, T.; Mills, A.; Neijat, M.; Kiarie, E. Interactive effects of calcium and top-dressed 25-hydroxy vitamin D3 on egg production, egg shell quality, and bones attributes in aged Lohmann LSL-lite layers. Poult. Sci. 2019, 98, 1254–1262. [Google Scholar] [CrossRef]
- Wang, J.; Qiu, L.; Gong, H.; Cel, P.; Yan, L.; Ding, X.; Bai, S.; Zeng, Q.; Mao, X.; Xu, S.; et al. Effect of dietary 25-hydroxycholecalciferol supplementation and high stocking density on performance, egg quality, and tibia quality in laying hens. Poult. Sci. 2020, 99, 2608–2615. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Ding, X.; Bai, S.; Wang, J.; Zeng, Q.; Peng, H.; Xuan, Y.; Zhang, K. Effects of supplementation of 25-hydroxyvitamin D3 as a vitamin D3 substitute on performance, bone traits, and egg quality of laying hens from 1 day to 72 weeks of age. Agriculture 2023, 13, 383. [Google Scholar] [CrossRef]
- Koreleski, J.; Witkiewicz, S. Efficacy of different levels of a cholecalciferol 25-OH-derivative in diets with two limestone forms in laying hen nutrition. J. Anim. Feed Sci. 2005, 14, 299–309. [Google Scholar] [CrossRef]
- Adhikari, R.; White, D.; House, J.D.; Kim, W.K. Effects of additional dosage of vitamin D3, vitamin D2, and 25-hydroxyvitamin D3 on calcium and phosphorus utilization, egg quality and bone mineralization in laying hens. Poult Sci. 2020, 99, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Tilgar, V.; Mnd, R.; Ots, I.; Mgi, M.; Kilgas, P. Calcium availability affects bone growth in nestlings of free-living great tits (Parus major), as detected by plasma alkaline phosphatase. J. Zool. 2004, 263, 269–274. [Google Scholar] [CrossRef]
- Zhang, L.H.; He, T.F.; Hu, J.X.; Li, M.; Piao, X.S. Effects of normal and low calcium and phosphorus levels and 25-hydroxycholecalciferol supplementation on performance, serum antioxidant status, meat quality, and bone properties of broilers. Poult. Sci. 2020, 99, 5663–5672. [Google Scholar] [CrossRef] [PubMed]
- Liermann, W.; Halle, I.; Frahm, J.; Hüther, L.; Weigend, S.; Kühn, J.; Stangl, G.I.; Dänicke, S. Genotype-dependent impact of dietary vitamin D3 on laying hens. Arch. Anim. Nutr. 2023, 77, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Matos, R.D. Calcium metabolism in birds. Vet. Clin. N. Am. Exot. Anim. Pract. 2008, 11, 59–82. [Google Scholar] [CrossRef]
- Safaa, H.M.; Serrano, M.P.; Valencia, D.G.; Frikha, M.; Jiménez-Moreno, E.; Mateos, G.G. Productive performance and egg quality of brown egg-laying hens in the late phase of production as influenced by level and source of calcium in the diet. Poult. Sci. 2008, 87, 2043–2051. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.F.; Chen, Y.P.; Jin, R.; Wang, C.; Wen, C.; Zhou, Y.M. Age-related changes in liver metabolism and antioxidant capacity of laying hens. Poult. Sci. 2021, 100, 101478. [Google Scholar] [CrossRef]
- Olson, K.R.; Gao, Y.; Arif, F.; Arora, K.; Patel, S.; DeLeon, E.R.; Sutton, T.R.; Feelisch, M.; Cortese-Krott, M.M.; Straub, K.D. Metabolism of hydrogen sulfide (H2S) and production of reactive sulfur species (RSS) by superoxide dismutase. Redox Biol. 2018, 15, 74–85. [Google Scholar] [CrossRef]
- Battin, E.E.; Brumaghim, J.L. Antioxidant activity of sulfur and selenium: A review of reactive oxygen species scavenging, glutathione peroxidase, and metal-binding antioxidant mechanisms. Cell. Biochem. Biophys. 2009, 55, 1–23. [Google Scholar] [CrossRef]
- Chou, P.C.; Chen, Y.H.; Chung, T.K.; Walzem, R.L.; Chen, S.E. Supplemental 25-hydroxycholecalciferol alleviates inflammation and cardiac fibrosis in hens. Int. J. Mol. Sci. 2020, 21, 8379. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.; Zhang, T.; Yan, L.; Qiu, L.; Yin, H.; Ding, X.; Bai, S.; Zeng, Q.; Mao, X.; et al. Dietary 25-hydroxyvitamin D improves intestinal health and microbiota of laying hens under high stocking density. Poult. Sci. 2021, 100, 101132. [Google Scholar] [CrossRef] [PubMed]
- Wauquier, F.; Leotoing, L.; Coxam, V.; Guicheux, J.; Wittrant, Y. Oxidative stress in bone remodelling and disease. Trends Mol. Med. 2009, 15, 468–477. [Google Scholar] [CrossRef]
- Altindag, O. Atherosclerosis in patients with osteoporosis. Turk. J. Rheumatol. 2009, 24, 60–61. [Google Scholar]
- Bertero, E.; Maack, C. Calcium signaling and reactive oxygen species in mitochondria. Circ. Res. 2018, 122, 1460–1478. [Google Scholar] [CrossRef]
- Gloux, A.; Le Roy, N.; Brionne, A.; Bonin, E.; Juanchich, A.; Benzoni, G.; Piketty, M.L.; Prié, D.; Nys, Y.; Gautron, J.; et al. Candidate genes of the transcellular and paracellular calcium absorption pathways in the small intestine of laying hens. Poult. Sci. 2019, 98, 6005–6018. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.M.; Wu, S.G.; Qi, G.H.; Fu, Y.; Wang, W.W.; Zhang, H.J.; Wang, J. Dietary supplemental xylooligosaccharide modulates nutrient digestibility, intestinal morphology, and gut microbiota in laying hens. Anim. Nutr. 2021, 7, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Porras, N.; Rebollada-Merino, A.; Bárcena, C.; Mayoral-Alegre, F.J.; Lomillos, J.M.; Domínguez, L.; Rodríguez-Bertos, A. Effect of biscuit flour and fermented defatted “alperujo” co-administration on intestinal mucosa morphology and productive performance in laying hens. Animals 2021, 11, 1075. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, K.; Bai, S.; Wang, J.; Zeng, Q.; Peng, H.; Su, Z.; Xuan, Y.; Qi, S.; Ding, X. Effect of 25-hydroxycholecalciferol with different vitamin D3 levels in the hens diet in the rearing period on growth performance, bone quality, egg production, and eggshell quality. Agriculture 2021, 11, 698. [Google Scholar] [CrossRef]
- Reich, T.; Gefen, A. Effect of trabecular bone loss on cortical strain rate during impact in an in vitro model of avian femur. Biomed. Eng. Online 2006, 5, 45. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Ingredients | Content, % | Nutrient Level 2 | |
|---|---|---|---|
| Corn | 63.65 | CP, % | 16.50 |
| Soybean meal | 25.41 | Metabolizable energy 2, MJ/kg | 11.30 |
| Soybean oil | 0.11 | Calcium, % | 3.50 |
| Stone powder | 9.18 | Total phosphorus, % | 0.49 |
| Calcium hydrophosphate | 0.90 | Available phosphorus, % | 0.29 |
| Salt | 0.30 | Met + Cys, % | 0.65 |
| DL-Methionine | 0.18 | Lysine, % | 0.79 |
| Premix 1 | 0.27 | Threonine, % | 0.56 |
| Total | 100.00 | Tryptophan, % | 0.18 |
| Items | Time/Week | 25-OHD3 Levels, μg/kg | SEM | p-Value | ||
|---|---|---|---|---|---|---|
| 0 | 50 | |||||
| 4000 IU VD3 | 2000 IU VD3 | 4000 IU VD3 | ||||
| Average egg weight, g | 1–4 | 62.04 | 61.99 | 61.88 | 0.20 | 0.95 |
| 5–8 | 61.32 | 61.47 | 61.16 | 0.27 | 0.91 | |
| 9–12 | 61.14 | 61.91 | 62.12 | 0.55 | 0.60 | |
| Average daily feed intake, g | 1–4 | 123.68 | 121.40 | 122.23 | 0.70 | 0.43 |
| 5–8 | 121.24 | 124.34 | 119.44 | 0.98 | 0.11 | |
| 9–12 | 125.35 | 121.78 | 119.78 | 2.22 | 0.61 | |
| Feed-to-egg ratio | 1–4 | 2.09 | 2.13 | 2.07 | 0.02 | 0.54 |
| 5–8 | 2.12 | 2.24 | 2.07 | 0.03 | 0.08 | |
| 9–12 | 2.21 | 2.06 | 2.17 | 0.03 | 0.11 | |
| Laying rate, % | 1–4 | 87.88 | 87.58 | 90.79 | 0.62 | 0.05 |
| 5–8 | 86.63 b | 89.54 a | 87.88 ab | 0.19 | 0.01 | |
| 9–12 | 78.19 b | 84.76 a | 80.47 b | 0.83 | <0.01 | |
| Items | Time/Week | 25-OHD3 Levels, μg/kg | SEM | p-Value | ||
|---|---|---|---|---|---|---|
| 0 | 50 | |||||
| 4000 IU VD3 | 2000 IU VD3 | 4000 IU VD3 | ||||
| Egg weight, g | 4 | 64.43 | 64.39 | 62.87 | 0.55 | 0.54 |
| 8 | 61.03 | 61.56 | 61.18 | 0.32 | 0.80 | |
| 12 | 62.09 | 62.64 | 62.03 | 0.54 | 0.89 | |
| Egg yolk weight, g | 4 | 17.02 | 17.04 | 16.97 | 0.12 | 0.97 |
| 8 | 16.40 | 16.48 | 16.26 | 0.18 | 0.90 | |
| 12 | 16.24 | 15.88 | 16.12 | 0.13 | 0.53 | |
| Egg yolk ratio,% | 4 | 26.41 | 26.50 | 26.47 | 0.30 | 0.99 |
| 8 | 26.86 | 26.76 | 26.58 | 0.28 | 0.93 | |
| 12 | 25.86 | 25.77 | 25.95 | 0.21 | 0.94 | |
| Albumen weight, g | 4 | 41.11 | 41.13 | 40.11 | 0.49 | 0.65 |
| 8 | 38.96 | 39.39 | 39.11 | 0.28 | 0.83 | |
| 12 | 40.39 | 40.48 | 39.93 | 0.42 | 0.86 | |
| Albumen ratio,% | 4 | 63.82 | 63.84 | 63.77 | 0.31 | 1.00 |
| 8 | 63.84 | 63.99 | 63.92 | 0.33 | 0.98 | |
| 12 | 65.04 | 64.63 | 64.35 | 0.23 | 0.51 | |
| Egg shape index | 4 | 1.23 | 1.24 | 1.24 | 0.005 | 0.31 |
| 8 | 1.31 | 1.32 | 1.32 | 0.005 | 0.99 | |
| 12 | 1.30 | 1.29 | 1.28 | 0.005 | 0.46 | |
| Eggshell strength, N | 4 | 37.60 | 39.32 | 40.00 | 0.45 | 0.07 |
| 8 | 33.32 b | 33.19 b | 36.66 a | 0.44 | <0.01 | |
| 12 | 33.36 b | 35.14 a | 34.21 ab | 0.28 | 0.02 | |
| Thick albumen height, mm | 4 | 6.44 b | 6.99 a | 7.09 a | 0.09 | <0.01 |
| 8 | 6.16 b | 7.07 a | 7.07 a | 0.12 | <0.01 | |
| 12 | 6.42 b | 7.50 a | 7.45 a | 0.14 | <0.01 | |
| Haugh unit | 4 | 75.89 b | 81.33 a | 82.37 a | 0.73 | <0.01 |
| 8 | 77.02 b | 83.53 a | 83.30 a | 0.85 | <0.01 | |
| 12 | 79.17 b | 85.32 a | 84.77 a | 0.74 | <0.01 | |
| Items | 25-OHD3 Levels, μg/kg | SEM | p-Value | ||
|---|---|---|---|---|---|
| 0 | 50 | ||||
| 4000 IU VD3 | 2000 IU VD3 | 4000 IU VD3 | |||
| ALT, U/L | 36.32 b | 41.51 a | 37.17 b | 0.77 | <0.01 |
| AST, U/L | 74.12 | 75.67 | 73.64 | 0.76 | 0.56 |
| ALP, U/L | 331.83 a | 298.09 b | 291.82 b | 5.42 | <0.01 |
| ALB, g/L | 24.32 b | 30.85 a | 29.51 a | 0.88 | <0.01 |
| UA, umol/L | 252.31 | 241.99 | 252.09 | 3.73 | 0.46 |
| CRE, umol/L | 73.89 | 77.16 | 74.43 | 0.78 | 0.20 |
| GLU, mmol/L | 12.52 | 12.33 | 13.10 | 0.24 | 0.42 |
| T-BIL, umol/L | 32.69 a | 26.94 b | 30.07 ab | 0.76 | <0.01 |
| Ca, mmolL | 2.45 b | 2.64 ab | 2.75 a | 0.05 | 0.02 |
| P, mmol/L | 2.40 b | 2.84 a | 2.73 a | 0.05 | <0.01 |
| Items | 25-OHD3 Levels, μg/kg | SEM | p-Value | ||
|---|---|---|---|---|---|
| 0 | 50 | ||||
| 4000 IU VD3 | 2000 IU VD3 | 4000 IU VD3 | |||
| 25-OHD3, ng/mL | 5.34 b | 5.81 a | 5.55 ab | 0.08 | 0.02 |
| FSH, IU/L | 10.09 c | 16.31 a | 13.16 b | 0.65 | <0.01 |
| T, nmol/L | 1.14 b | 1.39 a | 1.47 a | 0.04 | <0.01 |
| Progesterone, nmol/L | 8.01 b | 9.04 a | 8.07 b | 0.15 | <0.01 |
| CA, ng/mL | 5.21 | 5.60 | 5.40 | 0.07 | 0.06 |
| COR, ng/mL | 26.81 a | 18.25 b | 10.20 c | 1.65 | <0.01 |
| DAO, ng/mL | 7.79 | 7.51 | 7.26 | 0.10 | 0.10 |
| ET, ng/mL | 11.30 a | 11.37 a | 9.75 b | 0.22 | <0.01 |
| PTH, pg/mL | 16.88 ab | 17.26 a | 15.09 b | 0.38 | <0.01 |
| BGP, ng/mL | 1.50 a | 1.29 b | 1.20 b | 0.04 | <0.01 |
| CT, pg/mL | 4.28 a | 4.04 b | 3.95 b | 0.04 | <0.01 |
| Items | 25-OHD3 Levels, μg/kg | SEM | p-Value | ||
|---|---|---|---|---|---|
| 0 | 50 | ||||
| 4000 IU VD3 | 2000 IU VD3 | 4000 IU VD3 | |||
| GSH-Px, U/mL | 519.48 c | 893.23 a | 814.94 b | 39.37 | <0.01 |
| CAT, U/L | 5.46 c | 19.95 a | 11.86 b | 1.45 | <0.01 |
| SOD, U/mL | 14.92 b | 17.81 a | 15.21 b | 0.41 | 0.01 |
| MDA, nmol/mL | 21.62 a | 8.24 b | 7.90 b | 1.57 | <0.01 |
| Hydroxyl radical scavenging ability, U/mL | 238.15 b | 248.32 a | 255.02 a | 2.05 | <0.01 |
| Superoxide anion scavenging ability, U/mL | 435.51 b | 527.77 a | 516.36 a | 10.42 | <0.01 |
| T-AOC, U/mL | 18.99 | 20.49 | 21.57 | 0.42 | 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Qiu, K.; Zheng, J.; Zhang, H.; Wang, J.; Qi, X.; Wu, S. Dietary 25-Hydroxycholecalciferol Supplementation as a Vitamin D3 Substitute Improves Performance, Egg Quality, Blood Indexes, Jejunal Morphology, and Tibia Quality in Late-Phase Laying Hens. Animals 2024, 14, 878. https://doi.org/10.3390/ani14060878
Gao S, Qiu K, Zheng J, Zhang H, Wang J, Qi X, Wu S. Dietary 25-Hydroxycholecalciferol Supplementation as a Vitamin D3 Substitute Improves Performance, Egg Quality, Blood Indexes, Jejunal Morphology, and Tibia Quality in Late-Phase Laying Hens. Animals. 2024; 14(6):878. https://doi.org/10.3390/ani14060878
Chicago/Turabian StyleGao, Shan, Kai Qiu, Junjie Zheng, Haijun Zhang, Jing Wang, Xiaolong Qi, and Shugeng Wu. 2024. "Dietary 25-Hydroxycholecalciferol Supplementation as a Vitamin D3 Substitute Improves Performance, Egg Quality, Blood Indexes, Jejunal Morphology, and Tibia Quality in Late-Phase Laying Hens" Animals 14, no. 6: 878. https://doi.org/10.3390/ani14060878
APA StyleGao, S., Qiu, K., Zheng, J., Zhang, H., Wang, J., Qi, X., & Wu, S. (2024). Dietary 25-Hydroxycholecalciferol Supplementation as a Vitamin D3 Substitute Improves Performance, Egg Quality, Blood Indexes, Jejunal Morphology, and Tibia Quality in Late-Phase Laying Hens. Animals, 14(6), 878. https://doi.org/10.3390/ani14060878