
Effects of Succinate on Growth Performance, Meat Quality and Lipid Synthesis in Bama Miniature Pigs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Treatments
2.2. Cell Culture and Treatments
2.3. Sample Collection
2.4. Meat Quality Analysis
2.5. Serum Biochemical Indices
2.6. Histological Analysis
2.7. Succinate Determination
2.8. BODIPY Dyeing
2.9. Metabolomics Analysis
2.10. Quantitative Real-Time PCR Analysis
2.11. Immunoblotting Analyses
2.12. Statistical Analysis
3. Results
3.1. Sodium Succinate Improves the Growth and Meat Quality of Pigs
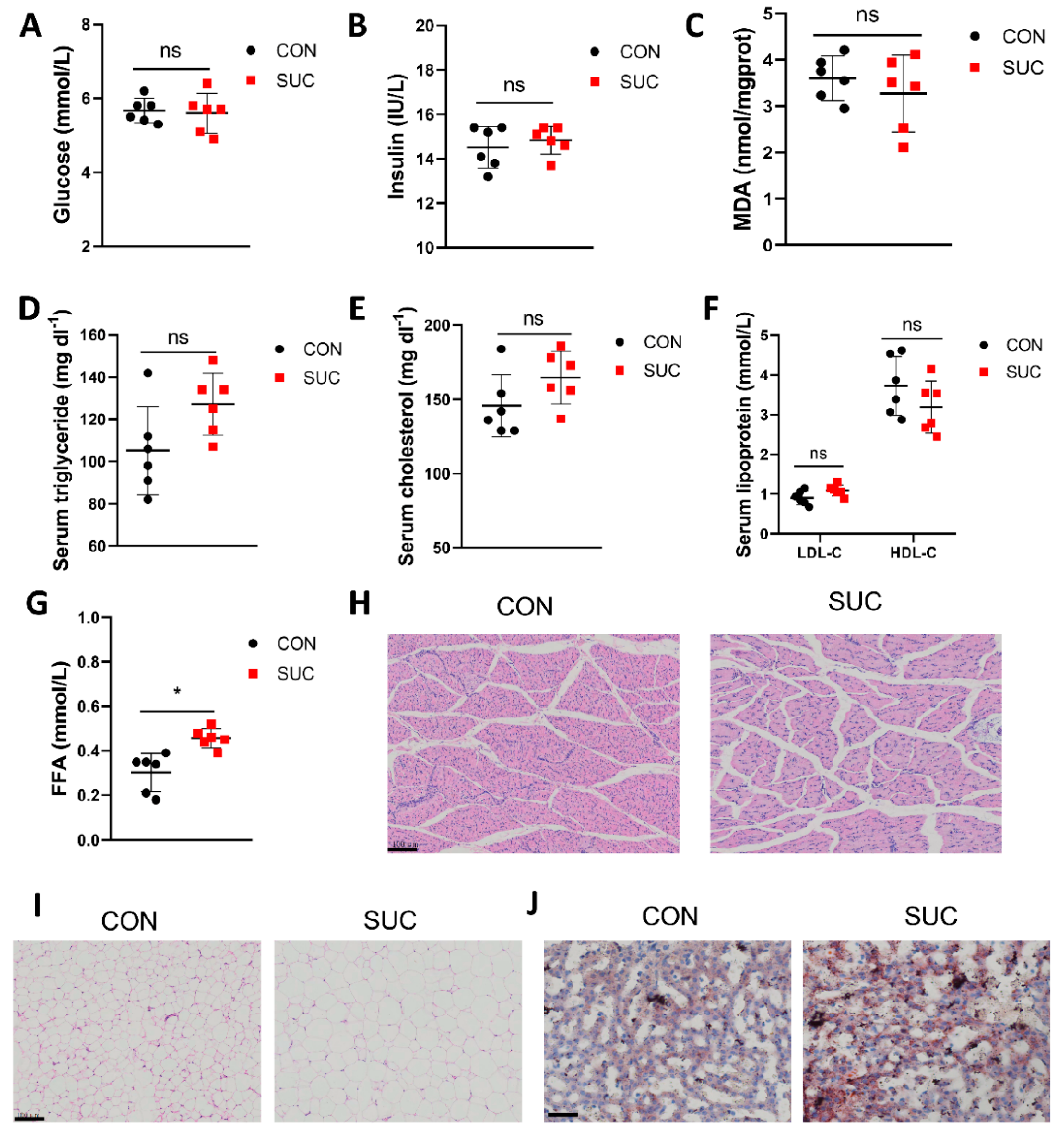
3.2. Sodium Succinate Promotes Lipid Accumulation in Pig Serum, Adipose Tissue, and Liver
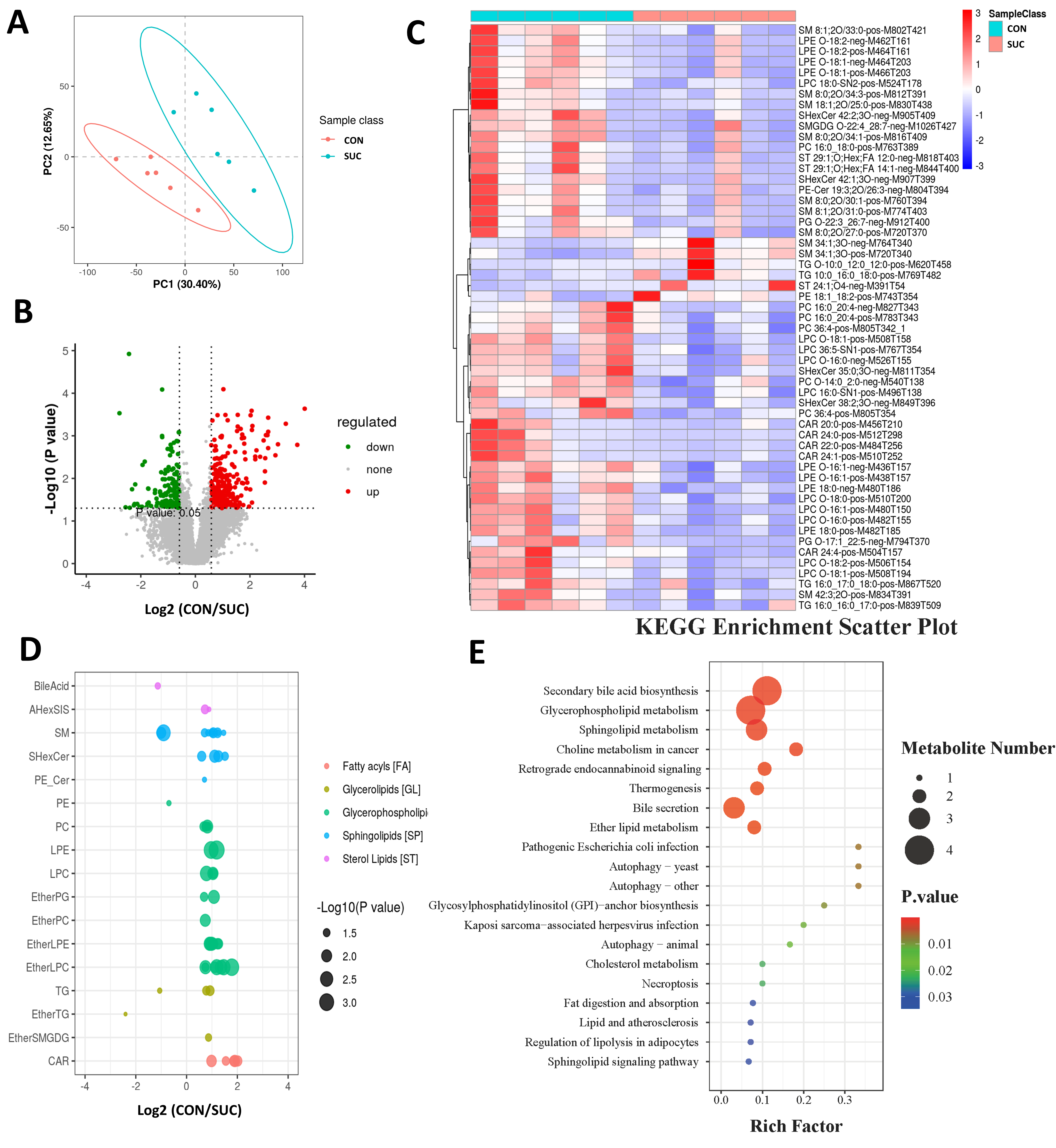
3.3. Sodium Succinate Regulates the Lipid Composition of LM
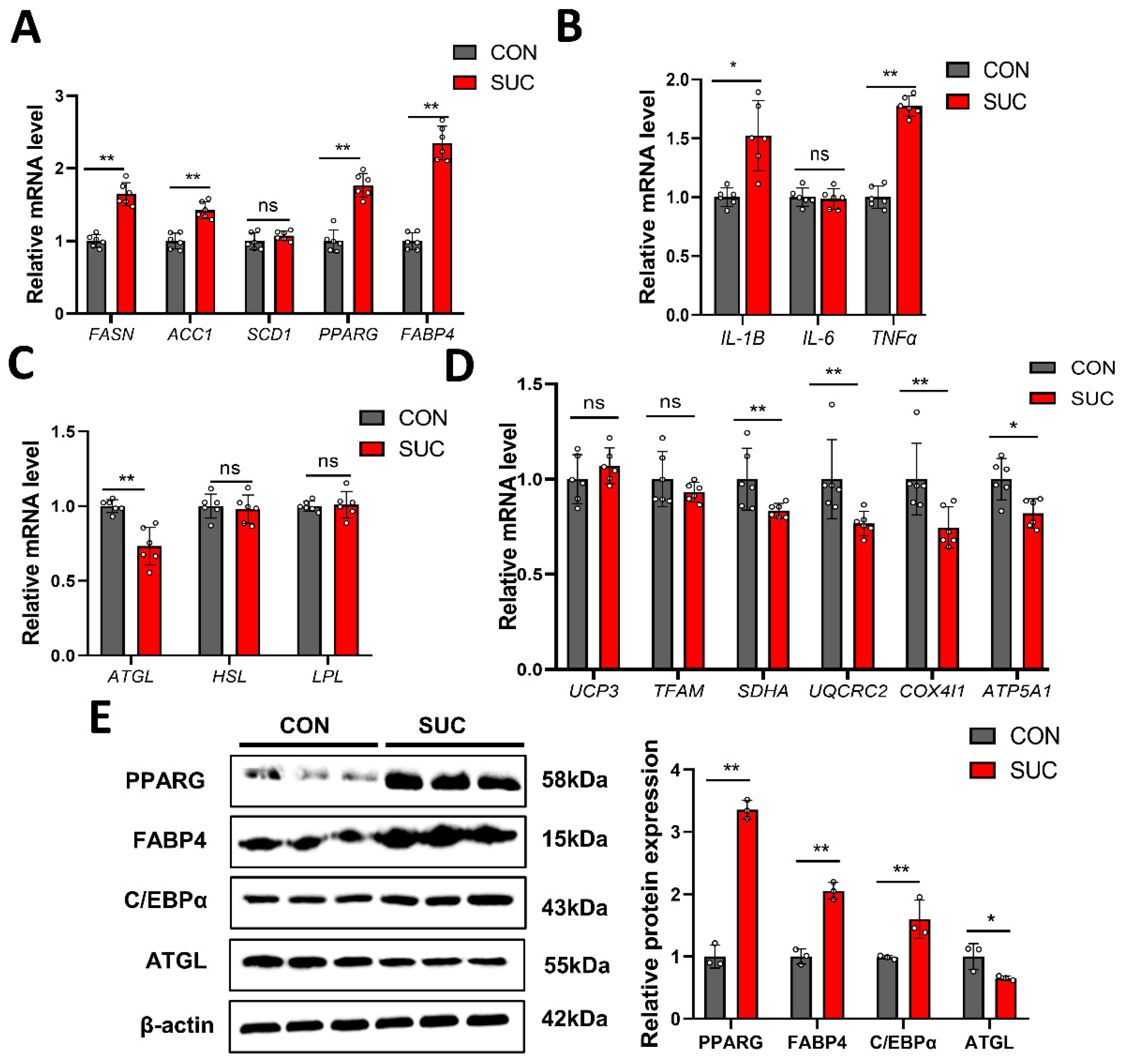
3.4. Succinate Regulates the Expression of Genes Related to Lipid Metabolism
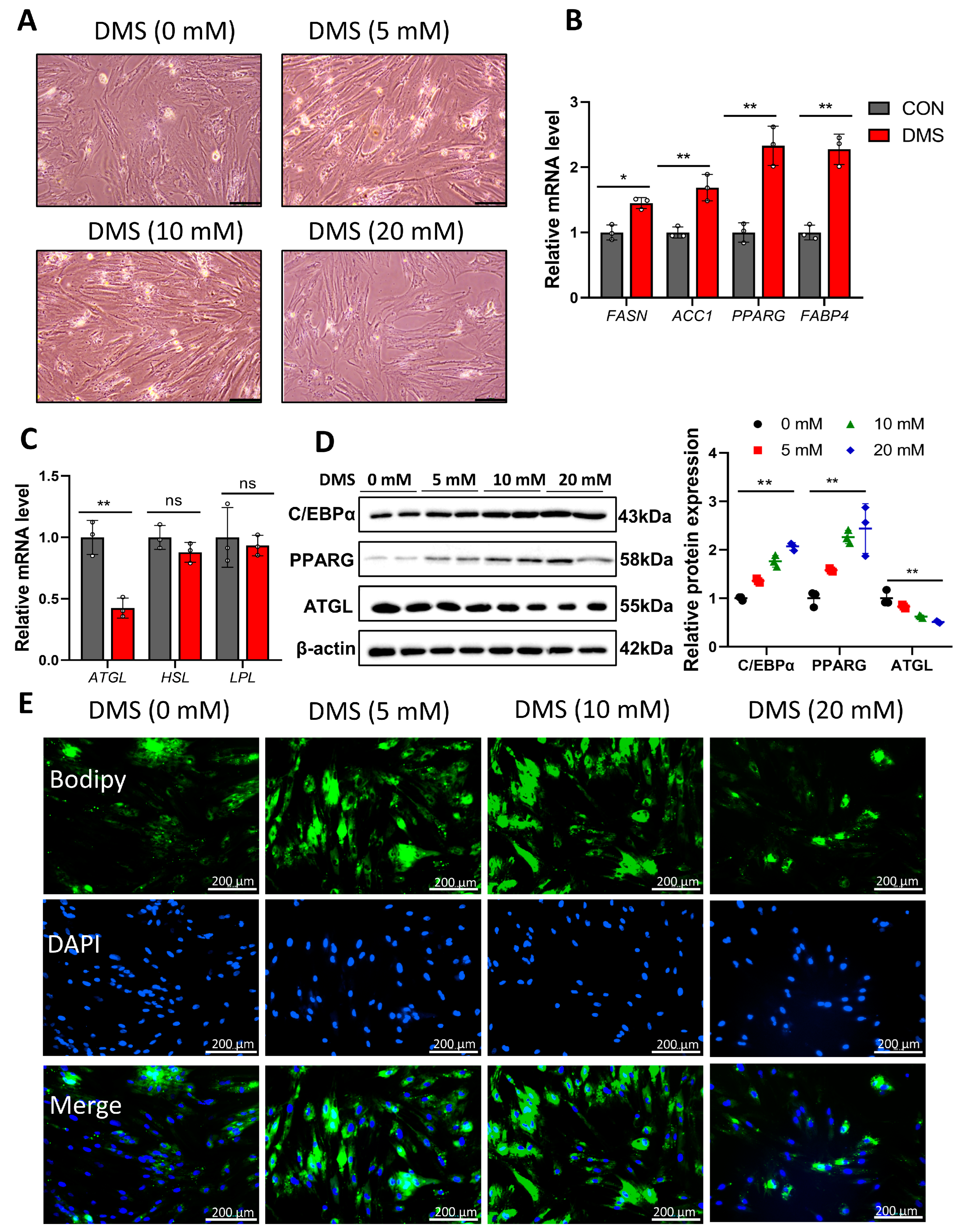
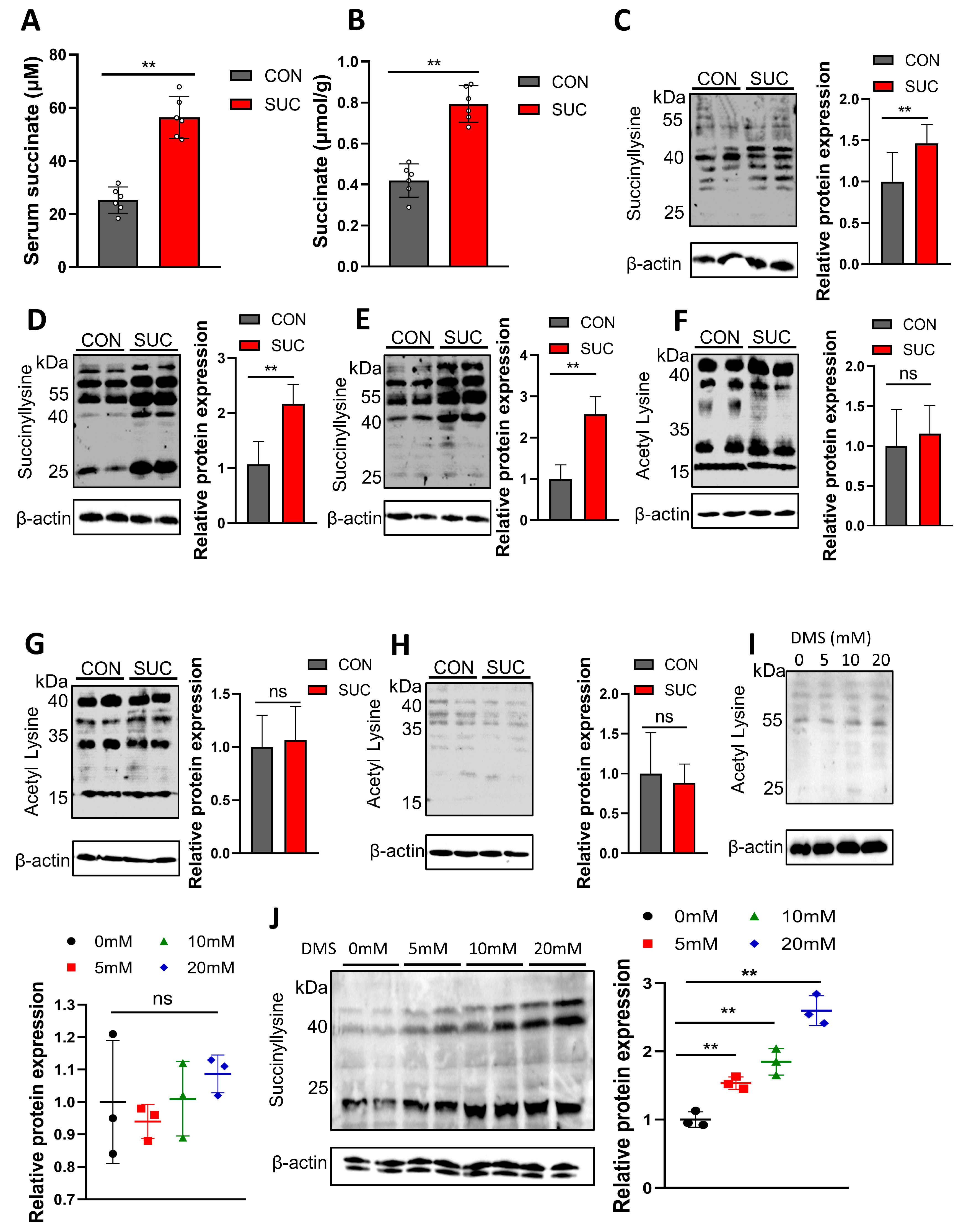
3.5. Succinate Promotes Lipid Deposition by Regulating Succinylation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, E.; Guo, J.; Yin, J. Nutritional regulation of skeletal muscle energy metabolism, lipid accumulation and meat quality in pigs. Anim. Nutr. 2023, 14, 185–192. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, Y.; Zhang, L.; Kong, X.; Li, F. Serine-to-glycine ratios in low-protein diets regulate intramuscular fat by affecting lipid metabolism and myofiber type transition in the skeletal muscle of growing-finishing pigs. Anim. Nutr. 2021, 7, 384–392. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, Y.; Wu, J.; Qiao, M.; Xu, Z.; Peng, X.; Mei, S. Proteomic and lipidomic analyses reveal saturated fatty acids, phosphatidylinositol, phosphatidylserine, and associated proteins contributing to intramuscular fat deposition. J. Proteom. 2021, 241, 104235. [Google Scholar] [CrossRef]
- Zomeño, C.; Gispert, M.; Čandek-Potokar, M.; Mörlein, D.; Font, I.F.M. A matter of body weight and sex type: Pig carcass chemical composition and pork quality. Meat Sci. 2023, 197, 109077. [Google Scholar] [CrossRef]
- Sarmiento-García, A.; Vieira-Aller, C. Improving Fatty Acid Profile in Native Breed Pigs Using Dietary Strategies: A Review. Animals 2023, 13, 1696. [Google Scholar] [CrossRef]
- Mo, J.; Lu, Y.; Xing, T.; Xu, D.; Zhang, K.; Zhang, S.; Wang, Y.; Yan, G.; Lan, G.; Liang, J. Blood metabolic and physiological profiles of Bama miniature pigs at different growth stages. Porc. Health Manag. 2022, 8, 35. [Google Scholar] [CrossRef]
- De Vadder, F.; Kovatcheva-Datchary, P.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-Produced Succinate Improves Glucose Homeostasis via Intestinal Gluconeogenesis. Cell Metab. 2016, 24, 151–157. [Google Scholar] [CrossRef]
- Li, X.; Ren, Y.; Huang, G.; Zhang, R.; Zhang, Y.; Zhu, W.; Yu, K. Succinate communicates pro-inflammatory signals to the host and regulates bile acid enterohepatic metabolism in a pig model. Food Funct. 2022, 13, 11070–11082. [Google Scholar] [CrossRef]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235.e5. [Google Scholar] [CrossRef]
- Mills, E.; O’Neill, L.A. Succinate: A metabolic signal in inflammation. Trends Cell Biol. 2014, 24, 313–320. [Google Scholar] [CrossRef]
- Tannahill, G.M.; Curtis, A.M.; Adamik, J.; Palsson-McDermott, E.M.; McGettrick, A.F.; Goel, G.; Frezza, C.; Bernard, N.J.; Kelly, B.; Foley, N.H.; et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature 2013, 496, 238–242. [Google Scholar] [CrossRef]
- Zhou, X.; Liu, Y.; Xiong, X.; Chen, J.; Tang, W.; He, L.; Zhang, Z.; Yin, Y.; Li, F. Intestinal accumulation of microbiota-produced succinate caused by loss of microRNAs leads to diarrhea in weanling piglets. Gut Microbes 2022, 14, 2091369. [Google Scholar] [CrossRef]
- Qiu, K.; Li, C.L.; Wang, J.; Qi, G.H.; Gao, J.; Zhang, H.J.; Wu, S.G. Effects of Dietary Supplementation With Bacillus subtilis, as an Alternative to Antibiotics, on Growth Performance, Serum Immunity, and Intestinal Health in Broiler Chickens. Front. Nutr. 2021, 8, 786878. [Google Scholar] [CrossRef]
- Zhou, Z.; Yu, L.; Cao, J.; Yu, J.; Lin, Z.; Hong, Y.; Jiang, S.; Chen, C.; Mi, Y.; Zhang, C.; et al. Lactobacillus salivarius Promotion of Intestinal Stem Cell Activity in Hens Is Associated with Succinate-Induced Mitochondrial Energy Metabolism. mSystems 2022, 7, e0090322. [Google Scholar] [CrossRef]
- Yuan, Y.; Xu, Y.; Xu, J.; Liang, B.; Cai, X.; Zhu, C.; Wang, L.; Wang, S.; Zhu, X.; Gao, P.; et al. Succinate promotes skeletal muscle protein synthesis via Erk1/2 signaling pathway. Mol. Med. Rep. 2017, 16, 7361–7366. [Google Scholar] [CrossRef]
- Liu, K.; Lin, L.; Li, Q.; Xue, Y.; Zheng, F.; Wang, G.; Zheng, C.; Du, L.; Hu, M.; Huang, Y.; et al. Scd1 controls de novo beige fat biogenesis through succinate-dependent regulation of mitochondrial complex II. Proc. Natl. Acad. Sci. USA 2020, 117, 2462–2472. [Google Scholar] [CrossRef]
- Zhao, G.; Zhen, J.; Liu, X.; Guo, J.; Li, D.; Xie, J.; Xie, L. Protein post-translational modification by lysine succinylation: Biochemistry, biological implications, and therapeutic opportunities. Genes Dis. 2023, 10, 1242–1262. [Google Scholar] [CrossRef]
- Sreedhar, A.; Wiese, E.K.; Hitosugi, T. Enzymatic and metabolic regulation of lysine succinylation. Genes Dis. 2020, 7, 166–171. [Google Scholar] [CrossRef]
- Wang, G.; Meyer, J.G.; Cai, W.; Softic, S.; Li, M.E.; Verdin, E.; Newgard, C.; Schilling, B.; Kahn, C.R. Regulation of UCP1 and Mitochondrial Metabolism in Brown Adipose Tissue by Reversible Succinylation. Mol. Cell 2019, 74, 844–857.e7. [Google Scholar] [CrossRef]
- Ding, Q.; Lu, C.; Hao, Q.; Zhang, Q.; Yang, Y.; Olsen, R.E.; Ringo, E.; Ran, C.; Zhang, Z.; Zhou, Z. Dietary Succinate Impacts the Nutritional Metabolism, Protein Succinylation and Gut Microbiota of Zebrafish. Front. Nutr. 2022, 9, 894278. [Google Scholar] [CrossRef]
- Honikel, K.O. Reference methods for the assessment of physical characteristics of meat. Meat Sci. 1998, 49, 447–457. [Google Scholar] [CrossRef]
- Yu, M.; Li, Z.; Chen, W.; Rong, T.; Wang, G.; Li, J.; Ma, X. Use of Hermetia illucens larvae as a dietary protein source: Effects on growth performance, carcass traits, and meat quality in finishing pigs. Meat Sci. 2019, 158, 107837. [Google Scholar] [CrossRef]
- Wei, D.; Tian, X.; Zhu, L.; Wang, H.; Sun, C. USP14 governs CYP2E1 to promote nonalcoholic fatty liver disease through deubiquitination and stabilization of HSP90AA1. Cell Death Dis. 2023, 14, 566. [Google Scholar] [CrossRef]
- Arnold, P.K.; Finley, L.W.S. Regulation and function of the mammalian tricarboxylic acid cycle. J. Biol. Chem. 2023, 299, 102838. [Google Scholar] [CrossRef]
- Akram, M. Citric acid cycle and role of its intermediates in metabolism. Cell Biochem. Biophys. 2014, 68, 475–478. [Google Scholar] [CrossRef]
- Niehaus, T.D.; Hillmann, K.B. Enzyme promiscuity, metabolite damage, and metabolite damage control systems of the tricarboxylic acid cycle. FEBS J. 2020, 287, 1343–1358. [Google Scholar] [CrossRef]
- Connors, J.; Dawe, N.; Van Limbergen, J. The Role of Succinate in the Regulation of Intestinal Inflammation. Nutrients 2018, 11, 25. [Google Scholar] [CrossRef]
- Mills, E.L.; Pierce, K.A.; Jedrychowski, M.P.; Garrity, R.; Winther, S.; Vidoni, S.; Yoneshiro, T.; Spinelli, J.B.; Lu, G.Z.; Kazak, L.; et al. Accumulation of succinate controls activation of adipose tissue thermogenesis. Nature 2018, 560, 102–106. [Google Scholar] [CrossRef]
- Fernandez, X.; Monin, G.; Talmant, A.; Mourot, J.; Lebret, B. Influence of intramuscular fat content on the quality of pig meat—2. Consumer acceptability of m. longissimus lumborum. Meat Sci. 1999, 53, 67–72. [Google Scholar] [CrossRef]
- Wang, T.; Xu, Y.Q.; Yuan, Y.X.; Xu, P.W.; Zhang, C.; Li, F.; Wang, L.N.; Yin, C.; Zhang, L.; Cai, X.C.; et al. Succinate induces skeletal muscle fiber remodeling via SUNCR1 signaling. EMBO Rep. 2019, 20, e47892. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.; Bozi, L.H.M.; Yaghi, O.K.; Mills, E.L.; Xiao, H.; Nicholson, H.E.; Paschini, M.; Paulo, J.A.; Garrity, R.; Laznik-Bogoslavski, D.; et al. pH-Gated Succinate Secretion Regulates Muscle Remodeling in Response to Exercise. Cell 2020, 183, 62–75.e17. [Google Scholar] [CrossRef]
- Gray, L.R.; Tompkins, S.C.; Taylor, E.B. Regulation of pyruvate metabolism and human disease. Cell. Mol. Life Sci. 2014, 71, 2577–2604. [Google Scholar] [CrossRef]
- Onodera, T.; Wang, M.Y.; Rutkowski, J.M.; Deja, S.; Chen, S.; Balzer, M.S.; Kim, D.S.; Sun, X.; An, Y.A.; Field, B.C.; et al. Endogenous renal adiponectin drives gluconeogenesis through enhancing pyruvate and fatty acid utilization. Nat. Commun. 2023, 14, 6531. [Google Scholar] [CrossRef]
- Nowinski, S.M.; Solmonson, A.; Rusin, S.F.; Maschek, J.A.; Bensard, C.L.; Fogarty, S.; Jeong, M.Y.; Lettlova, S.; Berg, J.A.; Morgan, J.T.; et al. Mitochondrial fatty acid synthesis coordinates oxidative metabolism in mammalian mitochondria. eLife 2020, 9, e58041. [Google Scholar] [CrossRef] [PubMed]
- Majithia, A.R.; Flannick, J.; Shahinian, P.; Guo, M.; Bray, M.A.; Fontanillas, P.; Gabriel, S.B.; Rosen, E.D.; Altshuler, D. Rare variants in PPARG with decreased activity in adipocyte differentiation are associated with increased risk of type 2 diabetes. Proc. Natl. Acad. Sci. USA 2014, 111, 13127–13132. [Google Scholar] [CrossRef] [PubMed]
- Lefterova, M.I.; Zhang, Y.; Steger, D.J.; Schupp, M.; Schug, J.; Cristancho, A.; Feng, D.; Zhuo, D.; Stoeckert, C.J., Jr.; Liu, X.S.; et al. PPARgamma and C/EBP factors orchestrate adipocyte biology via adjacent binding on a genome-wide scale. Genes Dev. 2008, 22, 2941–2952. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tan, M.; Xie, Z.; Dai, L.; Chen, Y.; Zhao, Y. Identification of lysine succinylation as a new post-translational modification. Nat. Chem. Biol. 2011, 7, 58–63. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Y.; Zhao, L.; Tian, Q.; deAvila, J.M.; Zhu, M.J.; Du, M. Dietary succinate supplementation to maternal mice improves fetal brown adipose tissue development and thermogenesis of female offspring. J. Nutr. Biochem. 2022, 100, 108908. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Percentage (%) | Nutrient Levels | Content |
|---|---|---|---|
| Corn | 65.76 | DE, Mcal/kg | 3.40 |
| Soybean meal | 18.60 | CP, % | 16.65 |
| Soybean oil | 2 | ||
| Whey powder | 8 | ||
| Fish meal | 2 | ||
| Lysine | 0.43 | ||
| Methionine | 0.08 | ||
| Threonine | 0.13 | ||
| Tryptophan | 0.02 | ||
| CaHPO4 | 1.05 | ||
| Calcium carbonate | 0.68 | ||
| Salt | 0.25 | ||
| Premix a | 1 | ||
| Total | 100 |
| Genes | Primers | Sequences (5′ to 3′) |
|---|---|---|
| FASN | Forward | TGGGCATGGTGAACTGTCTC |
| Reverse | GCGTGGTTGTTGGAAAGGTC | |
| ACC1 | Forward | GGCCATCAAGGACTTCAACC |
| Reverse | ACGATGTAAGCGCCGAACTT | |
| SCD1 | Forward | TCTGGGCGTTTGCCTACTATCT |
| Reverse | TCTTTGACGGCTGGGTGTTT | |
| PPARG | Forward | AACATTTCACAAGAGGTGACCA |
| Reverse | GATCTCGTGGACGCCATACT | |
| FABP4 | Forward | CAGGAAAGTCAAGAGCACCACAGGAAAGTCAAGA |
| Reverse | TCGGGACAATACATCCTCCAACA | |
| ATGL | Forward | ATGTTCCCCAAAGAGACGAC |
| Reverse | GGCGAAGCGGGTTATGAT | |
| HSL | Forward | CACAAGGGCTGCTTCTACGG |
| Reverse | AAGCGGCCACTGGTGAAGAG | |
| LPL | Forward | CGTGCTCAGATGCCCTACAA |
| Reverse | AGACTCCACGTGCTGTTCCT | |
| IL-1B | Forward | AAACCTTGACCTCAGCCCTC |
| Reverse | CTCCTCCTTTGCCACAATCAC | |
| IL-6 | Forward | CTTCAGTCCAGTCGCCTTCT |
| Reverse | CATCACCTTTGGCATCTTCTT | |
| TNF-α | Forward | CCACCAACGTTTTCCTCACT |
| Reverse | AATAAAGGGATGGACAGGGG | |
| UCP3 | Forward | TCACCTTCAGGACACGTTCG |
| Reverse | AGGCATCCATCCTAGTGGGT | |
| TFAM | Forward | TGCTTTGTCTACGGGTGCAA |
| Reverse | GCAAAACTGAACGGAGAGCG | |
| SDHA | Forward | ACATCAACGGAGGCAACACT |
| Reverse | ACAGCCCATCCAGTTTCTCG | |
| UQCRC2 | Forward | CTCCTGTAAGGCGGTTGTGA |
| Reverse | ACTGGATGCAAGACGAAGCA | |
| COX4I1 | Forward | GGTGGAGTCCCCTCTCGAT |
| Reverse | GGATGGGGCCGTACACATAG | |
| ATP5A1 | Forward | TCGTGGTGTTCGTCTGACTG |
| Reverse | TTTTCCCAACAGGGCTTGGT | |
| β-Actin | Forward | AACGGCTCCGGCATGTGCAA |
| Reverse | CTTCTGACCCATGCCCACCA |
| Items | CON | SUC | p-Value |
|---|---|---|---|
| Body weight (kg) | |||
| 30 d | 4.397 ± 0.170 | 4.398 ± 0.199 | 0.988 |
| 60 d | 9.110 ± 0.201 | 9.227 ± 0.196 | 0.333 |
| 90 d | 14.111 ± 0.249 | 14.555 ± 0.353 | 0.031 |
| 120 d | 22.452 ± 0.641 | 24.097 ± 0.679 | 0.002 |
| Average daily gain (kg/d) | |||
| 31–60 d | 0.157 ± 0.001 | 0.161 ± 0.001 | 0.505 |
| 61–90 d | 0.167 ± 0.008 | 0.178 ± 0.007 | 0.038 |
| 91–120 d | 0.278 ± 0.026 | 0.318 ± 0.030 | 0.032 |
| Average daily feed intake (kg/d) | |||
| 31–60 d | 0.432 ± 0.021 | 0.435 ± 0.029 | 0.824 |
| 61–90 d | 0.624 ± 0.016 | 0.630 ± 0.027 | 0.667 |
| 91–120 d | 0.924 ± 0.039 | 0.935 ± 0.042 | 0.649 |
| Feed/gain ratio | |||
| 31–60 d | 2.763 ± 0.290 | 2.714 ± 0.277 | 0.772 |
| 61–90 d | 3.750 ± 0.146 | 3.551 ± 0.186 | 0.066 |
| 91–120 d | 3.339 ± 0.244 | 2.956 ± 0.257 | 0.024 |
| Items | CON | SUC | p-Value |
|---|---|---|---|
| Carcass weight (kg) | 13.62 ± 0.55 | 13.88 ± 0.41 | 0.38 |
| Dressing percentage (%) | 65.70 ± 1.39 | 66.13 ± 0.90 | 0.54 |
| Backfat depth (mm) | 24.69 ± 0.66 | 26.33 ± 1.06 | 0.01 |
| Eye muscle area (cm2) | 7.17 ± 0.44 | 7.87 ± 0.26 | 0.01 |
| Total muscle (%) | 22.52 ± 1.14 | 22.24 ± 1.36 | 0.72 |
| Total fat (%) | 18.34 ± 0.73 | 20.13 ± 0.71 | 0.00 |
| Total bone (%) | 9.26 ± 0.37 | 8.95 ± 0.36 | 0.20 |
| Items | CON | SUC | p-Value |
|---|---|---|---|
| PH 45 min | 6.27 ± 0.10 | 6.25 ± 0.10 | 0.79 |
| PH 24 h | 5.28 ± 0.12 | 5.27 ± 0.10 | 0.80 |
| Meat color score | 2.5 ± 0.55 | 2.33 ± 0.52 | 0.60 |
| Drip loss (%) | 3.55 ± 0.19 | 3.47 ± 0.19 | 0.46 |
| Cooking yield (%) | 65.35 ± 1.54 | 64.93 ± 1.27 | 0.61 |
| Shear force (N) | 56.03 ± 1.67 | 51.80 ± 2.05 | 0.00 |
| Marbling score | 1.33 ± 0.52 | 2.33 ± 0.52 | 0.01 |
| Intramuscular fat content (%) | 2.16 ± 0.24 | 2.86 ± 0.30 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, X.; Dang, L.; Wang, S.; Li, W.; Sun, C. Effects of Succinate on Growth Performance, Meat Quality and Lipid Synthesis in Bama Miniature Pigs. Animals 2024, 14, 999. https://doi.org/10.3390/ani14070999
Zhai X, Dang L, Wang S, Li W, Sun C. Effects of Succinate on Growth Performance, Meat Quality and Lipid Synthesis in Bama Miniature Pigs. Animals. 2024; 14(7):999. https://doi.org/10.3390/ani14070999
Chicago/Turabian StyleZhai, Xiangyun, Liping Dang, Shiyu Wang, Wenyuan Li, and Chao Sun. 2024. "Effects of Succinate on Growth Performance, Meat Quality and Lipid Synthesis in Bama Miniature Pigs" Animals 14, no. 7: 999. https://doi.org/10.3390/ani14070999
APA StyleZhai, X., Dang, L., Wang, S., Li, W., & Sun, C. (2024). Effects of Succinate on Growth Performance, Meat Quality and Lipid Synthesis in Bama Miniature Pigs. Animals, 14(7), 999. https://doi.org/10.3390/ani14070999