
Early Ontogeny of Cichlids Using Selected Species as Examples
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials
2.1. Collection of Spawners
2.2. Acclimatization and Preparation of Fish for Spawning
2.3. Experimental Procedure
3. Methods Used
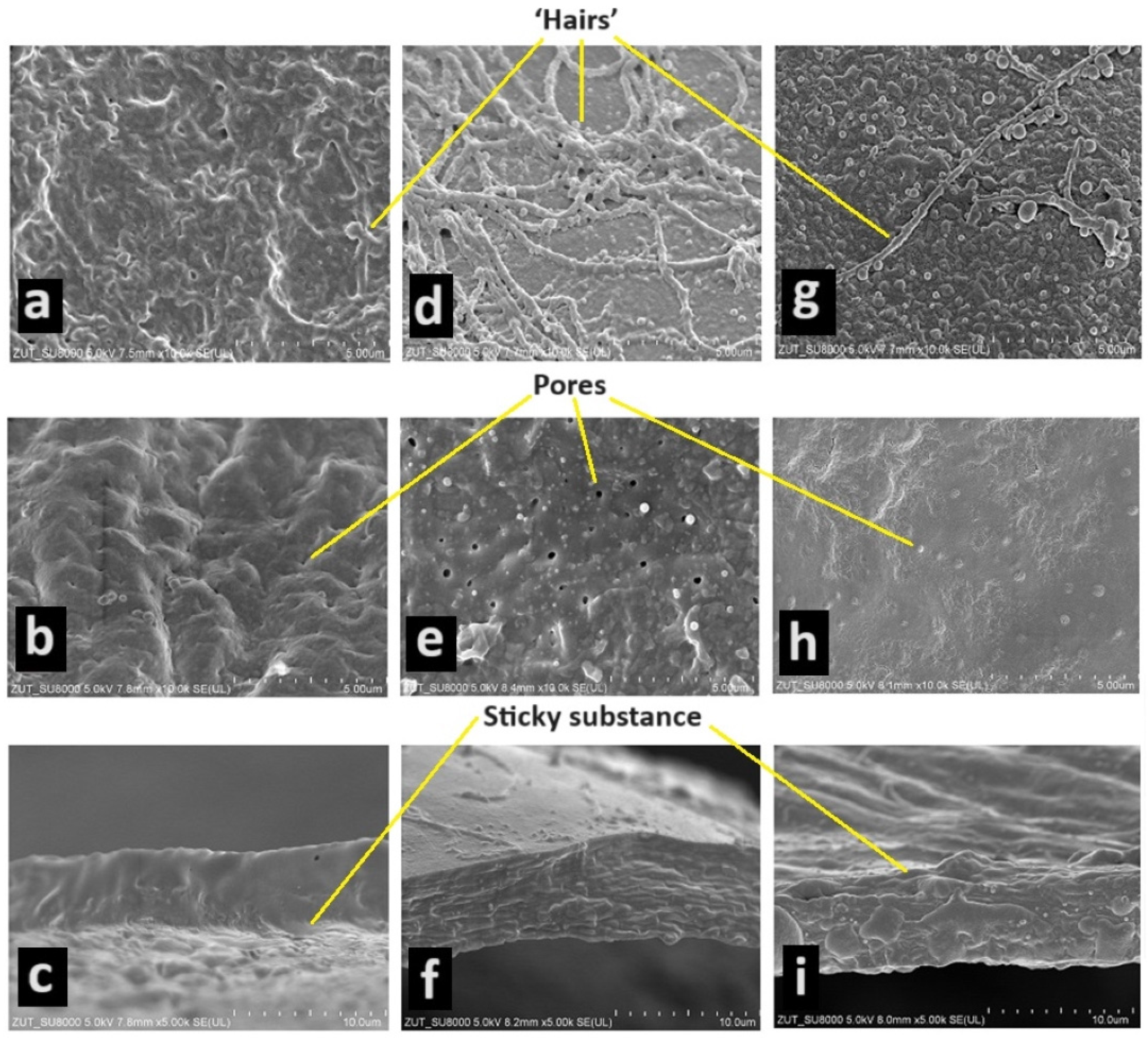
3.1. Analysis of Eggshell Structure
3.2. Observation of Developing Embryos
4. Results
4.1. The Spawning Process
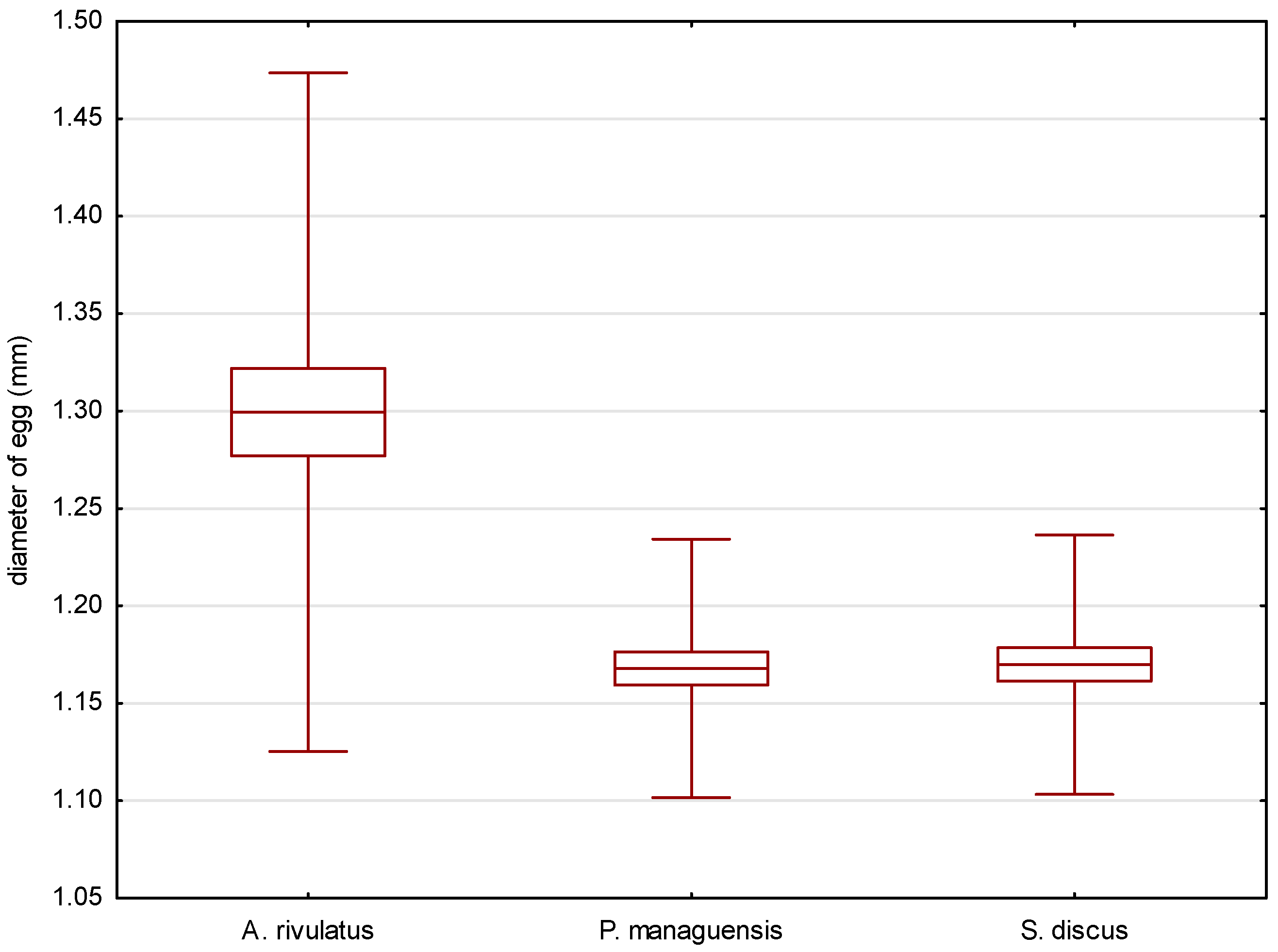
4.2. Characteristics of Eggs
4.3. Embryonic Development
4.4. Cleavage
4.5. Gastrulation
4.6. Organogenesis
4.7. Hatching
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pérez-Vega, M.H.; Uribe-Aranzabal, M.C.; García-Lorenzana, M.; Romero-Ramírez, M.C.; Arredondo-Figueroa, J.L. Description of the ovarian follicle growth of the neotropical cichlids Petenia splendida and Parachromis managuensis (Perciformes: Cichlidae). J. Appl. Ichthyol. 2006, 22, 515–520. [Google Scholar] [CrossRef]
- Angulo, A.; Garita-Alvaro, C.A.; Bussing, W.A.; Lopez, M.I. Annotated checklist of the freshwater fishes of continental and insular Costa Rica: Additions and nomenclatural revisions. Check List 2013, 9, 987–1019. [Google Scholar] [CrossRef]
- Matamoros, W.A.; Schaefer, J.F.; Kreiser, B.R. Annotated checklist of the freshwater fishes of continental and insular Honduras. Zootaxa 2009, 2307, 1–38. [Google Scholar] [CrossRef]
- Nassar, M.A.; Nonell, J.G. Growth of jaguar cichlid (Cichlasoma managuense) juveniles at different oxygen levels. Uniciencia 1992, 9, 3–5. [Google Scholar]
- Bussing, W.A. Peces de las aguas continentales de costa rica. Freshwater fishes of Costa Rica. Rev. Biol. Trop. 1998, 46, 1–468. [Google Scholar]
- Agasen, E.V.; Clemente, J.P., Jr.; Rosana, M.R.; Kawit, N.S. Biological investigation of jaguar guapote Parachromis managuensis (Gunther) in Taal Lake, Philippines. J. Environ. Sci. Manag. 2006, 9, 20–30. [Google Scholar]
- Mendoza-Carranza, M.; Sepúlveda-Lozada, A.; Dias-Ferreira, C.; Geissen, V. Distribution and bioconcentration of heavy metals in a tropical aquatic food web: A case study of a tropical estuarine lagoon in SE Mexico. Environ. Pollut. 2016, 210, 155–165. [Google Scholar] [CrossRef]
- Pease, A.A.; Mendoza-Carranza, M.; Winemiller, K.O. Feeding ecology and ecomorphology of cichlid assemblages in a large Mesoamerican river delta. Environ. Biol. Fishes 2018, 101, 867–879. [Google Scholar] [CrossRef]
- Page, L.M.; Burr, B.M. Peterson Field Guide to Freshwater Fishes of North America, North of Mexico; Houghton Mifflin Company: New York, NY, USA, 2011; pp. 333–335. [Google Scholar]
- Gutierre, S.M.M.; Vitule, J.R.S.; Freire, C.A.; Prodocimo, V. Physiological tools to predict invasiveness and spread via estuarine bridges: Tolerance of Brazilian native and worldwide introduced freshwater fishes to increased salinity. Mar. Freshw. Res. 2014, 65, 425–436. [Google Scholar] [CrossRef]
- Dunseth, D.R.; Bayne, D.R. Recruitment control and production of Tilapia aurea (Steindachner) with the predator, Parachromis managuensis (Günther). Aquaculture 1978, 14, 383–390. [Google Scholar] [CrossRef]
- Conkel, D. Cichlids of North and Central America; T.F.H. Publications Inc.: New York, NY, USA, 1997; p. 65. [Google Scholar] [CrossRef]
- Ortega, H.; Vari, R.P. Annotated Checklist of the Freshwater Fishes of Peru; Smithsonian Institution Press: Washington, DC, USA, 1986; pp. 16–24. ISBN 0-395-91091-9. [Google Scholar]
- Kullander, S.O. Family Cichlidae Check List of the Freshwater Fishes of South and Central America; EDIPUCRS: Porto Alegrel, Brazil, 2003; pp. 605–654. [Google Scholar]
- Musilová, Z.; Říčan, O.; Janko, K.; Novák, J. Molecular phylogeny and biogeography of the Neotropical cichlid fish tribe Cichlasomatini (Teleostei: Cichlidae: Cichlasomatinae). Mol. Phylogenet. Evol. 2008, 46, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Nirchio, M.; Veintimilla, O.C.; Cordero, P.F.Q.; Hernández, J.G.; Oliveira, C. Genotoxic effects of mercury chloride on the Neotropical fish Andinoacara rivulatus (Cichlidae: Cichlasomatini). Rev. Biol. Trop. 2019, 67, 745–754. [Google Scholar] [CrossRef]
- Méndez-Martínez, Y.; Puente-Mendoza, M.M.; Torres-Navarrete, Y.G.; Zamora-Zambrano, R.J.; Botello-León, A.; Ramírez, J.L. Comparative study of serum biochemical and hematological parameters of Andinoacara rivulatus and Ichthyoelephas humeralis in Los Ríos Province, Ecuador. Lat. Am. J. Aquat. Res. 2022, 50, 289–300. [Google Scholar] [CrossRef]
- Franco, M.; Arce, E. Aggressive interactions and consistency of dominance hierarchies of the native and nonnative cichlid fishes of the Balsas basin. Aggress. Behav. 2022, 48, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Bleher, H.; Stölting, K.N.; Salzburger, W.; Meyer, A. Revision of the genus Symphysodon Heckel, 1840 (Teleostei: Perciformes: Cichlidae) based on molecular and morphological characters. Aqua Int. J. Ichthyol. 2007, 12, 133–174. [Google Scholar]
- Crampton, W.G. Ecology and life history of an Amazon floodplain cichlid: The discus fish Symphysodon (Perciformes: Cichlidae). Neotrop. Ichthyol. 2008, 6, 599–612. [Google Scholar] [CrossRef]
- Ready, J.S.; Ferreira, E.J.G.; Kullander, S.O. Discus fishes: Mitochondrial DNA evidence for a phylogeographic barrier in the Amazon genus Symphysodon (Teleostei: Cichlidae). J. Fish Biol. 2006, 69, 200–211. [Google Scholar] [CrossRef]
- Hildemann, W.H. A cichlid fish, Symphysodon discus, with unique nurture habits. Am. Nat. 1959, 93, 27–34. [Google Scholar] [CrossRef]
- Önal, U.; Celik, I.; Cirik, Ş. Histological development of digestive tract in discus, Symphysodon spp. larvae. Aquac. Int. 2009, 18, 589–601. [Google Scholar] [CrossRef]
- Rossoni, F.; Amadio, S.; Ferreira, E.; Zuanon, J. Reproductive and population parameters of discus fish Symphysodon aequifasciatus Pellegrin, 1904 (Perciformes: Cichlidae) from Piagaçu-Purus sustainable development reserve (RDS-PP), lower Purus River, Amazonas, Brazil. Neotrop. Ichthyol. 2010, 8, 379–383. [Google Scholar] [CrossRef]
- Satoh, S.; Tanoue, H.; Mohri, M. Costs and benefits of biparental mucus provisioning in discus fish (Symphysodon aequifasciatus). Ichthyol. Res. 2018, 65, 510–514. [Google Scholar] [CrossRef]
- Moral-Flores, D.; Fernando, L.; López-Segovia, E.; Colis-Torres, A.; Hernández-Arellano, T. Record of three non-native fish species from the Alvarado Lagoon, Veracruz, Mexico. BioInvasions Rec. 2021, 10, 200–209. [Google Scholar] [CrossRef]
- Gonzalez-Martinez, A.; Angón, E.; González, M.A.; Rodríguez Tobar, J.M.; Barba Capote, C.; García Martínez, A.R. Effect of rearing system and sex on the composition and fatty acid profile of Andinoacara rivulatus meat from Ecuador. Rev. Fac. Cienc. Agrar. Univ. Nac. Cuyo 2021, 53, 232–242. [Google Scholar] [CrossRef]
- Mesquita, D.R.; Porto, J.I.R.; Feldberg, E. Chromosomal variability in the wild ornamental species of Symphysodon (Perciformes: Cichlidae) from Amazon. Neotrop. Ichthyol. 2008, 6, 181–190. [Google Scholar] [CrossRef]
- Mattos, D.C.; Screnci-Ribeiro, R.; Cardoso, L.D.; Vidal Junior, M.V. Description of the reproductive behavior of Symphysodon aequifasciatus (Cichlidae) in captivity. Acta Amaz. 2016, 46, 433–438. [Google Scholar] [CrossRef]
- Alvarez-Mieles, G.; Irvine, K.; Griensven, A.V.; Arias-Hidalgo, M.; Torres, A.; Mynett, A.E. Relationships between aquatic biotic communities and water quality in a tropical river–wetland system (Ecuador). Environ. Sci. Policy 2013, 34, 115–127. [Google Scholar] [CrossRef]
- Arias-Hidalgo, M.; Villa-Cox, G.; Griensven, A.V.; Solórzano, G.; Villa-Cox, R.; Mynett, A.E.; Debels, P. A decision framework for wetland management in a river basin context: The “Abras de Mantequilla” case study in the Guayas River Basin, Ecuador. Environ. Sci. Policy 2013, 34, 103–114. [Google Scholar] [CrossRef]
- Resende, A.G.A.; França, E.J.D.; Oliveira, C.D.L.D.; Santana, F.M. Maturity, growth and natural mortality rate of the introduced fish Parachromis managuensis (Perciformes: Cichlidae) in the semiarid region of Brazil. Acta Limnol. Bras. 2020, 32, 2–7. [Google Scholar] [CrossRef]
- Prazdnikov, D.V. Influence of Triiodothyronine (T3) on the Reproduction and Development of the Green Terror Andinoacara rivulatus (Cichlidae). J. Ichthyol. 2018, 58, 953–958. [Google Scholar] [CrossRef]
- Tirado, J.O.; Bangeppagari, M.; Benavides, A.; Muñoz, D.; Romero, P.; Lee, S.J. Steroid profiles of wild terror green (Andinoacara rivulatus) associated with gonadal histology in the Baba river, Ecuador. Int. J. Adv. Sci. 2021, 11, 1612. [Google Scholar] [CrossRef]
- Townshend, T.J.; Wootton, J. Effects of food supply on the reproduction of the convict cichlid, Cichlasoma nigrofasciatum. J. Fish Biol. 1984, 24, 91–104. [Google Scholar] [CrossRef]
- Coward, K.; Bromage, N.R. Spawning periodicity, fecundity and egg size in laboratory-held stocks of a substrate-spawning tilapiine, Tilapia zillii (Gervais). Aquaculture 1999, 171, 251–267. [Google Scholar] [CrossRef]
- Teresa, F.B.; Gonçalves-de-Freitas, E. Reproductive behavior and parental roles of the cichlid fish Laetacara araguaiae. Neotrop. Ichthyol. 2011, 9, 355–362. [Google Scholar] [CrossRef]
- Kratochwil, C.F.; Sefton, M.M.; Meyer, A. Embryonic and larval development in the Midas cichlid fish species flock (Amphilophus spp.): A new evo-devo model for the investigation of adaptive novelties and species differences. BMC Dev. Biol. 2015, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Salis, P.; Lee, S.H.; Roux, N.; Lecchini, D.; Laudet, V. The real Nemo movie: Description of embryonic development in Amphiprion ocellaris from first division to hatching. Dev. Dyn. 2021, 250, 1651–1667. [Google Scholar] [CrossRef] [PubMed]
- Brylińska, M. Ryby słodkowodne Polski; Wyd. Nauk. PWN: Warsaw, Poland, 1986; pp. 116–124. ISBN 83-01131-00-4. [Google Scholar]
- Reid, J.E.; Chaput, G. Spawning history influence on fecundity, egg size, and egg survival of Atlantic salmon (Salmo salar) from the Miramichi River, New Brunswick, Canada. J. Mar. Sci. 2012, 69, 1678–1685. [Google Scholar] [CrossRef]
- Spíndola Linhares, J.C.; Resende Manna, L.; Mazzoni, R.; Ferreira Rezende, C.; Feitosa Silva, J.R. Reproductive tactics optimizing the survival of the offspring of Cichlasoma orientale (Perciformes: Cichlidae). Rev. Biol. Trop. 2014, 62, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Rollinson, N.; Rowe, L. The positive correlation between maternal size and offspring size: Fitting pieces of a life-history puzzle. Biol. Rev. 2016, 91, 1134–1148. [Google Scholar] [CrossRef] [PubMed]
- Chávez-López, R.; Peterson, M.S.; Brown-Peterson, N.J.; Morales-Gómez, A.A.; Franco-López, J. Ecology of the Mayan Cichlid, Cichlasoma urophthalmus Günther, in the Alvarado Lagoonal System, Veracruz, Mexico. Gulf Caribb. Res. 2005, 16, 123–131. [Google Scholar] [CrossRef]
- Kupren, K.; Żarski, D.; Targońska, K.; Rzecznego, K.R.J. Rozród Karpiowatych ryb Reofilnych w Warunkach Kontrolowanych. Wybrane Aspekty Rozrodu Karpiowatych ryb Reofilnych w Warunkach Kontrolowanych; Mercurius: Olsztyn, Poland, 2008; pp. 54–55. ISBN 978-83-923-855-4-7. [Google Scholar]
- Paes, M.D.C.F.; Makino, L.C.; Vasquez, L.A.; Fernandes, J.B.K.; Nakaghi, L.S.O. Early development of Astronotus ocellatus under stereomicroscopy and scanning electron microscopy. Zygote 2012, 20, 269–276. [Google Scholar] [CrossRef]
- Coward, K.; Bromage, N.R. Histological classification of oocyte growth and the dynamics of ovarian recrudescence in Tilapia zillii. J. Fish Biol. 1998, 53, 285–302. [Google Scholar] [CrossRef]
- Fleig, R. Embryogenesis in mouth-breeding cichlids (Osteichthyes, Teleostei) structure and fate of the enveloping layer. Roux’s Arch. Dev. Biol. 1993, 203, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Korzelecka-Orkisz, A.; Szalast, Z.; Pawlos, D.; Smaruj, I.; Tañski, A.; Szulc, J.; Formicki, K. Early ontogenesis of the angelfish, Pterophyllum scalare Schultze, 1823 (Cichlidae). Neotrop. Ichthyol. 2012, 10, 567–576. [Google Scholar] [CrossRef]
- Tański, A.; Korzelecka, A.; Bonisławska, M.; Winnicki, A.; Formicki, K. New data on morphomechanical changes during embryogenesis of pike (Esox lucius L.). Piscaria 2000, 27, 207–214. [Google Scholar]
- Horbowa, K.; Fey, D.P. Atlas Wczesnych Stadiów Rozwojowych ryb. 34 Gatunki ryb Bałtyku Południowego i Jego Zalewów; Wydawnictwo Morski Instytut Rybacki—Państwowy Instytut Badawczy: Gdynia, Poland, 2013; ISBN 978-83-61650-10-2. [Google Scholar]
- Krzystolik, J.; Tański, A.; Korzelecka-Orkisz, A.; Formicki, K.; Piesiewicz, R.; Pender, R.; Potkański, Ł. Survival of sea trout (Salmo trutta m.trutta) eggs in selected rivers of Western Pomerania. In Activities of Fishing and Angling Entities in 2020; Kowalska, A., Wołos, A., Eds.; IRS Publishing House: Olsztyn, Poland, 2021; pp. 139–147. (In Polish) [Google Scholar]
- Lugowska, K.; Witeska, M. The effect of temperature on early development of barbel Barbus barbus (L.). Aquac. Res. 2018, 49, 2495–2502. [Google Scholar] [CrossRef]
- Dubey, M.K.; Kamalam, B.S.; Rajesh, M.; Sarma, D.; Pandey, A.; Baral, P.; Sharma, P. Exposure to different temperature regimes at early life stages affects hatching, developmental morphology, larval growth, and muscle cellularity in rainbow trout, Oncorhynchus mykiss. Fish Physiol. Biochem. 2023, 49, 219–238. [Google Scholar] [CrossRef]
- Syafariyah, N.K.; Sulmartiwi, L.; Budi, D.S. Incubation temperature effects on some hatching parameters of silver rasbora (Rasbora argyrotaenia) egg. J. Appl. Aquac. 2023, 35, 16–26. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Jaguar Cichlid | Green Terror | Red Discus | |||||
|---|---|---|---|---|---|---|---|
| Degree Hour (°H) | Hour (Hpf) | Degree Hour (°H) | Hour (Hpf) | Degree Hour (°H) | Hour (Hpf) | ||
| Fertilization | 0 | 0 | 0 | 0 | 0 | 0 | |
| Cleavage | 2 blastomeres | 43 | 1.66 | 39 | 1.5 | 26 | 1 |
| 4 blastomeres | 56 | 2.16 | 52 | 2 | 39 | 1.5 | |
| 8 blastomeres | 69 | 2.66 | 61 | 2.35 | 52 | 2 | |
| 16 blastomeres | 82 | 3.16 | 78 | 3 | 65 | 2.5 | |
| Gastrulation | beginning | 520 | 20 | 468 | 18 | 455 | 17.5 |
| epiboly 1/3 | 598 | 23 | 572 | 22 | 546 | 21 | |
| epiboly 2/3 | 728 | 28 | 650 | 25 | 637 | 24.5 | |
| 3/4 epiboly | 910 | 35 | 741 | 28.5 | 728 | 28 | |
| Closure of the blastopore | 1066 | 41 | 858 | 33 | 845 | 32.5 | |
| Organogenesis | Making the head part visible | 1118 | 43 | 1040 | 40 | 1014 | 39 |
| Eye primordia | 1248 | 48 | 1118 | 43 | 1092 | 42 | |
| First heart contractions | 1443 | 55.5 | 1170 | 45 | 1105 | 42.5 | |
| First movements of the embryo | 1651 | 63.5 | 1404 | 54 | 1339 | 51.5 | |
| Making the brain visible | 1677 | 64.5 | 1456 | 56 | 1443 | 55.5 | |
| Pigment cells become visible in the lens | 1716 | 66 | 1508 | 58 | 1495 | 57.5 | |
| The appearance of pigment cells on the body | 1794 | 69 | 1924 | 74 | 1872 | 72 | |
| Red Discus | Jaguar Cichlid | Green Terror | |||||
|---|---|---|---|---|---|---|---|
| Degree Hour (°H) | Hour (Hpf) | Degree Hour (°H) | Hour (Hpf) | Degree Hour(°H) | Hour (Hpf) | ||
| Hatching | First hatching | 2132 | 82 | 2262 | 87 | 2158 | 83 |
| Hatching 50% | 2210 | 85 | 2366 | 91 | 2236 | 86 | |
| Hatching 100% | 2314 | 89 | 2470 | 95 | 2340 | 90 | |
| Larvae after hatching | |||
|---|---|---|---|
| Body length | Length of yolk sac | Width of the yolk sac | Volume of the yolk sac |
| 5.301 mm | 1.988 mm | 1.345 mm | 1.885 mm3 |
| Larvae after starting to swim | |||
| Body length | Length of yolk sac | Width of the yolk sac | Volume of the yolk sac |
| 6.545 mm | 1.518 mm | 1.123 mm | 1.024 mm3 |
| Larvae immediately after hatching | |||
|---|---|---|---|
| Body length | Length of yolk sac | Width of the yolk sac | Volume of the yolk sac |
| 5.203 mm | 1.554 mm | 1.182 mm | 1.137 mm3 |
| Larvae immediately after starting to swim | |||
| Body length | Length of yolk sac | Width of the yolk sac | Volume of the yolk sac |
| 6.136 mm | 1.106 mm | 0.754 mm | 0.329 mm3 |
| Larvae immediately after hatching | |||
|---|---|---|---|
| Body length | Length of yolk sac | Width of the yolk sac | Volume of the yolk sac |
| 4.346 mm | 1.403 mm | 1.112 mm | 0.909 mm3 |
| Larvae immediately after starting to swim | |||
| Body length | Length of yolk sac | Width of the yolk sac | Volume of the yolk sac |
| 5.447 mm | 0.987 mm | 0.882 mm | 0.402 mm3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piesiewicz, R.; Krzystolik, J.; Korzelecka-Orkisz, A.; Tański, A.; Formicki, K. Early Ontogeny of Cichlids Using Selected Species as Examples. Animals 2024, 14, 1238. https://doi.org/10.3390/ani14081238
Piesiewicz R, Krzystolik J, Korzelecka-Orkisz A, Tański A, Formicki K. Early Ontogeny of Cichlids Using Selected Species as Examples. Animals. 2024; 14(8):1238. https://doi.org/10.3390/ani14081238
Chicago/Turabian StylePiesiewicz, Radosław, Jan Krzystolik, Agata Korzelecka-Orkisz, Adam Tański, and Krzysztof Formicki. 2024. "Early Ontogeny of Cichlids Using Selected Species as Examples" Animals 14, no. 8: 1238. https://doi.org/10.3390/ani14081238
APA StylePiesiewicz, R., Krzystolik, J., Korzelecka-Orkisz, A., Tański, A., & Formicki, K. (2024). Early Ontogeny of Cichlids Using Selected Species as Examples. Animals, 14(8), 1238. https://doi.org/10.3390/ani14081238