
Role of Maternal Antibodies in the Protection of Broiler Chicks against Campylobacter Colonization in the First Weeks of Life

 , , , ,
, , , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
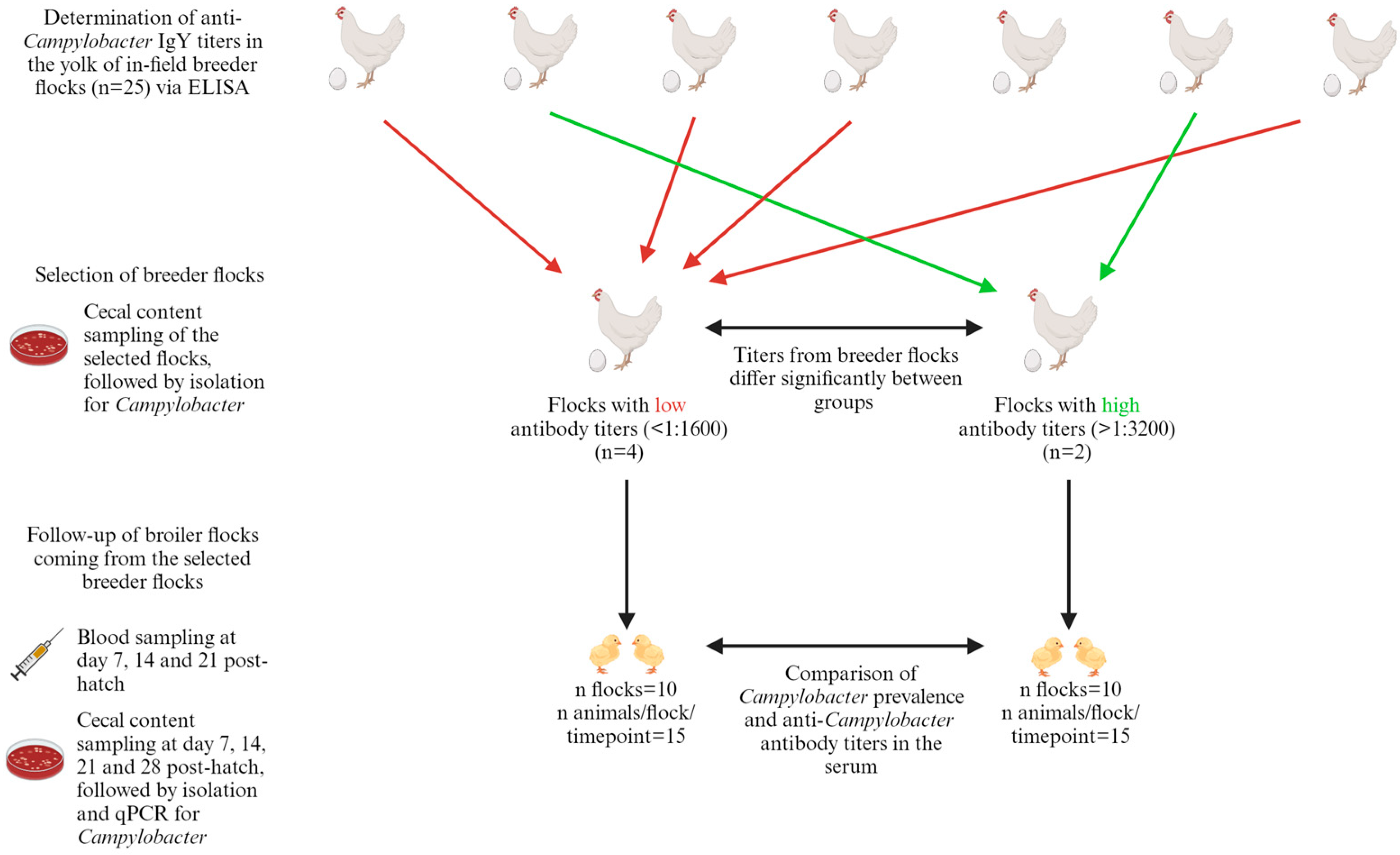
2.1. Field Study: Anti-Campylobacter IgY in Broiler Breeders’ Egg Yolks and Resulting IgY Titers and Probability of Colonization in Their Offspring
2.2. Vaccination Study: Selection and Immunization of in Field Seropositive Broiler Breeders and Determination of Maternal Antibodies after Vaccination
2.3. Vaccination Study: Protective Efficacy of Bacterin and Subunit Derived Maternal Antibodies on Cecal Colonization in Broilers Using a Threshold Model
2.4. Bacterial Strains, Culture Conditions and Isolation
2.5. Characterization of Campylobacter Isolates
2.6. Bacterin and Subunit Vaccine Preparation
2.7. Determination of IgY Titers in Egg Yolk and Serum
2.8. Cecal Campylobacter Enumeration by qPCR
2.9. Statistical Analysis
3. Results
3.1. Field Study: Trend of Decreased Susceptibility to Campylobacter Colonization in Broilers Originating from Breeders with High Antibody Levels
3.2. Field Study: C. coli Dominant in Breeders, an Equal Prevalence of C. jejuni and C. coli in Broilers
3.3. Field Study: Low Maternal Antibody Titers in Broilers from Breeders with Varying Antibody Levels
3.4. Vaccination Study: Immunization of Seropositive Broiler Breeders Results in Temporarily Higher Anti-Campylobacter Titers in Broiler Breeders
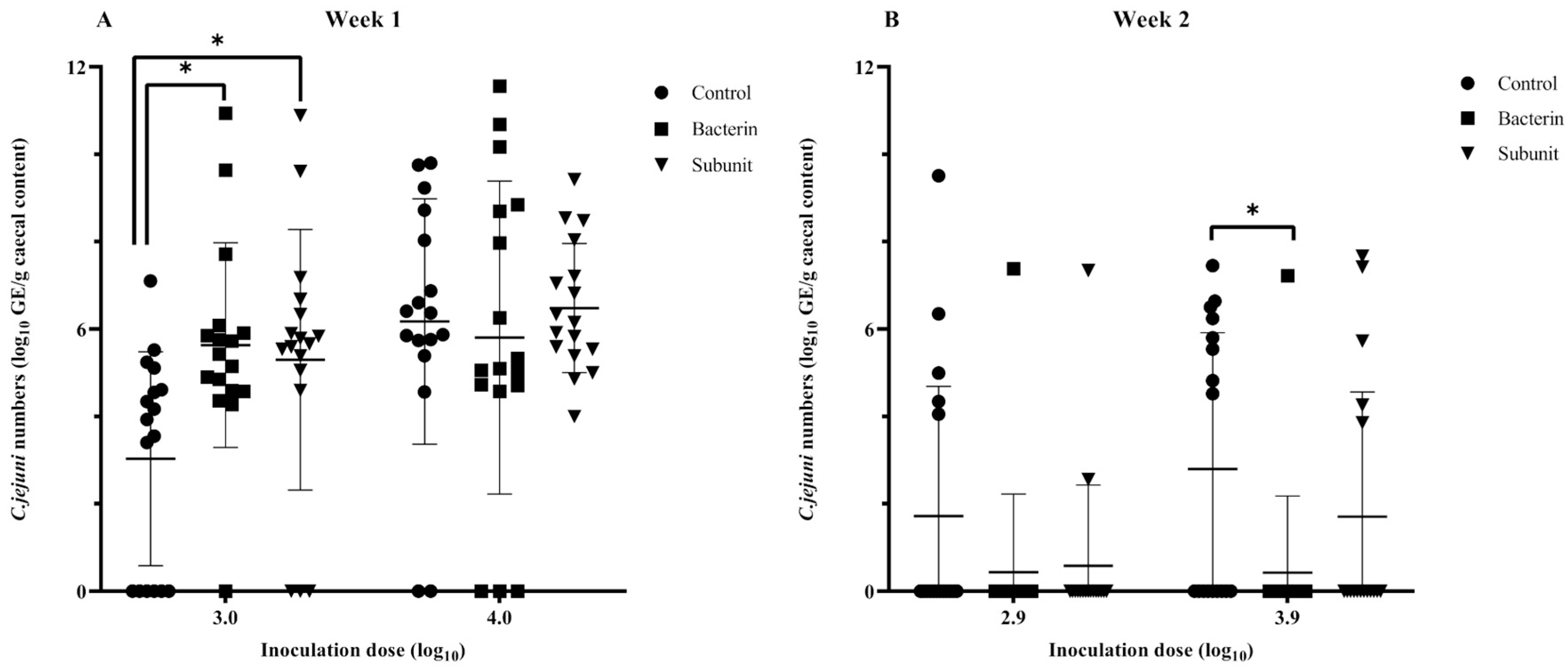
3.5. Vaccination Study: Protective Efficacy of Bacterin and Subunit Derived Maternal Antibodies on Cecal Colonization in Broilers Using a Threshold Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nachamkin, I.; Allos, B.M.; Ho, T. Campylobacter Species and Guillain-Barré Syndrome. Clin. Microbiol. Rev. 1998, 11, 555–567. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union One Health 2022 Zoonoses Report. EFSA J. 2023, 21, e8442. [Google Scholar] [CrossRef] [PubMed]
- Herman, L.; Heyndrickx, M.; Grijspeerdt, K.; Vandekerckhove, D.; Rollier, I.; De ZUTTER, L. Routes for Campylobacter Contamination of Poultry Meat: Epidemiological Study from Hatchery to Slaughterhouse. Epidemiol. Infect. 2003, 131, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Rasschaert, G.; Houf, K.; Van Hende, J.; De Zutter, L. Campylobacter Contamination during Poultry Slaughter in Belgium. J. Food Prot. 2006, 69, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Seliwiorstow, T.; Duarte, A.; Baré, J.; Botteldoorn, N.; Dierick, K.; Uyttendaele, M.; De Zutter, L. Comparison of Sample Types and Analytical Methods for the Detection of Highly Campylobacter-Colonized Broiler Flocks at Different Stages in the Poultry Meat Production Chain. Foodborne Pathog. Dis. 2015, 12, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Hertogs, K.; Haegeman, A.; Schaumont, D.; Gelaude, P.; De Zutter, L.; Dewulf, J.; Heyndrickx, M.; Rasschaert, G. Contamination Sources and Transmission Routes for Campylobacter on (Mixed) Broiler Farms in Belgium, and Comparison of the Gut Microbiota of Flocks Colonized and Uncolonized with Campylobacter. Pathogens 2021, 10, 66. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, S.; Chaloner, G.; Kemmett, K.; Davidson, N.; Williams, N.; Kipar, A.; Humphrey, T.; Wigley, P. Campylobacter Jejuni Is Not Merely a Commensal in Commercial Broiler Chickens and Affects Bird Welfare. mBio 2014, 5, e01364-14. [Google Scholar] [CrossRef]
- Pédron, T.; Sansonetti, P. Commensals, Bacterial Pathogens and Intestinal Inflammation: An Intriguing Ménage à Trois. Cell Host Microbe 2008, 3, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Hermans, D.; Van Deun, K.; Messens, W.; Martel, A.; Van Immerseel, F.; Haesebrouck, F.; Rasschaert, G.; Heyndrickx, M.; Pasmans, F. Campylobacter Control in Poultry by Current Intervention Measures Ineffective: Urgent Need for Intensified Fundamental Research. Vet. Microbiol. 2011, 152, 219–228. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Shehata, A.M.; Arif, M.; Paswan, V.K.; Batiha, G.E.-S.; Khafaga, A.F.; Elbestawy, A.R. Approaches to Prevent and Control Campylobacter spp. Colonization in Broiler Chickens: A Review. Environ. Sci. Pollut. Res. 2021, 28, 4989–5004. [Google Scholar] [CrossRef]
- Sahin, O.; Morishita, T.Y.; Zhang, Q. Campylobacter Colonization in Poultry: Sources of Infection and Modes of Transmission. Anim. Health Res. Rev. 2002, 3, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Newell, D.G.; Elvers, K.T.; Dopfer, D.; Hansson, I.; Jones, P.; James, S.; Gittins, J.; Stern, N.J.; Davies, R.; Connerton, I.; et al. Biosecurity-Based Interventions and Strategies to Reduce Campylobacter spp. on Poultry Farms. Appl. Environ. Microbiol. 2011, 77, 8605–8614. [Google Scholar] [CrossRef] [PubMed]
- Cox, N.A.; Richardson, L.J.; Maurer, J.J.; Berrang, M.E.; Fedorka-Cray, P.J.; Buhr, R.J.; Byrd, J.A.; Lee, M.D.; Hofacre, C.L.; O’Kane, P.M.; et al. Evidence for Horizontal and Vertical Transmission in Campylobacter Passage from Hen to Her Progeny. J. Food Prot. 2012, 75, 1896–1902. [Google Scholar] [CrossRef] [PubMed]
- Callicott, K.A.; Friðriksdóttir, V.; Reiersen, J.; Lowman, R.; Bisaillon, J.-R.; Gunnarsson, E.; Berndtson, E.; Hiett, K.L.; Needleman, D.S.; Stern, N.J. Lack of Evidence for Vertical Transmission of Campylobacter spp. in Chickens. Appl. Environ. Microbiol. 2006, 72, 5794–5798. [Google Scholar] [CrossRef] [PubMed]
- Shanker, S.; Lee, A.; Sorrell, T.C. Campylobacter jejuni in Broilers: The Role of Vertical Transmission. J. Hyg. 1986, 96, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Evans, S.J.; Sayers, A.R. A Longitudinal Study of Campylobacter Infection of Broiler Flocks in Great Britain. Prev. Vet. Med. 2000, 46, 209–223. [Google Scholar] [CrossRef]
- Jacobs-Reitsma, W.F.; Van de Giessen, A.W.; Bolder, N.M.; Mulder, R.W.A.W. Epidemiology of Campylobacter spp. at Two Dutch Broiler Farms. Epidemiol. Infect. 1995, 114, 413–421. [Google Scholar] [CrossRef]
- Schoeni, J.L.; Doyle, M.P. Reduction of Campylobacter Jejuni Colonization of Chicks by Cecum-Colonizing Bacteria Producing Anti-C. Jejuni Metabolites. Appl. Environ. Microbiol. 1992, 58, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Sahin, O.; Luo, N.; Huang, S.; Zhang, Q. Effect of Campylobacter-Specific Maternal Antibodies on Campylobacter jejuni Colonization in Young Chickens. Appl. Environ. Microbiol. 2003, 69, 5372–5379. [Google Scholar] [CrossRef]
- Cawthraw, S.A.; Newell, D.G. Investigation of the Presence and Protective Effects of Maternal Antibodies Against Campylobacter jejuni in Chickens. Avian Dis. 2010, 54, 86–93. [Google Scholar] [CrossRef]
- Sahin, O.; Zhang, Q.; Meitzler, J.C.; Harr, B.S.; Morishita, T.Y.; Mohan, R. Prevalence, Antigenic Specificity, and Bactericidal Activity of Poultry Anti- Campylobacter Maternal Antibodies. Appl. Environ. Microbiol. 2001, 67, 3951–3957. [Google Scholar] [CrossRef]
- Haems, K.; Van Rysselberghe, N.; Goossens, E.; Strubbe, D.; Rasschaert, G.; Martel, A.; Pasmans, F.; Garmyn, A. Reducing Campylobacter Colonization in Broilers by Active Immunization of Naive Broiler Breeders Using a Bacterin and Subunit Vaccine. Poult. Sci. 2023, 102, 103075. [Google Scholar] [CrossRef]
- Hermans, D.; Martel, A.; Garmyn, A.; Verlinden, M.; Heyndrickx, M.; Gantois, I.; Haesebrouck, F.; Pasmans, F. Application of Medium-Chain Fatty Acids in Drinking Water Increases Campylobacter jejuni Colonization Threshold in Broiler Chicks. Poult. Sci. 2012, 91, 1733–1738. [Google Scholar] [CrossRef] [PubMed]
- Hermans, D.; Van Steendam, K.; Verbrugghe, E.; Verlinden, M.; Martel, A.; Seliwiorstow, T.; Heyndrickx, M.; Haesebrouck, F.; De Zutter, L.; Deforce, D.; et al. Passive Immunization to Reduce Campylobacter jejuni Colonization and Transmission in Broiler Chickens. Vet. Res. 2014, 45, 27. [Google Scholar] [CrossRef]
- Vandeputte, J.; Martel, A.; Canessa, S.; Van Rysselberghe, N.; De Zutter, L.; Heyndrickx, M.; Haesebrouck, F.; Pasmans, F.; Garmyn, A. Reducing Campylobacter jejuni Colonization in Broiler Chickens by in-Feed Supplementation with Hyperimmune Egg Yolk Antibodies. Sci. Rep. 2019, 9, 8931. [Google Scholar] [CrossRef]
- Van Deun, K.; Pasmans, F.; Ducatelle, R.; Flahou, B.; Vissenberg, K.; Martel, A.; Van den Broeck, W.; Van Immerseel, F.; Haesebrouck, F. Colonization Strategy of Campylobacter jejuni Results in Persistent Infection of the Chicken Gut. Vet. Microbiol. 2008, 130, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Phelan, V.V.; Simkovsky, R.; Watrous, J.D.; Trial, R.M.; Fleming, T.C.; Wenter, R.; Moore, B.S.; Golden, S.S.; Pogliano, K.; et al. Primer on Agar-Based Microbial Imaging Mass Spectrometry. J. Bacteriol. 2012, 194, 6023–6028. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-Access Bacterial Population Genomics: BIGSdb Software, the PubMLST.Org Website and Their Applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Nieves, W.; Heang, J.; Asakrah, S.; Höner zu Bentrup, K.; Roy, C.J.; Morici, L.A. Immunospecific Responses to Bacterial Elongation Factor Tu during Burkholderia Infection and Immunization. PLoS ONE 2010, 5, e14361. [Google Scholar] [CrossRef]
- Tsugawa, H.; Ito, H.; Ohshima, M.; Okawa, Y. Cell Adherence-Promoted Activity of Plesiomonas shigelloides GroEL. J. Med. Microbiol. 2007, 56, 23–29. [Google Scholar] [CrossRef]
- Zhang, M.; Meng, F.; Cao, F.; Qiao, B.; Liu, G.; Liu, H.; Zhou, Y.; Dong, H.; Gu, Y.; Xiao, D.; et al. Cloning, Expression, and Antigenicity of 14 Proteins from Campylobacter jejuni. Foodborne Pathog. Dis. 2012, 9, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Lardeux, F.; Torrico, G.; Aliaga, C. Calculation of the ELISA’s Cut-off Based on the Change-Point Analysis Method for Detection of Trypanosoma cruzi Infection in Bolivian Dogs in the Absence of Controls. Memórias Inst. Oswaldo Cruz 2016, 111, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Killick, R.; Eckley, I.A. Changepoint: An R Package for Changepoint Analysis. J. Stat. Softw. 2014, 58, 1–19. [Google Scholar] [CrossRef]
- Lund, M.; Nordentoft, S.; Pedersen, K.; Madsen, M. Detection of Campylobacter spp. in ChickenFecal Samples by Real-TimePCR. J. Clin. Microbiol. 2004, 42, 5125–5132. [Google Scholar] [CrossRef] [PubMed]
- Botteldoorn, N.; Van Coillie, E.; Piessens, V.; Rasschaert, G.; Debruyne, L.; Heyndrickx, M.; Herman, L.; Messens, W. Quantification of Campylobacter spp. in Chicken Carcass Rinse by Real-Time PCR. J. Appl. Microbiol. 2008, 105, 1909–1918. [Google Scholar] [CrossRef]
- Rasschaert, G.; Houf, K.; Van Hende, J.; De Zutter, L. Investigation of the Concurrent Colonization with Campylobacter and Salmonella in Poultry Flocks and Assessment of the Sampling Site for Status Determination at Slaughter. Vet. Microbiol. 2007, 123, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Meunier, M.; Guyard-Nicodème, M.; Vigouroux, E.; Poezevara, T.; Béven, V.; Quesne, S.; Amelot, M.; Parra, A.; Chemaly, M.; Dory, D. A DNA Prime/Protein Boost Vaccine Protocol Developed against Campylobacter jejuni for Poultry. Vaccine 2018, 36, 2119–2125. [Google Scholar] [CrossRef] [PubMed]
- Mortada, M.; Cosby, D.E.; Akerele, G.; Ramadan, N.; Oxford, J.; Shanmugasundaram, R.; Ng, T.T.; Selvaraj, R.K. Characterizing the Immune Response of Chickens to Campylobacter jejuni (Strain A74C). PLoS ONE 2021, 16, e0247080. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Zhou, Q.; Tang, M.-J.; Pu, J.-H.; Fan, Y.-F.; Lu, J.-X.; Huang, J.-L.; Gao, Y.-S. Expression of the Campylobacter jejuni FliD Protein and Its Reaction to Chicken Sera. FEMS Microbiol. Lett. 2020, 367, fnaa115. [Google Scholar] [CrossRef]
- Vandeplas, S.; Dubois-Dauphin, R.; Palm, R.; Beckers, Y.; Thonart, P.; Théwis, A. Prevalence and Sources of Campylobacter spp. Contamination in Free-Range Broiler Production in the Southern Part of Belgium. Biotechnol. Agron. Soc. Environ. 2010, 14, 279–288. [Google Scholar]
- Schets, F.M.; Jacobs-Reitsma, W.F.; van der Plaats, R.Q.J.; Heer, L.K.-D.; van Hoek, A.H.A.M.; Hamidjaja, R.A.; de Roda Husman, A.M.; Blaak, H. Prevalence and Types of Campylobacter on Poultry Farms and in Their Direct Environment. J. Water Health 2017, 15, 849–862. [Google Scholar] [CrossRef]
- Iannetti, S.; Calistri, P.; Di Serafino, G.; Marotta, F.; Alessiani, A.; Antoci, S.; Neri, D.; Perilli, M.; Iannitto, G.; Iannetti, L.; et al. Campylobacter jejuni and Campylobacter coli: Prevalence, Contamination Levels, Genetic Diversity and Antibiotic Resistance in Italy. Vet. Ital. 2020, 56, 23–34. [Google Scholar] [CrossRef]
- García-Sánchez, L.; Melero, B.; Diez, A.M.; Jaime, I.; Canepa, A.; Rovira, J. Genotyping, Virulence Genes and Antimicrobial Resistance of Campylobacter spp. Isolated during Two Seasonal Periods in Spanish Poultry Farms. Prev. Vet. Med. 2020, 176, 104935. [Google Scholar] [CrossRef]
- Marinou, I.; Bersimis, S.; Ioannidis, A.; Nicolaou, C.; Mitroussia-Ziouva, A.; Legakis, N.J.; Chatzipanagiotou, S. Identification and Antimicrobial Resistance of Campylobacter Species Isolated from Animal Sources. Front. Microbiol. 2012, 3, 58. [Google Scholar] [CrossRef]
- Repérant, E.; Laisney, M.J.; Nagard, B.; Quesne, S.; Rouxel, S.; Le Gall, F.; Chemaly, M.; Denis, M. Influence of Enrichment and Isolation Media on the Detection of Campylobacter spp. in Naturally Contaminated Chicken Samples. J. Microbiol. Methods 2016, 128, 42–47. [Google Scholar] [CrossRef]
- Wieczorek, K.; Kania, I.; Osek, J. Prevalence and Antimicrobial Resistance of Campylobacter spp. Isolated from Poultry Carcasses in Poland. J. Food Prot. 2013, 76, 1451–1455. [Google Scholar] [CrossRef]
- Andritsos, N.D.; Tzimotoudis, N.; Mataragas, M. Prevalence and Distribution of Thermotolerant Campylobacter Species in Poultry: A Comprehensive Review with a Focus on the Factors Affecting the Detection and Enumeration of Campylobacter jejuni and Campylobacter coli in Chicken Meat. Appl. Sci. 2023, 13, 8079. [Google Scholar] [CrossRef]
- Babacan, O.; Harris, S.A.; Pinho, R.M.; Hedges, A.; Jørgensen, F.; Corry, J.E.L. Factors Affecting the Species of Campylobacter Colonizing Chickens Reared for Meat. J. Appl. Microbiol. 2020, 129, 1071–1078. [Google Scholar] [CrossRef]
- El-Shibiny, A.; Connerton, P.L.; Connerton, I.F. Campylobacter Succession in Broiler Chickens. Vet. Microbiol. 2007, 125, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Lapierre, L.; Gatica, M.A.; Riquelme, V.; Vergara, C.; Yañez, J.M.; San Martín, B.; Sáenz, L.; Vidal, M.; Martínez, M.C.; Araya, P.; et al. Characterization of Antimicrobial Susceptibility and Its Association with Virulence Genes Related to Adherence, Invasion, and Cytotoxicity in Campylobacter jejuni and Campylobacter coli Isolates from Animals, Meat, and Humans. Microb. Drug Resist. 2016, 22, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, A.; Johansson, C.; Skarp, A.; Kaden, R.; Bertilsson, S.; Rautelin, H. Survival of Campylobacter jejuni and Campylobacter coli Water Isolates in Lake and Well Water. APMIS 2018, 126, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, K.; Bocian, Ł.; Osek, J. Prevalence and Antimicrobial Resistance of Campylobacter Isolated from Carcasses of Chickens Slaughtered in Poland—A Retrospective Study. Food Control 2020, 112, 107159. [Google Scholar] [CrossRef]
- Dingle, K.E.; Colles, F.M.; Falush, D.; Maiden, M.C.J. Sequence Typing and Comparison of Population Biology of Campylobacter coli and Campylobacter jejuni. J. Clin. Microbiol. 2005, 43, 340–347. [Google Scholar] [CrossRef]
- Kittl, S.; Heckel, G.; Korczak, B.M.; Kuhnert, P. Source Attribution of Human Campylobacter Isolates by MLST and Fla-Typing and Association of Genotypes with Quinolone Resistance. PLoS ONE 2013, 8, e81796. [Google Scholar] [CrossRef]
- Miller, W.G.; Englen, M.D.; Kathariou, S.; Wesley, I.V.; Wang, G.; Pittenger-Alley, L.; Siletz, R.M.; Muraoka, W.; Fedorka-Cray, P.J.; Mandrell, R.E. Identification of Host-Associated Alleles by Multilocus Sequence Typing of Campylobacter coli Strains from Food Animals. Microbiology 2006, 152, 245–255. [Google Scholar] [CrossRef]
- Nohra, A.; Grinberg, A.; Midwinter, A.C.; Marshall, J.C.; Collins-Emerson, J.M.; French, N.P. Molecular Epidemiology of Campylobacter coli Strains Isolated from Different Sources in New Zealand between 2005 and 2014. Appl. Environ. Microbiol. 2016, 82, 4363–4370. [Google Scholar] [CrossRef]
- Sheppard, S.K.; Dallas, J.F.; Strachan, N.J.C.; MacRae, M.; McCarthy, N.D.; Wilson, D.J.; Gormley, F.J.; Falush, D.; Ogden, I.D.; Maiden, M.C.J.; et al. Campylobacter Genotyping to Determine the Source of Human Infection. Clin. Infect. Dis. 2009, 48, 1072–1078. [Google Scholar] [CrossRef]
- Thakur, S.; Morrow, W.E.M.; Funk, J.A.; Bahnson, P.B.; Gebreyes, W.A. Molecular Epidemiologic Investigation of Campylobacter coli in Swine Production Systems, Using Multilocus Sequence Typing. Appl. Environ. Microbiol. 2006, 72, 5666–5669. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.; Seliwiorstow, T.; Miller, W.G.; De Zutter, L.; Uyttendaele, M.; Dierick, K.; Botteldoorn, N. Discriminative Power of Campylobacter Phenotypic and Genotypic Typing Methods. J. Microbiol. Methods 2016, 125, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Elhadidy, M.; Miller, W.G.; Arguello, H.; Álvarez-Ordóñez, A.; Dierick, K.; Botteldoorn, N. Molecular Epidemiology and Antimicrobial Resistance Mechanisms of Campylobacter coli from Diarrhoeal Patients and Broiler Carcasses in Belgium. Transbound. Emerg. Dis. 2019, 66, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Habib, I.; Uyttendaele, M.; De Zutter, L. Survival of Poultry-Derived Campylobacter jejuni of Multilocus Sequence Type Clonal Complexes 21 and 45 under Freeze, Chill, Oxidative, Acid and Heat Stresses. Food Microbiol. 2010, 27, 829–834. [Google Scholar] [CrossRef]
- Elhadidy, M.; Arguello, H.; Álvarez-Ordóñez, A.; Miller, W.G.; Duarte, A.; Martiny, D.; Hallin, M.; Vandenberg, O.; Dierick, K.; Botteldoorn, N. Orthogonal Typing Methods Identify Genetic Diversity among Belgian Campylobacter jejuni Strains Isolated over a Decade from Poultry and Cases of Sporadic Human Illness. Int. J. Food Microbiol. 2018, 275, 66–75. [Google Scholar] [CrossRef]
- Bailey, R.A.; Kranis, A.; Psifidi, A.; Watson, K.A.; Rothwell, L.; Hocking, P.M.; Kaiser, P.; Stevens, M.P.; Avendano, S. Colonization of a Commercial Broiler Line by Campylobacter Is under Limited Genetic Control and Does Not Significantly Impair Performance or Intestinal Health. Poult. Sci. 2018, 97, 4167–4176. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time of Inoculation | Challenge Dose in log10 cfu | Sample Size per Group (n) |
|---|---|---|
| Week 1 * | 3.0 | Bacterin, subunit, control (n = 17) |
| 4.0 | Bacterin, subunit, control (n = 17) | |
| Week 2 * | 2.9 | Bacterin, subunit, control (n = 17) |
| 3.9 | Bacterin, subunit, control (n = 17) | |
| Week 2 ‡ | 2.7 | Bacterin, subunit, control (n = 19) |
| 3.6 | Bacterin, subunit, control (n = 14) | |
| Week 3 ‡ | 2.8 | Bacterin, subunit, control (n = 19) |
| 3.7 | Bacterin, subunit, control (n = 14) |
| MAB− | |||||||
|---|---|---|---|---|---|---|---|
| Breeder Flock | Species | CC | ST | Broiler Flock | Species | CC | ST |
| D (Pooled) | C. coli | UA * | 6549 | D1 | C. jejuni | 21 | 21 |
| E | No permission granted | E1 | C. jejuni | UA | 2274 | ||
| E2 | C. coli | UA | 10,044 | ||||
| E3 | Negative | ||||||
| E4 | Negative | ||||||
| P (Stable 1) | C. coli | Not analyzed * | P1 | Negative | |||
| P (Stable 2) | C. coli | Not analyzed | P2 | Negative | |||
| P3 | Negative | ||||||
| P4 | Negative | ||||||
| U (Stable 1) | C. coli | 828 | UA | U1 | C. coli | 828 | 832 |
| U (Stable 2) | C. coli | 828 | 828 | ||||
| MAB+ | |||||||
| Breeder Flock | Species | CC | ST | Broiler Flock | Species | CC | ST |
| J (Pooled) | C. coli | Not analyzed | J1 | Negative | |||
| J2 | Negative | ||||||
| W (Stable 1) | C. coli | 828 | 1058 | W1 | C. coli | 828 | 832 |
| W (Stable 2) | C. coli | 828 | 828 | W2 | C. jejuni | 21 | 50 |
| W (Stable 3) | C. coli | 828 | 828 | W3 | Negative | ||
| W4 | Negative | ||||||
| W5 | Negative | ||||||
| W6 | Negative | ||||||
| W7 | Negative | ||||||
| W8 | Negative | ||||||
| Group | Inoculation Dose (log10 cfu) | KC40 Whole-Cell Coating | Subunit Coating | ||
|---|---|---|---|---|---|
| Bacterin | Control | Subunit | Control | ||
| Week 1 * | 3.0 | 1:16,000 | 1:400 | 1:32,000 | Negative |
| 4.0 | 1:16,000 | 1:800 | 1:64,000 | Negative | |
| Week 2 * | 2.9 | 1:8000 | Negative | 1:25,600 | Negative |
| 3.9 | 1:8000 | Negative | 1:12,800 | Negative | |
| Week 2 ‡ | 2.7 | 1:1600 | Negative | 1:6400 | Negative |
| 3.6 | 1:1600 | 1:400 | 1:6400 | Negative | |
| Week 3 ‡ | 2.8 | 1:400 | Negative | 1:3200 | Negative |
| 3.7 | 1:800 | Negative | 1:1600 | Negative | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haems, K.; Strubbe, D.; Van Rysselberghe, N.; Rasschaert, G.; Martel, A.; Pasmans, F.; Garmyn, A. Role of Maternal Antibodies in the Protection of Broiler Chicks against Campylobacter Colonization in the First Weeks of Life. Animals 2024, 14, 1291. https://doi.org/10.3390/ani14091291
Haems K, Strubbe D, Van Rysselberghe N, Rasschaert G, Martel A, Pasmans F, Garmyn A. Role of Maternal Antibodies in the Protection of Broiler Chicks against Campylobacter Colonization in the First Weeks of Life. Animals. 2024; 14(9):1291. https://doi.org/10.3390/ani14091291
Chicago/Turabian StyleHaems, Kristof, Diederik Strubbe, Nathalie Van Rysselberghe, Geertrui Rasschaert, An Martel, Frank Pasmans, and An Garmyn. 2024. "Role of Maternal Antibodies in the Protection of Broiler Chicks against Campylobacter Colonization in the First Weeks of Life" Animals 14, no. 9: 1291. https://doi.org/10.3390/ani14091291
APA StyleHaems, K., Strubbe, D., Van Rysselberghe, N., Rasschaert, G., Martel, A., Pasmans, F., & Garmyn, A. (2024). Role of Maternal Antibodies in the Protection of Broiler Chicks against Campylobacter Colonization in the First Weeks of Life. Animals, 14(9), 1291. https://doi.org/10.3390/ani14091291