
Phenotypic and Functional Characterization of Bovine Adipose-Derived Mesenchymal Stromal Cells
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
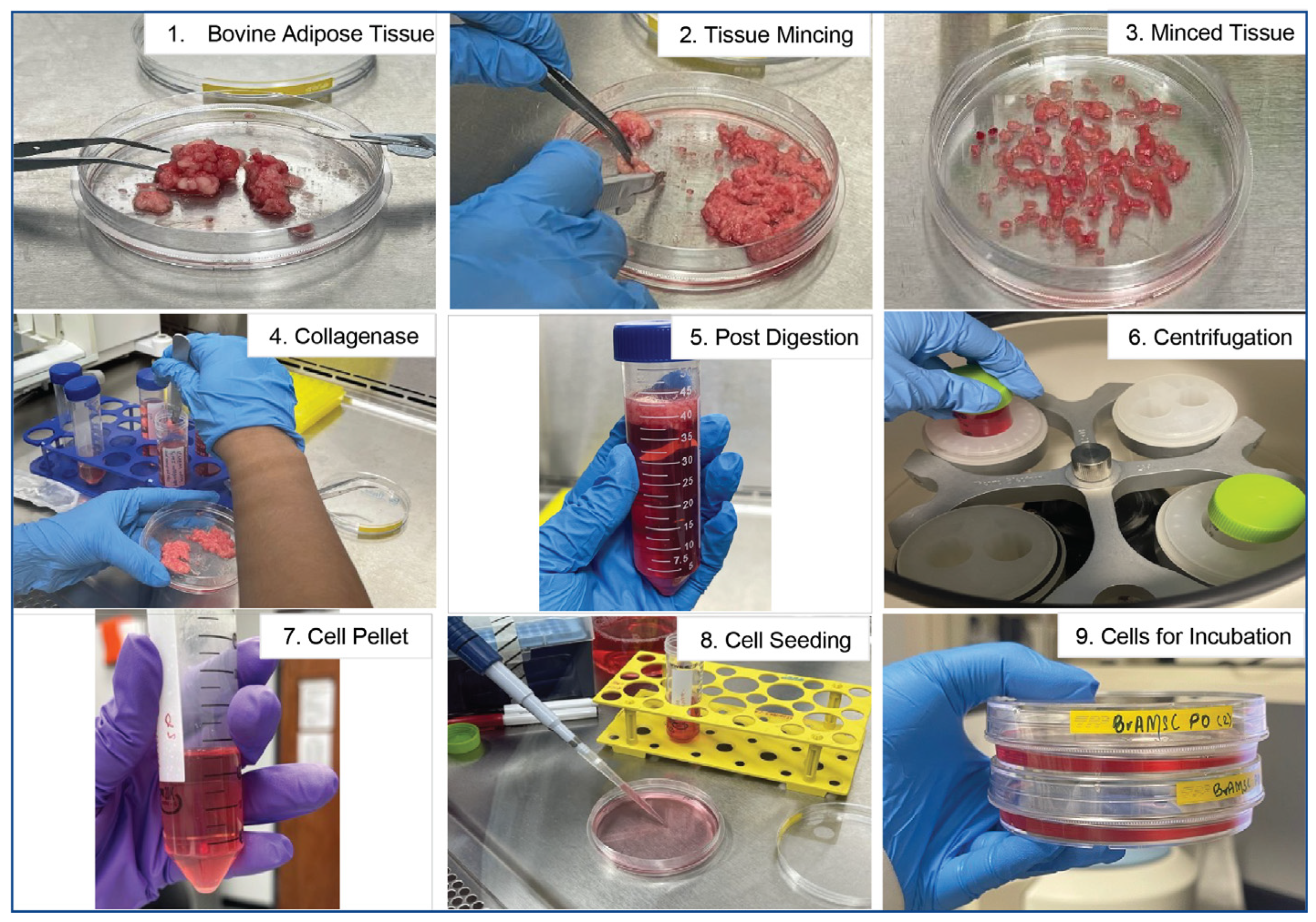
2.1. Collection of Adipose (Fat) Tissue from Cows
2.2. Isolation of Bovine Adipose-Derived Mesenchymal Stem Cells (BvAdMSCs)
2.3. Cell Culture and Maintenance
2.4. Morphological Characterization
2.5. Colony Forming Unit–Fibroblastic Assay
2.6. Proliferation Assay
2.6.1. Growth Assay
2.6.2. Cell Counting Kit-8 (CCK-8) Assay
2.7. In-Vitro Osteogenic and Adipogenic Differentiation
2.8. Surface Marker Characterization
2.8.1. Microscopic Imaging
2.8.2. Flow Cytometric Analysis
2.9. Indoleamine 2,3-Dioxygenase (IDO) Activity Assay
2.10. Statistical Analyses
3. Results
3.1. Bovine Adipose Tissue Yields Self-Renewing and Proliferative Stromal Cells
3.2. Bovine Adipose MSCs Express Characteristic Cell Surface Markers
3.3. Bovine Adipose MSCs Differentiate into Osteo and Adipogenic Lineages
3.4. Bovine Adipose MSCs Have Inducible IDO Activity and Immunosuppressive Potential
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPs | Antimicrobial peptides |
| BvAdMSCs | Bovine Adipose-derived Mesenchymal Stem Cells |
| CCK-8 | Cell Counting Kit-8 |
| CD | Cluster of Differentiation |
| CFU–F | Colony Forming Unit–Fibroblastic |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DMEM | Dulbecco’s Modified Eagle Medium |
| EVOS | EVOS XL CORE imaging system |
| FBS | Fetal Bovine Serum |
| hBM-MSCs | Human Bone Marrow-derived Mesenchymal Stem Cells |
| IACUC | Institutional Animal Care and Use Committee |
| IDO | Indoleamine 2,3 dioxygenase |
| IFN-γ | Interferon gamma |
| IgG | Immunoglobulin G |
| IL-10 | Interleukin |
| MEM α | Alpha Minimum Essential Medium |
| MSC | Mesenchymal Stem Cells |
| P/S | Penicillin–Streptomycin |
| PBS | Phosphate Buffered Saline |
| PFA | Paraformaldehyde |
| PGE2 | Prostaglandin E2 |
| TGFβ1 | Transforming growth factor-beta 1 |
References
- Food Outlook; Food and Agriculture Organization of the United Nations: Rome, Italy, 2017.
- Dairy Market Review; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018.
- Duff, G.C.; Galyean, M.L. Board-invited review: Recent advances in management of highly stressed, newly received feedlot cattle. J. Anim. Sci. 2007, 85, 823–840. [Google Scholar] [CrossRef]
- McGill, J.L.; Sacco, R.E. The Immunology of Bovine Respiratory Disease: Recent Advancements. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 333–348. [Google Scholar] [CrossRef]
- Perry, B.D.; Grace, D.; Sones, K. Current drivers and future directions of global livestock disease dynamics. Proc. Natl. Acad. Sci. USA 2013, 110, 20871–20877. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Medicinal signalling cells: They work, so use them. Nature 2019, 566, 39. [Google Scholar] [CrossRef]
- Chow, L.; Johnson, V.; Impastato, R.; Coy, J.; Strumpf, A.; Dow, S. Antibacterial activity of human mesenchymal stem cells mediated directly by constitutively secreted factors and indirectly by activation of innate immune effector cells. Stem Cells Transl. Med. 2020, 9, 235–249. [Google Scholar] [CrossRef]
- Cahuascanco, B.; Bahamonde, J.; Huaman, O.; Jervis, M.; Cortez, J.; Palomino, J.; Escobar, A.; Retamal, P.; Torres, C.G.; Peralta, O.A. Bovine fetal mesenchymal stem cells exert antiproliferative effect against mastitis causing pathogen Staphylococcus aureus. Vet. Res. 2019, 50, 25. [Google Scholar] [CrossRef] [PubMed]
- Peralta, O.A.; Carrasco, C.; Vieytes, C.; Tamayo, M.J.; Muñoz, I.; Sepulveda, S.; Tadich, T.; Duchens, M.; Melendez, P.; Mella, A.; et al. Safety and efficacy of a mesenchymal stem cell intramammary therapy in dairy cows with experimentally induced Staphylococcus aureus clinical mastitis. Sci. Rep. 2020, 10, 2843. [Google Scholar] [CrossRef] [PubMed]
- Grayson, W.L.; Bunnell, B.A.; Martin, E.; Frazier, T.; Hung, B.P.; Gimble, J.M. Stromal cells and stem cells in clinical bone regeneration. Nat. Rev. Endocrinol. 2015, 11, 140–150. [Google Scholar] [CrossRef]
- Alcayaga-Miranda, F.; Cuenca, J.; Khoury, M. Antimicrobial Activity of Mesenchymal Stem Cells: Current Status and New Perspectives of Antimicrobial Peptide-Based Therapies. Front. Immunol. 2017, 8, 339. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise Review: Multifaceted Characterization of Human Mesenchymal Stem Cells for Use in Regenerative Medicine. Stem Cells Transl. Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef]
- Alipour, R.; Masoumi Karimi, M.; Hashemi-Beni, B.; Adib, M.; Sereshki, N.; Sadeghi, F. Indoleamine 2,3-Dioxygenase Is Dispensable for The Immunomodulatory Function of Stem Cells from Human Exfoliated Deciduous Teeth. Cell J. 2017, 18, 597–608. [Google Scholar] [PubMed]
- Zheng, G.; Qiu, G.; Ge, M.; He, J.; Huang, L.; Chen, P.; Wang, W.; Xu, Q.; Hu, Y.; Shu, Q.; et al. Human adipose-derived mesenchymal stem cells alleviate obliterative bronchiolitis in a murine model via IDO. Respir. Res. 2017, 18, 119. [Google Scholar] [CrossRef]
- Wu, H.; Gong, J.; Liu, Y. Indoleamine 2,3-dioxygenase regulation of immune response (Review). Mol. Med. Rep. 2018, 17, 4867–4873. [Google Scholar] [PubMed]
- Kelly, K.; Rasko, J.E.J. Mesenchymal Stromal Cells for the Treatment of Graft Versus Host Disease. Front. Immunol. 2021, 12, 761616. [Google Scholar] [CrossRef]
- Gugliandolo, A.; Bramanti, P.; Mazzon, E. Mesenchymal Stem Cells: A Potential Therapeutic Approach for Amyotrophic Lateral Sclerosis? Stem Cells Int. 2019, 2019, 3675627. [Google Scholar] [CrossRef]
- Li, W.; Shi, L.; Hu, B.; Hong, Y.; Zhang, H.; Li, X.; Zhang, Y. Mesenchymal Stem Cell-Based Therapy for Stroke: Current Understanding and Challenges. Front. Cell. Neurosci. 2021, 15, 628940. [Google Scholar] [CrossRef]
- Lopez-Santalla, M.; Bueren, J.A.; Garin, M.I. Mesenchymal stem/stromal cell-based therapy for the treatment of rheumatoid arthritis: An update on preclinical studies. eBioMedicine 2021, 69, 103427. [Google Scholar] [CrossRef]
- Shi, L.; Yuan, X.; Yao, W.; Wang, S.; Zhang, C.; Zhang, B.; Song, J.; Huang, L.; Xu, Z.; Fu, J.-L.; et al. Human mesenchymal stem cells treatment for severe COVID-19: 1-year follow-up results of a randomized, double-blind, placebo-controlled trial. eBioMedicine 2022, 75, 103789. [Google Scholar] [CrossRef]
- Rider, D.A.; Dombrowski, C.; Sawyer, A.A.; Ng, G.H.B.; Leong, D.; Hutmacher, D.W.; Nurcombe, V.; Cool, S.M. Autocrine fibroblast growth factor 2 increases the multipotentiality of human adipose-derived mesenchymal stem cells. Stem Cells 2008, 26, 1598–1608. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Dudakovic, A.; Manzar, B.; Sen, B.; Dietz, A.B.; Cool, S.M.; Rubin, J.; van Wijnen, A.J. Osteogenic Stimulation of Human Adipose-Derived Mesenchymal Stem Cells Using a Fungal Metabolite That Suppresses the Polycomb Group Protein EZH2. Stem Cells Transl. Med. 2018, 7, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Samsonraj, R.M.; Raghunath, M.; Hui, J.H.; Ling, L.; Nurcombe, V.; Cool, S.M. Telomere length analysis of human mesenchymal stem cells by quantitative PCR. Gene 2013, 519, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Samsonraj, R.M.; Rai, B.; Sathiyanathan, P.; Puan, K.J.; Rötzschke, O.; Hui, J.H.; Raghunath, M.; Stanton, L.W.; Nurcombe, V.; Cool, S.M. Establishing criteria for human mesenchymal stem cell potency. Stem Cells 2015, 33, 1878–1891. [Google Scholar] [CrossRef] [PubMed]
- Ferragut, F.; Vachetta, V.S.; Troncoso, M.F.; Rabinovich, G.A.; Elola, M.T. ALCAM/CD166: A pleiotropic mediator of cell adhesion, stemness and cancer progression. Cytokine Growth Factor Rev. 2021, 61, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Senbanjo, L.T.; Chellaiah, M.A. CD44: A Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front. Cell Dev. Biol. 2017, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Secunda, R.; Vennila, R.; Mohanashankar, A.M.; Rajasundari, M.; Jeswanth, S.; Surendran, R. Isolation, expansion and characterisation of mesenchymal stem cells from human bone marrow, adipose tissue, umbilical cord blood and matrix: A comparative study. Cytotechnology 2015, 67, 793–807. [Google Scholar] [CrossRef] [PubMed]
- Ghaneialvar, H.; Soltani, L.; Rahmani, H.R.; Lotfi, A.S.; Soleimani, M. Characterization and Classification of Mesenchymal Stem Cells in Several Species Using Surface Markers for Cell Therapy Purposes. Indian J. Clin. Biochem. 2018, 33, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Moraes, D.A.; Sibov, T.T.; Pavon, L.F.; Alvim, P.Q.; Bonadio, R.S.; Da Silva, J.R.; Pic-Taylor, A.; Toledo, O.A.; Marti, L.C.; Azevedo, R.B.; et al. A reduction in CD90 (THY-1) expression results in increased differentiation of mesenchymal stromal cells. Stem Cell Res. Ther. 2016, 7, 97. [Google Scholar] [CrossRef]
- Yeola, A.; Subramanian, S.; Oliver, R.A.; Lucas, C.A.; Thoms, J.A.I.; Yan, F.; Olivier, J.; Chacon, D.; Tursky, M.L.; Srivastava, P.; et al. Induction of muscle-regenerative multipotent stem cells from human adipocytes by PDGF-AB and 5-azacytidine. Sci. Adv. 2021, 7, eabd1929. [Google Scholar] [CrossRef]
- Nakano, A.; Harada, T.; Morikawa, S.; Kato, Y. Expression of Leukocyte Common Antigen (CD45) on Various Human Leukemia/Lymphoma Cell Lines. Pathol. Int. 1990, 40, 107–115. [Google Scholar] [CrossRef]
- Kumar, S.; Rajkumar, S.V.; Kimlinger, T.; Greipp, P.R.; Witzig, T.E. CD45 expression by bone marrow plasma cells in multiple myeloma: Clinical and biological correlations. Leukemia 2005, 19, 1466–1470. [Google Scholar] [CrossRef]
- Alzheimer’s Association: 2016 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2016, 12, 459–509. [CrossRef]
- Burrow, K.L.; Hoyland, J.A.; Richardson, S.M. Human Adipose-Derived Stem Cells Exhibit Enhanced Proliferative Capacity and Retain Multipotency Longer than Donor-Matched Bone Marrow Mesenchymal Stem Cells during Expansion In Vitro. Stem Cells Int. 2017, 2017, 2541275. [Google Scholar] [CrossRef]
- Mohamed-Ahmed, S.; Fristad, I.; Lie, S.A.; Suliman, S.; Mustafa, K.; Vindenes, H.; Idris, S.B. Adipose-derived and bone marrow mesenchymal stem cells: A donor-matched comparison. Stem Cell Res. Ther. 2018, 9, 168. [Google Scholar] [CrossRef]
- Sathiyanathan, P.; Samsonraj, R.M.; Tan, C.L.L.; Ling, L.; Lezhava, A.; Nurcombe, V.; Stanton, L.W.; Cool, S.M. A genomic biomarker that identifies human bone marrow-derived mesenchymal stem cells with high scalability. Stem Cells 2020, 38, 1124–1136. [Google Scholar] [CrossRef]
- Hill, A.B.T.; Bressan, F.F.; Murphy, B.D.; Garcia, J.M. Applications of mesenchymal stem cell technology in bovine species. Stem Cell Res. Ther. 2019, 10, 44. [Google Scholar] [CrossRef]
- Oyarzo, R.; Valderrama, X.; Valenzuela, F.; Bahamonde, J. Bovine Fetal Mesenchymal Stem Cells Obtained from Omental Adipose Tissue and Placenta Are More Resistant to Cryoprotectant Exposure Than Those from Bone Marrow. Front. Vet. Sci. 2021, 8, 708972. [Google Scholar] [CrossRef]
- Ozden Akkaya, O.; Dikmen, T.; Nawaz, S.; Kibria, A.G.; Altunbaş, K.; Yağci, A.; Erdoğan, M.; Yaprakci, M.V. Comparison of proliferation and osteogenic differentiation potential of bovine adipose tissue and bone marrow derived stem cells. Biotech. Histochem. 2023, 98, 267–279. [Google Scholar] [CrossRef]
- Xiong, H.; Bai, C.; Wu, S.; Gao, Y.; Lu, T.; Hu, Q.; Guan, W.; Ma, Y. Biological characterization of mesenchymal stem cells from bovine umbilical cord. Anim. Cells Syst. 2014, 18, 59–67. [Google Scholar] [CrossRef]
- Noronha, N.d.C.; Mizukami, A.; Caliári-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Res. Ther. 2019, 10, 131. [Google Scholar] [CrossRef]
- Wang, G.; Cao, K.; Liu, K.; Xue, Y.; Roberts, A.I.; Li, F.; Han, Y.; Rabson, A.B.; Wang, Y.; Shi, Y. Kynurenic acid, an IDO metabolite, controls TSG-6-mediatedimmunosuppression of human mesenchymal stem cells. Cell Death Differ. 2018, 25, 1209–1223. [Google Scholar] [CrossRef]
- Wi, H.; Lee, S.; Kim, Y.; No, J.-G.; Lee, P.; Lee, B.R.; Oh, K.B.; Hur, T.; Ock, S.A. Immunosuppression-enhancing effect of the administration of allogeneic canine adipose-derived mesenchymal stem cells (cA-MSCs) compared with autologous cA-MSCs in vitro. J. Vet. Sci. 2021, 22, e63. [Google Scholar] [CrossRef]
- English, K.; Barry, F.P.; Field-Corbett, C.P.; Mahon, B.P. IFN-γ and TNF-α differentially regulate immunomodulation by murine mesenchymal stem cells. Immunol. Lett. 2007, 110, 91–100. [Google Scholar] [CrossRef]
- Yoo, K.H.; Jang, I.K.; Lee, M.W.; Kim, H.E.; Yang, M.S.; Eom, Y.; Lee, J.E.; Kim, Y.J.; Yang, S.K.; Jung, H.L.; et al. Comparison of immunomodulatory properties of mesenchymal stem cells derived from adult human tissues. Cell. Immunol. 2009, 259, 150–156. [Google Scholar] [CrossRef]
- Chang, C.; Yen, M.; Chen, Y.; Chien, C.; Huang, H.; Bai, C.; Yen, B.L. Placenta-Derived Multipotent Cells Exhibit Immunosuppressive Properties That Are Enhanced in the Presence of Interferon-γ. Stem Cells 2006, 24, 2466–2477. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maldonado, V.V.; Pokharel, S.; Powell, J.G.; Samsonraj, R.M. Phenotypic and Functional Characterization of Bovine Adipose-Derived Mesenchymal Stromal Cells. Animals 2024, 14, 1292. https://doi.org/10.3390/ani14091292
Maldonado VV, Pokharel S, Powell JG, Samsonraj RM. Phenotypic and Functional Characterization of Bovine Adipose-Derived Mesenchymal Stromal Cells. Animals. 2024; 14(9):1292. https://doi.org/10.3390/ani14091292
Chicago/Turabian StyleMaldonado, Vitali V., Sriya Pokharel, Jeremy G. Powell, and Rebekah M. Samsonraj. 2024. "Phenotypic and Functional Characterization of Bovine Adipose-Derived Mesenchymal Stromal Cells" Animals 14, no. 9: 1292. https://doi.org/10.3390/ani14091292
APA StyleMaldonado, V. V., Pokharel, S., Powell, J. G., & Samsonraj, R. M. (2024). Phenotypic and Functional Characterization of Bovine Adipose-Derived Mesenchymal Stromal Cells. Animals, 14(9), 1292. https://doi.org/10.3390/ani14091292