Mammary Defences and Immunity against Mastitis in Sheep

 ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
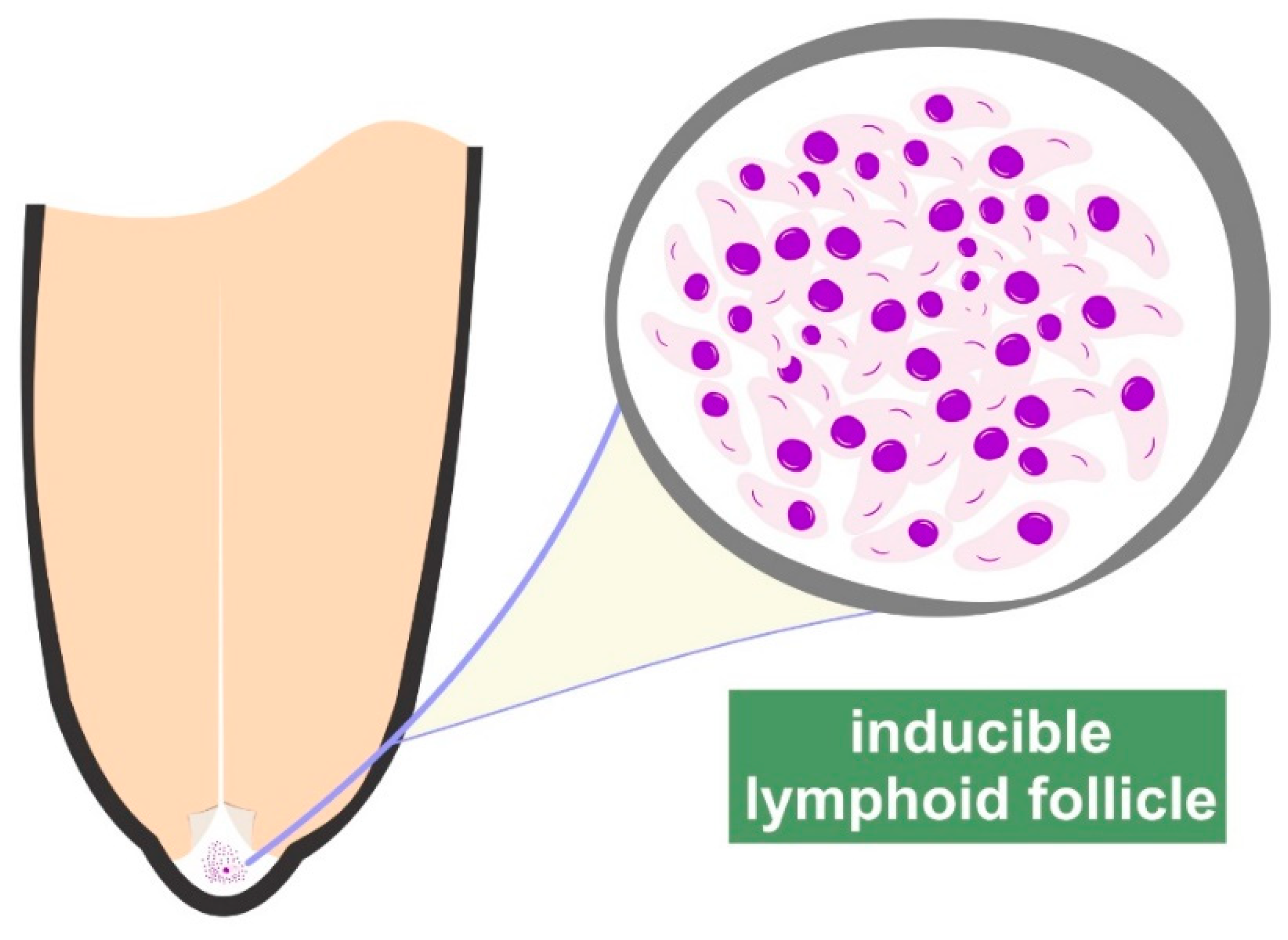
2. The Teat of the Udder
3. Cells in the Mammary Gland
3.1. Mammary Epithelial Cells
3.2. Leucocytes
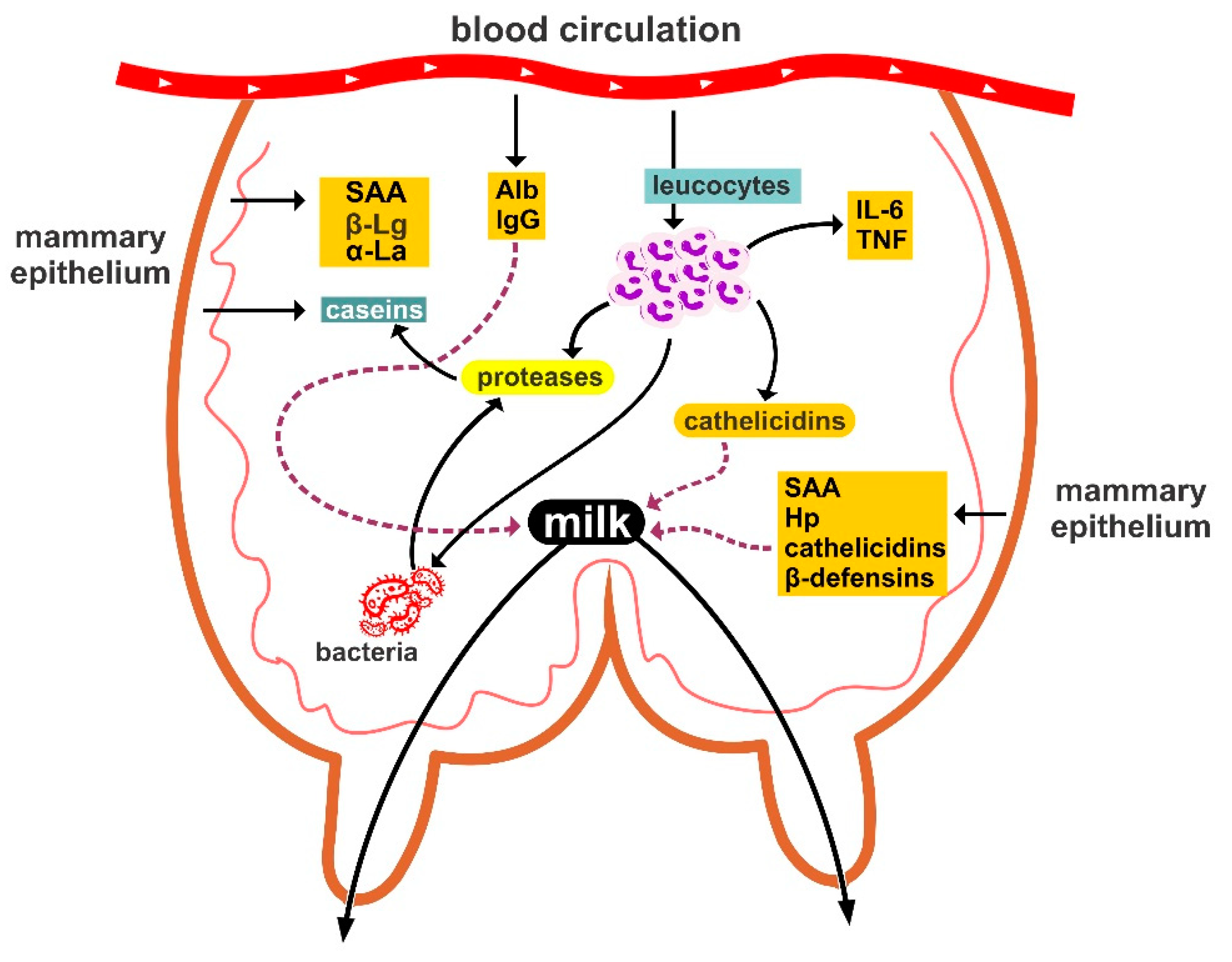
4. Immunoglobulins and Complement System
4.1. Immunoglobulins
4.2. Complement System
5. Chemical Antibacterial Agents
5.1. Lactoferrin
5.2. Lysozyme
5.3. Lactoperoxidase
6. Modulation of Mammary Defences
6.1. Factors Affecting Mammary Defences
6.2. Immunisation
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rainard, P.; Riollet, C. Innate immunity of the bovine mammary gland. Vet. Res. 2006, 37, 369–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickerson, S.C. Mammary resistance mechanisms/anatomical. In Encyclopedia of Dairy Sciences; Roginski, H., Fuquay, J.W., Fox, P.F., Eds.; Academic Press: London, UK, 2002; pp. 1697–1701. [Google Scholar]
- Mavrogianni, V.S.; Fthenakis, G.C.; Brooks, H.; Papaioannou, N.; Cripps, P.J.; Taitzoglou, I.; Brellou, G.; Saratsis, P. The effects of inoculation of Mannheimia haemolytica into the teat of lactating ewes. Vet. Res. 2005, 36, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Alnakip, M.E.; Quintela-Baluja, M.; Böhme, K.; Fernández-No, I.; Caamaño-Antelo, S.; Calo-Mata, P.; Barros-Velázquez, J. The immunology of mammary gland of dairy ruminants between healthy and inflammatory conditions. J. Vet. Med. 2014, 31, 659801. [Google Scholar] [CrossRef] [PubMed]
- Peeler, E.J.; Green, M.J.; Fitzpatrick, J.L.; Morgan, K.L.; Green, L.E. Risk factors associated with clinical mastitis in low somatic cell count British dairy herds. J. Dairy Sci. 2000, 83, 2464–2472. [Google Scholar] [CrossRef]
- DeVries, T.J.; Dufour, S.; Scholl, D.T. Relationship between feeding strategy, lying behavior patterns, and incidence of intramammary infection in dairy cows. J. Dairy Sci. 2010, 93, 1987–1997. [Google Scholar] [CrossRef]
- McDonald, J. Radiographic method for anatomic study of the teat canal: Changes between milking periods. Am. J. Vet. Res. 1975, 36, 1241–1242. [Google Scholar] [PubMed]
- Schultze, W.D.; Bright, S.C. Changes in penetrability of bovine papillary duct to endotoxin after milking. Am. J. Vet. Res. 1983, 44, 2373–2375. [Google Scholar] [PubMed]
- Alejandro, M.; Roca, A.; Romero, G.; Díaz, J.R. How does the milk removal method affect teat tissue and teat recovery in dairy ewes? J. Dairy Res. 2014, 81, 350–357. [Google Scholar] [CrossRef]
- Fragkou, I.A.; Dagleish, M.P.; Papaioannou, N.; Cripps, P.J.; Boscos, C.M.; Ververidis, H.N.; Orfanou, D.C.; Solomakos, N.; Finlayson, J.; Govaris, A.; et al. The induction of lymphoid follicle-like structures in the ovine teat duct following experimental infection with Mannheimia haemolytica. Vet. J. 2010, 184, 194–200. [Google Scholar] [CrossRef]
- Ferreras-Estrada, M.C.; Campo, R.; González-Lanza, C.; Pérez, V.; García-Marín, J.F.; Manga-González, M.Y. Immunohistochemical study of the local immune response in lambs experimentally infected with Dicrocoelium dendriticum (Digenea). Parasitol. Res. 2007, 101, 547–555. [Google Scholar] [CrossRef]
- Fragkou, I.A.; Mavrogianni, V.S.; Papaioannou, N.; Boscos, C.; Cripps, P.J.; Skoufos, J.; Fthenakis, G.C. Presence of sub-epithelial lymphoid tissues in the teat of ewe-lambs and adult ewes. Small Rumin. Res. 2007, 70, 286–291. [Google Scholar] [CrossRef]
- Rainard, P. Mammary microbiota of dairy ruminants: Fact or fiction? Vet. Res. 2017, 48, 25. [Google Scholar] [CrossRef] [PubMed]
- Vasileiou, N.G.C.; Cripps, P.J.; Ioannidi, K.S.; Chatzopoulos, D.C.; Gougoulis, D.A.; Sarrou, S.; Orfanou, D.C.; Politis, A.; Calvo Gonzalez-Valerio, T.; Argyros, S.; et al. Extensive countrywide field investigation of subclinical mastitis in sheep in Greece. J. Dairy Sci. 2018, 101, 7297–7310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhoeven, P.O.; Gagnaire, J.; Botelho-Nevers, E.; Grattard, F.; Carricajo, A.; Lucht, F.; Pozzetto, B.; Berthelot, P. Detection and clinical relevance of Staphylococcus aureus nasal carriage: An update. Exp. Rev. Anti-Infect. Ther. 2014, 12, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Vasileiou, N.G.C.; Chatzopoulos, D.C.; Sarrou, S.; Fragkou, A.I.; Katsafadou, A.I.; Mavrogianni, V.S.; Petinaki, E.; Fthenakis, G.C. Role of staphylococci in mastitis in sheep. J. Dairy Res. 2019, 86, 254–266. [Google Scholar] [CrossRef] [Green Version]
- Fragkou, I.A.; Mavrogianni, V.S.; Cripps, P.J.; Gougoulis, D.A.; Fthenakis, G.C. The bacterial flora in the teat duct of ewes can protect against and can cause mastitis. Vet. Res. 2007, 38, 525–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brook, I. The role of bacterial interference in otitis, sinusitis and tonsillitis. Otolaryngol. Head Neck Surg. 2005, 133, 139–146. [Google Scholar] [CrossRef]
- Melchior, M.B.; Vaarkamp, H.; Fink-Gremmels, J. Biofilms: A role in recurrent mastitis infections? Vet. J. 2006, 171, 398–407. [Google Scholar] [CrossRef]
- Vilela, C.L.; Fitzpatrick, J.; Morgan, K.L. In vitro adherence and invasion of ovine mammary epithelium by Mannheimia haemolytica. Vet. J. 2004, 167, 211–213. [Google Scholar] [CrossRef]
- Isenberg, H.D.; D’Amato, R.F. Indigenous and pathogenic microorganisms of humans. In Manual of Clinical Microbiology; Lennette, E.H., Balows, H., Hausler, W.J., Jr., Shadomy, H.J., Eds.; American Society for Microbiology: Washington, DC, USA, 1985; pp. 24–35. [Google Scholar]
- Smith, H. The revival of interest in mechanisms of bacterial pathogenicity. Biol. Rev. 1995, 70, 277–316. [Google Scholar] [CrossRef]
- De Vliegher, S.; Opsomer, G.; Vanrolleghem, A.; Devriese, L.A.; Sampimon, O.C.; Sol, J.; Barkema, H.W.; Haesebrouck, F.; de Kruif, A. In vitro growth inhibition of major mastitis pathogens by Staphylococcus chromogenes originating from teat apices of dairy heifers. Vet. Microbiol. 2004, 101, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Rainard, P.; Poutrel, B. Effect of naturally occurring intramammary infections by minor pathogens on new infections by major pathogens in cattle. Am. J. Vet. Res. 1988, 49, 327–329. [Google Scholar] [PubMed]
- Targowski, S.P. Role of immune factors in protection of mammary gland. J. Dairy Sci. 1983, 6, 1781–1789. [Google Scholar] [CrossRef]
- Günther, J.; Seyfert, H.M. The first line of defence: Insights into mechanisms and relevance of phagocytosis in epithelial cells. Semin. Immunopathol. 2018, 40, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Petzl, W.; Zerbe, H.; Günther, J.; Yang, W.; Seyfert, H.M.; Nürnberg, G.; Schuberth, H.J. Escherichia coli, but not Staphylococcus aureus triggers an early increased expression of factors contributing to the innate immune defense in the udder of the cow. Vet. Res. 2008, 39, 18. [Google Scholar] [CrossRef] [PubMed]
- Pauwels, A.M.; Trost, M.; Beyaert, R.; Hoffmann, E. Patterns, receptors, and signals: Regulation of phagosome maturation. Trends Immunol. 2017, 38, 407–422. [Google Scholar] [CrossRef] [PubMed]
- Katsafadou, A.I. Proteomic Study of Ovine Mastitis associated with Mannheimia haemolytica. Ph.D. Thesis, University of Thessaly, Karditsa, Greece, 2017. [Google Scholar]
- Gao, J.; Li, T.; Lu, Z.; Wang, X.; Zhao, X.; Ma, Y. Proteomic analyses of mammary glands provide insight into the immunity and metabolism pathways associated with clinical mastitis in meat sheep. Animals 2019, 9, 309. [Google Scholar] [CrossRef]
- Katsafadou, A.I.; Tsangaris, G.T.; Anagnostopoulos, A.K.; Billinis, C.; Barbagianni, M.S.; Vasileiou, N.G.C.; Spanos, S.A.; Mavrogianni, V.S.; Fthenakis, G.C. Differential quantitative proteomics study of experimental Mannheimia haemolytica mastitis in sheep. J. Proteom. 2019, 205, 103393. [Google Scholar] [CrossRef]
- Bauer, I.; Günther, J.; Wheeler, T.T.; Engelmann, S.; Seyfert, H.M. Extracellular milieu grossly alters pathogen-specific immune response of mammary epithelial cells. BMC Vet. Res. 2015, 11, 172. [Google Scholar] [CrossRef]
- Günther, J.; Czabanska, A.; Bauer, I.; Leigh, J.A.; Holst, O.; Seyfert, H.M. Streptococcus uberis strains isolated from the bovine mammary gland evade immune recognition by mammary epithelial cells, but not of macrophages. Vet. Res. 2016, 47, 13. [Google Scholar] [CrossRef]
- Lahouassa, H.; Moussay, E.; Rainard, P.; Riollet, C. Differential cytokine and chemokine responses of bovine mammary epithelial cells to Staphylococcus aureus and Escherichia coli. Cytokine 2007, 38, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.M.; Stelwagen, K.; Dobson, J.; Henderson, H.V.; Davis, S.R.; Farr, V.C.; Singh, K. Transcriptome profiling of Streptococcus uberis-induced mastitis reveals fundamental differences between immune gene expression in the mammary gland and in a primary cell culture model. J. Dairy Sci. 2009, 92, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Brenaut, P.; Lefevre, L.; Rau, A.; Laloe, D.; Pisoni, G.; Moroni, P.; Bevilacqua, C.; Martin, P. Contribution of mammary epithelial cells to the immune response during early stages of a bacterial infection to Staphylococcus aureus. Vet. Res. 2014, 45, 16. [Google Scholar] [CrossRef] [PubMed]
- Eckersall, P.D.; Young, F.J.; Nolan, A.M.; Knight, C.H.; McComb, C.; Waterston, M.M.; Hogarth, C.J.; Scott, E.M.; Fitzpatrick, J.L. Acute phase proteins in bovine milk in an experimental model of Staphylococcus aureus subclinical mastitis. J. Dairy Sci. 2006, 89, 488–501. [Google Scholar] [CrossRef]
- Eckersall, P.D.; Young, F.J.; McComb, C.; Hogarth, C.J.; Safi, S.; Weber, A.; McDonald, T.; Nolan, A.M.; Fitzpatrick, J.L. Acute phase proteins in serum and milk from dairy cows with clinical mastitis. Vet. Rec. 2001, 148, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Smolenski, G.; Haines, S.; Kwan, F.Y.S.; Bond, J.; Farr, V.; Davis, S.R.; Stelwagen, Κ.; Wheeler, Τ.Τ. Characterisation of host defense proteins in milk using a proteomic approach. J. Proteome Res. 2007, 6, 207–215. [Google Scholar] [CrossRef] [PubMed]
- McDonald, T.L.; Larson, M.A.; Mack, D.R.; Weber, A. Elevated extrahepatic expression and secretion of mammary associated serum amyloid A 3 (M-SAA3) into colostrum. Vet. Immunol. Immunopathol. 2001, 83, 203–211. [Google Scholar] [CrossRef]
- Cubeddu, T.; Cacciotto, C.; Pisanu, S.; Tedde, V.; Alberti, A.; Pittau, M.; Dore, S.; Cannas, A.; Uzzau, S.; Rocca, S.; et al. Cathelicidin production and release by mammary epithelial cells during infectious mastitis. Vet. Immunol. Immunopathol. 2017, 89, 66–70. [Google Scholar] [CrossRef]
- West, M.A.; Heagy, W. Endotoxin tolerance: A review. Crit. Care Med. 2002, 30, S64–S73. [Google Scholar] [CrossRef]
- Broad, A.; Jones, D.E.; Kirby, J.A. Toll-like receptor (TLR) response tolerance: A key physiological “damage limitation” effect and an important potential opportunity for therapy. Curr. Med. Chem. 2006, 13, 2487–2502. [Google Scholar] [CrossRef]
- Hodgson, J.C. Endotoxin and mammalian host responses during experimental disease. J. Comp. Pathol. 2006, 135, 157–175. [Google Scholar] [CrossRef] [PubMed]
- Petzl, W.; Günther, J.; Pfister, T.; Sauter-Louis, C.; Goetze, L.; von Aulock, S.; Hafner-Marx, A.; Schuberth, H.J.; Seyfert, H.M.; Zerbe, H. Lipopolysaccharide pretreatment of the udder protects against experimental Escherichia coli mastitis. Innate Immun. 2012, 18, 467–477. [Google Scholar] [CrossRef]
- Schukken, Y.H.; Günther, J.; Fitzpatrick, J.; Fontaine, M.C.; Goetze, L.; Holste, O.; Leigh, J.; Petzlg, W.; Schuberth, H.J.; Sipkah, A.; et al. Host-response patterns of intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 2011, 144, 270–289. [Google Scholar] [CrossRef]
- Wysocka, M.; Robertson, S.; Riemann, H.; Caamano, J.; Hunter, C.; Mackiewicz, A.; Montaner, L.J.; Trinchieri, G.; Karp, C.L. IL-12 suppression during experimental endotoxin tolerance: Dendritic cell loss and macrophage hyporesponsiveness. J. Immunol. 2001, 166, 7504–7513. [Google Scholar] [CrossRef]
- Günther, J.; Petzl, W.; Zerbe, H.; Schuberth, H.J.; Koczan, D.; Goetze, L.; Seyfert, H.M. Lipopolysaccharide priming enhances expression of effectors of immune defence while decreasing expression of pro-inflammatory cytokines in mammaryepithelial cells from cows. BMC Genom. 2012, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Breyne, K.; Steenbrugge, J.; Demeyere, K.; Vanden Berghe, T.; Meyer, E. Preconditioning with lipopolysaccharide or lipoteichoic acid protects against Staphylococcus aureus mammary infection in mice. Front. Immunol. 2017, 8, 833. [Google Scholar] [CrossRef] [PubMed]
- Petzl, W.; Zerbea, H.; Günther, J.; Seyfert, H.M.; Hussen, J.; Schubert, H.J. Pathogen-specific responses in the bovine udder. Models and immunoprophylactic concepts. Res. Vet. Sci. 2018, 116, 55–61. [Google Scholar] [CrossRef]
- Günther, J.; Petzl, W.; Zerbe, H.; Schuberth, H.J.; Seyfert, H.M. TLR ligands, but not modulators of histone modifiers, can induce the complex immune response pattern of endotoxin tolerance in mammary epithelial cells. Innate Immun. 2017, 23, 155–164. [Google Scholar] [CrossRef]
- Gelasakis, A.I.; Mavrogianni, V.S.; Petridis, I.G.; Vasileiou, N.G.C.; Fthenakis, G.C. Mastitis in sheep-The last 10 years and the future of research. Vet. Microbiol. 2015, 181, 136–146. [Google Scholar] [CrossRef]
- Hughes, K.; Watson, C.J. The mammary microenvironment in mastitis in humans, dairy ruminants, rabbits and rodents: A One Health focus. J. Mammary Gland Biol. Neopl. 2018, 23, 27–41. [Google Scholar] [CrossRef]
- Moroni, P.; Cuccuru, C. Relationship between mammary gland infections and some milk immune parameters in Sardenian ewes. Small Rumin. Res. 2001, 41, 1–7. [Google Scholar] [CrossRef]
- Bergonier, D.; Berthelot, X. New advances in epizootiology and control of ewe mastitis. Liv. Prod. Sci. 2003, 79, 1–16. [Google Scholar] [CrossRef]
- Berthelot, X.; Lagriffoul, G.; Concordet, D.; Barillet, F.; Bergonier, D. Physiological and pathological thresholds of somatic cell counts in ewe milk. Small Rumin. Res. 2006, 62, 27–31. [Google Scholar] [CrossRef]
- Albenzio, M.; Santillo, A.; Caroprese, M.; Ruggieri, D.; Ciliberti, M.; Sevi, A. Immune competence of the mammary gland as affected by somatic cell and pathogenic bacteria in ewes with subclinical mastitis. J. Dairy Sci. 2012, 95, 3877–3887. [Google Scholar] [CrossRef] [PubMed]
- Souza, F.N.; Blagitz, M.G.; Penna, F.A.M.; Della Libera, A.M.M.P.; Heinemann, M.B.; Cerqueira, M.M.O.P. Somatic cell count in small ruminants: Friend or foe? Small Rumin. Res. 2012, 107, 65–75. [Google Scholar] [CrossRef]
- Cuccuru, C.; Moroni, P.; Zecconi, A.; Casu, S.; Caria, A.; Contini, A. Milk differential cell counts in relation to total counts in Sardinian ewes. Small Rumin. Res. 1997, 25, 169–173. [Google Scholar] [CrossRef]
- Boutinaud, M.; Jammes, H. Potential uses of milk epithelial cells: A review. Reprod. Nutr. Dev. 2002, 42, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Paape, M.J.; Wiggans, G.R.; Bannerman, D.D.; Thomas, D.L.; Sanders, A.H.; Contreras, A.; Moroni, P.; Miller, R.H. Monitoring goat and sheep milk somatic cell counts. J. Dairy Sci. 2007, 68, 114–125. [Google Scholar] [CrossRef]
- Albenzio, M.; Figliola, L.; Caroprese, M.; Marino, R.; Sevi, A.; Santillo, A. Somatic cell count in sheep milk. Small Rumin. Res. 2019, 176, 24–30. [Google Scholar] [CrossRef]
- Paape, M.J.; Poutrel, B.; Contreras, A.; Marco, J.C.; Capuco, A.V. Milk somatic cells and lactation in small ruminants. J. Dairy Sci. 2001, 84, E237–E244. [Google Scholar] [CrossRef]
- Leitner, G.; Merin, U.; Krifucks, O.; Blum, S.; Rivas, A.; Silanikove, N. Effects of intramammary bacterial infection with coagulase negative staphylococci and stage of lactation on shedding of epithelial cells and infiltration of leukocytes into milk: Comparison among cows, goats and sheep. Vet. Immunol. Immunopathol. 2012, 147, 202–210. [Google Scholar] [CrossRef] [PubMed]
- El-Masannat, E.T.S. A Study of Ovine Mastitis with Special Reference to Mastitis caused by Pasteurella haemolytica. Ph.D. Thesis, University of London, London, UK, 1987. [Google Scholar]
- Fthenakis, G. Ovine Mastitis with Special Reference to Subclinical Mastitis associated with Coagulase-Negative Staphylococci. Ph.D. Thesis, University of London, London, UK, 1988. [Google Scholar]
- Cheville, N.F. Ultrastructural Pathology: The Comparative Cellular Basis of Disease, 2nd ed.; Wiley-Blackwell: Ames, IA, USA, 2009; p. 1000. [Google Scholar]
- Persson, W.; Bengtsson, K.B.; Lindberg, A.; Nyman, A.; Unnerstad, H.E. Incidence of mastitis and bacterial findings at clinical mastitis in Swedish primiparous cows. Influence of breed and stage of lactation. Vet. Microbiol. 2009, 134, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Persson Waller, K.; Colditz, I.G.; Flapper, P.; Seow, H.F. Leukocyte and cytokine accumulation in the ovine teat and udder during endotoxin-induced inflammation. Vet. Res. Comm. 1997, 21, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Barbagianni, M.S. Experimental Study of Pregnancy Toxaemia in Ewes and its Association with Mastitis in the; Post-Partum Period. Ph.D. Thesis, University of Thessaly, Karditsa, Greece, 2017. [Google Scholar]
- Barbagianni, M.S.; Mavrogianni, V.S.; Vasileiou, N.G.C.; Fthenakis, G.C.; Petridis, I.G. Ultrasonographic examination of the udder in sheep. Small Rumin. Res. 2017, 152, 86–99. [Google Scholar] [CrossRef]
- Hodgson, J.C.; Moon, G.M.; Quirie, M.; Donachie, W. Association of LPS chemotype of Mannheimia (Pasteurella) haemolytica A1 with disease virulence in a model of ovine pneumonic pasteurellosis. J. Endotoxin Res. 2003, 9, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Addis, M.F.; Pisanu, S.; Marogna, G.; Cubeddu, T.; Pagnozzi, D.; Cacciotto, C.; Campesi, F.; Schianchi, G.; Rocca, S.; Uzzau, S. Defense proteins by mammary epithelial cells following Streptococcus uberis infection in sheep. Inf. Immun. 2013, 81, 3182–3197. [Google Scholar] [CrossRef] [PubMed]
- Winter, P.; Colditz, I.G. Immunological responses of the lactating ovine udder following experimental challenge with Staphylococcus epidermidis. Vet. Immunol. Immunopathol. 2002, 89, 57–65. [Google Scholar] [CrossRef]
- Vray, B. Macrophages in parasitic infection. In The Macrophage; Burke, B., Lewis, C.E., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 253–304. [Google Scholar]
- De Garnica, M.L.; Linage, B.; Carriedo, J.A.; De La Fuente, L.F.; García-Jimeno, M.C.; Santos, J.A.; Gonzalo, C. Relationship among specific bacterial counts and total bacterial and somatic cell counts and factors influencing their variation in ovine bulk tank milk. J. Dairy Res. 2013, 96, 1021–1029. [Google Scholar] [CrossRef]
- Fthenakis, G.C. California Mastitis Test and Whiteside Test in diagnosis of subclinical mastitis of dairy ewes. Small Rumin. Res. 1995, 16, 271–276. [Google Scholar] [CrossRef]
- Fragkou, I.A.; Boscos, C.M.; Fthenakis, G.C. Diagnosis of clinical or subclinical mastitis in ewes. Small Rumin. Res. 2014, 118, 86–92. [Google Scholar] [CrossRef]
- Van Oostveld, K.; Paape, M.J.; Dosogne, H.; Burvenich, C. Effect of apoptosis on phagocytosis, respiratory burst and CD18 adhesion receptor expression of bovine neutrophil. Dom. Anim. Endocrinol. 2002, 22, 37–50. [Google Scholar] [CrossRef]
- Van Harten, R.M.; van Woudenbergh, E.; van Dijk, A.; Haagsman, H.P. Cathelicidins: Immunomodulatory antimicrobials. Vaccines 2018, 6, 63. [Google Scholar] [CrossRef] [PubMed]
- Sladek, Z.; Rysanek, D. Neutrophil apoptosis during the resolution of bovine mammary gland injury. Res. Vet. Sci. 2001, 70, 41–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldsby, R.A. Cells and organs of the immune system. In Kuby Immunology, 6th ed.; Osborne, B.A., Goldsby, R.A., Kindt, T.J., Eds.; Freeman: New York, NY, USA, 2006; pp. 40–47. [Google Scholar]
- Bonelli, P.; Dimauro, C.; Re, R.; Pilo, G.; Dore, S.; Cannas, A.E.; Nicolussi, P.S. Peripheral blood and milk leukocytes subsets of lactating Sarda ewes. Ital. J. Anim. Sci. 2013, 12, e34. [Google Scholar] [CrossRef]
- Swiderek, W.P.; Charon, K.M.; Winnicka, A.; Gruszczynska, J. Relationship between blood lymphocyte phenotype, DRB1 MHC class II) gene polymorphism and somatic cell count in ewe milk. Bull. Vet. Inst. Pulaway 2006, 50, 73–77. [Google Scholar]
- Rainard, P.; Foucras, G.; Boichard, D.; Rupp, R. Low milk somatic cell count and susceptibility to mastitis. J. Dairy Sci. 2018, 101, 6703–6714. [Google Scholar] [CrossRef] [PubMed]
- Craven, N.; Williams, M.R. Defences of the bovine mammary gland against infection and prospects for their enhancement. Vet. Immunol. Immunopathol. 1985, 10, 71–127. [Google Scholar] [CrossRef]
- Norcross, N.L. Specific defence mechanisms of the udder. Flem. Vet. J. 1991, 62, 129–139. [Google Scholar]
- Oestensson, K.; Lun, S. Transfer of immunoglobulins through the mammary endothelium and epithelium and in the local lymph node of cows during the initial response after intramammary challenge with E. coli endotoxin. Acta Vet. Scand. 2008, 50, 26. [Google Scholar] [CrossRef]
- Wellnitz, O.; Zbinden, C.; Lüttgenau, J.; Bollwein, H.; Bruckmaier, R.M. Different chronological patterns of appearance of blood derived milk components during mastitis indicate different mechanisms of transfer from blood into milk. J. Dairy Res. 2015, 82, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Hine, B.C.; Hunt, P.W.; Beasley, A.M.; Windon, R.G.; Glover, S.A.; Colditz, I.G. Selective transport of IgE into ovine mammary secretions. Res. Vet. Sci. 2010, 89, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Barrio, M.B.; Rainard, P.; Poutrel, B. Milk complement and the opsonophagocytosis and killing of Staphylococcus aureus mastitis isolates by bovine neutrophils. Microb. Pathogen. 2003, 34, 1–9. [Google Scholar] [CrossRef]
- Marnila, P.; Korhonen, H. Immunoglobulins. In Encyclopedia of Dairy Sciences; Roginski, H., Fuquay, J.W., Fox, P.F., Eds.; Academic Press: London, UK, 2002; pp. 1950–1956. [Google Scholar]
- Mehra, R.; Marnila, P.; Korhonen, H. Milk immunoglobulins for health promotion. Int. Dairy J. 2006, 16, 1262–1271. [Google Scholar] [CrossRef]
- Hernández-Castellano, L.E.; Almeida, A.; Renaut, J.; Argüello, A.; Castro, N. A proteomics study of colostrum and milk from the two-major small ruminant dairy breeds from the Canary Islands: A bovine milk comparison perspective. J. Dairy Res. 2016, 83, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Rainard, P. The complement in milk and defense of the bovine mammary gland against infections. Vet. Res. 2003, 34, 647–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arbore, G.; Kemper, C.; Kolev, M. Intracellular complement-the complosome-in immune cell regulation. Mol. Immunol. 2017, 89, 2–9. [Google Scholar] [CrossRef] [PubMed]
- DiCarlo, A.; Paape, M. Comparison of C3b binding to bovine peripheral blood and mammary gland neutrophils (PMN). Am. J. Vet. Res. 1996, 57, 151–156. [Google Scholar]
- Cunnion, K.M.; Lee, J.C.; Frank, M.M. Capsule production and growth phase influence binding of complement to Staphylococcus aureus. Infect. Immun. 2001, 69, 6796–6803. [Google Scholar] [CrossRef] [PubMed]
- Katsafadou, A.I.; Tsangaris, G.T.; Billinis, C.; Fthenakis, G.C. Use of proteomics in the study of microbial diseases of small ruminants. Vet. Microbiol. 2015, 181, 27–33. [Google Scholar] [CrossRef]
- Pereyra, E.A.L.; Sacco, S.C.; Dure, A.; Baravalle, C.; Renna, M.S.; Andreotti, C.S.; Carolina, S.; Monecke, S.; Calvinho, L.F.; Dallard, B.E. Immune response of Staphylococcus aureus strains in a mouse mastitis model is linked to adaptive capacity and genotypic profile. Vet. Microbiol. 2017, 204, 64–76. [Google Scholar] [CrossRef]
- Pierce, A.; Legrand, D.; Mazurier, J. Lactoferrin: A multifunctional protein. Med. Sci. (Paris) 2009, 25, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, K.; Kawai, K. Advances in lactoferrin research concerning bovine mastitis. Bioch. Cell Biol. 2017, 95, 69–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isobe, N. Control mechanisms for producing antimicrobial factors in ruminant mammary gland. Anim. Sci. J. 2017, 88, 937–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Rowney, M.; Guo, P.; Hobman, P. Biological properties of lactoferrin: An overview. Aust. J. Dairy Technol. 2007, 62, 31–42. [Google Scholar]
- Lemos, V.F.; Guaraná, E.L.S.; Afonso, J.A.B.; Fagliari, J.J.; Silva, P.S.; Soares, P.C.; de Mendonça, C.L. Effects of intramammary infection on whey proteinograms of sheep during lactation. Pesq. Bras. 2015, 35, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Alekish, M.; Ababneh, H.; Ismail, Z.; Alshehabat, M. The relationship between lactoferrin gene polymorphism and subclinical mastitis in Awassi ewes. J. Anim. Plant. Sci. 2019, 29, 1193–1197. [Google Scholar]
- Liu, H.; Zheng, F.; Cao, Q.; Ren, B.; Zhu, L.; Striker, G.; Vlassara, H. Amelioration of oxidant stress by the defensin lysozyme. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E824–E832. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kovacs-Nolan, J.; Chengbo, Y.; Archbold, T.; Fan, M.Z.; Mine, Y. Hen egg lysozyme attenuates inflammation and modulates local gene expression in a porcine model of dextran sodium sulfate (DSS)-induced colitis. J. Agric. Food Chem. 2009, 57, 62233–62240. [Google Scholar] [CrossRef]
- Fox, P.F.; Kelly, A.L. Indigenous enzymes in milk: Overview and historical aspects-part 1. Int. Dairy J. 2006, 16, 500–516. [Google Scholar] [CrossRef]
- Noble, W.C.; Somerville, D.A. Skin as a habitat. In Microbiology of Human Skin; Noble, W.C., Ed.; Saunders: London, UK, 1974; pp. 3–78. [Google Scholar]
- Sieber, R.L.; Farnsworth, R.J. Differential diagnosis of bovine teat lesions. Vet. Clin. N. Am. Large Anim. Pract. 1984, 6, 313–321. [Google Scholar] [CrossRef]
- Fox, L.K.; Hancock, D.D. Effect of segregation on prevention of intramammary infections by Staphylococcus aureus. J. Dairy Sci. 1989, 72, 540–544. [Google Scholar] [CrossRef]
- Fox, L.K.; Nagy, J.A.; Hillers, J.K.; Cronrath, J.D.; Ratkowsky, D.A. Effects of post-milking teat treatment on the colonization of Staphylococcus aureus on chapped teat skin. Am. J. Vet. Res. 1991, 52, 799–802. [Google Scholar] [PubMed]
- Mavrogianni, V.S.; Cripps, P.J.; Papaioannou, N.; Taitzoglou, I.; Fthenakis, G.C. Teat disorders predispose ewes to clinical mastitis after challenge with Mannheimia haemolytica. Vet. Res. 2006, 37, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Fragkou, I.A.; Papaioannou, N.; Cripps, P.J.; Boscos, C.M.; Fthenakis, G.C. Teat lesions predispose to invasion of the ovine mammary gland by Mannheimia haemolytica. J. Comp. Pathol. 2007, 137, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Fragkou, I.A.; Skoufos, J.; Cripps, P.J.; Kyriazakis, I.; Papaioannou, N.; Boscos, C.M.; Tzora, A.; Fthenakis, G.C. Differences in susceptibility to M. haemolytica-associated mastitis between two breeds of dairy sheep. J. Dairy Res. 2007, 74, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Baro, J.A.; Carriedo, J.A.; San Primitivo, F. Genetic paramerers of test day measure for somatic cell count, milk yield and protein percentage of milking ewes. J. Dairy Sci. 1994, 77, 2568–2662. [Google Scholar] [CrossRef]
- Barillet, F.; Rupp, R.; Mignon-Grasteau, S.; Astruc, J.M.; Jacquin, M. Genetic analysis for mastitis resistance and milk somatic cell score in French Lacaune dairy sheep. Gen. Sel. Evol. 2001, 33, 397–415. [Google Scholar] [CrossRef] [Green Version]
- Bishop, S.C. Genetic resistance to infections in sheep. Vet. Microbiol. 2015, 181, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Rupp, R.; Senin, P.; Sarry, J.; Allain, C.; Tasca, C.; Ligat, L.; Portes, D.; Woloszyn, F.; Bouchez, O.; Tabouret, G.; et al. A point mutation in suppressor of cytokine signalling 2 (Socs2) increases the susceptibility to inflammation of the mammary gland while associated with higher body weight and size and higher milk production in a sheep model. PLoS Genet. 2015, 11, e1005629. [Google Scholar] [CrossRef]
- Li, T.T.; Gao, J.F.; Zhao, X.X.; Ma, Y.J. Digital gene expression analyses of mammary glands from meat ewes naturally infected with clinical mastitis. R. Soc. Open Sci. 2019, 6, 181604. [Google Scholar] [CrossRef] [Green Version]
- Barbagianni, M.S.; Mavrogianni, V.S.; Katsafadou, A.I.; Spanos, S.A.; Tsioli, V.; Galatos, A.D.; Nakou, M.; Valasi, I.; Gouletsou, P.G.; Fthenakis, G.C. Pregnancy toxaemia as predisposing factor for development of mastitis in sheep during the immediately post-partum period. Small Rumin. Res. 2015, 130, 246–251. [Google Scholar] [CrossRef]
- Giadinis, N.D.; Panousis, N.; Petridou, E.J.; Siarkou, V.I.; Lafi, S.Q.; Pourliotis, K.; Hatzopoulou, E.; Fthenakis, G.C. Selenium, vitamin E and vitamin A blood concentrations in dairy sheep flocks with increased or low clinical mastitis incidence. Small Rumin. Res. 2011, 95, 193–196. [Google Scholar] [CrossRef]
- Koutsoumpas, A.T.; Giadinis, N.D.; Petridou, E.J.; Konstantinou, E.; Brozos, C.; Lafi, S.Q.; Fthenakis, G.C.; Karatzias, H. Consequences of reduced vitamin A administration on mammary health of dairy ewes. Small Rumin. Res. 2013, 110, 120–123. [Google Scholar] [CrossRef]
- Azara, E.; Longheu, C.; Sanna, G.; Tola, S. Biofilm formation and virulence factor analysis of Staphylococcus aureus isolates collected from ovine mastitis. J. Appl. Microbiol. 2017, 123, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.M.; Prenafeta, A.; Valle, J.; Penades, J.; Rota, C.; Solano, C.; Marco, J.; Grillo, M.J.; Lasa, I.; Irache, J.M.; et al. Protection from Staphylococcus aureus mastitis associated with poly-N-acetyl beta-1,6 glucosamine specific antibody production using biofilm-embedded bacteria. Vaccine 2009, 17, 2379–2386. [Google Scholar] [CrossRef] [PubMed]
- Vasileiou, N.G.C.; Chatzopoulos, D.C.; Cripps, P.J.; Ioannidi, K.S.; Gougoulis, D.A.; Chouzouris, T.M.; Lianou, D.T.; Calvo Gonzalez-Valerio, T.; Guix Vallverdu, R.; Argyros, S.; et al. Evaluation of efficacy of a biofilm-embedded bacteria-based vaccine against staphylococcal mastitis in sheep-A randomized, placebo-controlled field study. J. Dairy Sci. 2019, 102, 9328–9344. [Google Scholar] [CrossRef] [PubMed]
- Ballingall, K.T.; Todd, H.; Tassi, R. Intra-mammary vaccine delivery protects sheep against mastitis caused by mannheimia haemolytica infection. In Proceedings of the Abstract Book of International Veterinary Immunology Symposium, Seattle, WA, USA, 13–16 August 2019; pp. 71–72. [Google Scholar]



{kind=link}
{kind=link}
| Leucocyte Types | Proportion of Total Leucocytes [59,60,61] |
|---|---|
| Macrophages | 40–85% |
| Neutrophils | 5–35% |
| Lymphocytes | 10–20% |
| Cytokine | Main Role |
|---|---|
| Interleukin-1 | Upregulation of neutrophil migration to the mammary gland, increase of neutrophil numbers, promotion of phagocytic activity in the mammary gland. |
| Interleukin-2 | Upregulation of macrophage proliferation in the mammary gland, improvement of lymphocyte antibacterial properties. |
| Interleukin-8 | Upregulation of neutrophil migration to the mammary gland. |
| Granulocyte colony-stimulating factor | Increase of neutrophil numbers, promotion of phagocytic activity in the mammary gland. |
| Granulocyte macrophage colony-stimulating factor | Improvement of neutrophil chemotaxis and bactericidal properties, increase of neutrophil numbers in the mammary gland. |
| Macrophage colony-stimulating factor | Upregulation of macrophage proliferation. |
| Interferon-γ | Promotion of neutrophil phagocytic activity. |
| Tumour necrosis factor | Enhancement of inflammatory process, promotion of neutrophil phagocytic activity. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katsafadou, A.I.; Politis, A.P.; Mavrogianni, V.S.; Barbagianni, M.S.; Vasileiou, N.G.C.; Fthenakis, G.C.; Fragkou, I.A. Mammary Defences and Immunity against Mastitis in Sheep. Animals 2019, 9, 726. https://doi.org/10.3390/ani9100726
Katsafadou AI, Politis AP, Mavrogianni VS, Barbagianni MS, Vasileiou NGC, Fthenakis GC, Fragkou IA. Mammary Defences and Immunity against Mastitis in Sheep. Animals. 2019; 9(10):726. https://doi.org/10.3390/ani9100726
Chicago/Turabian StyleKatsafadou, Angeliki I., Antonis P. Politis, Vasia S. Mavrogianni, Mariana S. Barbagianni, Natalia G. C. Vasileiou, George C. Fthenakis, and Ilektra A. Fragkou. 2019. "Mammary Defences and Immunity against Mastitis in Sheep" Animals 9, no. 10: 726. https://doi.org/10.3390/ani9100726
APA StyleKatsafadou, A. I., Politis, A. P., Mavrogianni, V. S., Barbagianni, M. S., Vasileiou, N. G. C., Fthenakis, G. C., & Fragkou, I. A. (2019). Mammary Defences and Immunity against Mastitis in Sheep. Animals, 9(10), 726. https://doi.org/10.3390/ani9100726