Grape Seed Procyanidin Extract (GSPE) Improves Goat Sperm Quality When Preserved at 4 °C

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Reagents
2.2. Goat Semen Collection
2.3. Goat Semen Preservation
2.4. Sperm Motility
2.5. Sperm Functional Plasma Membrane Integrity
2.6. Acrosomal Membrane Integrity
2.7. Mitochondrial Activity
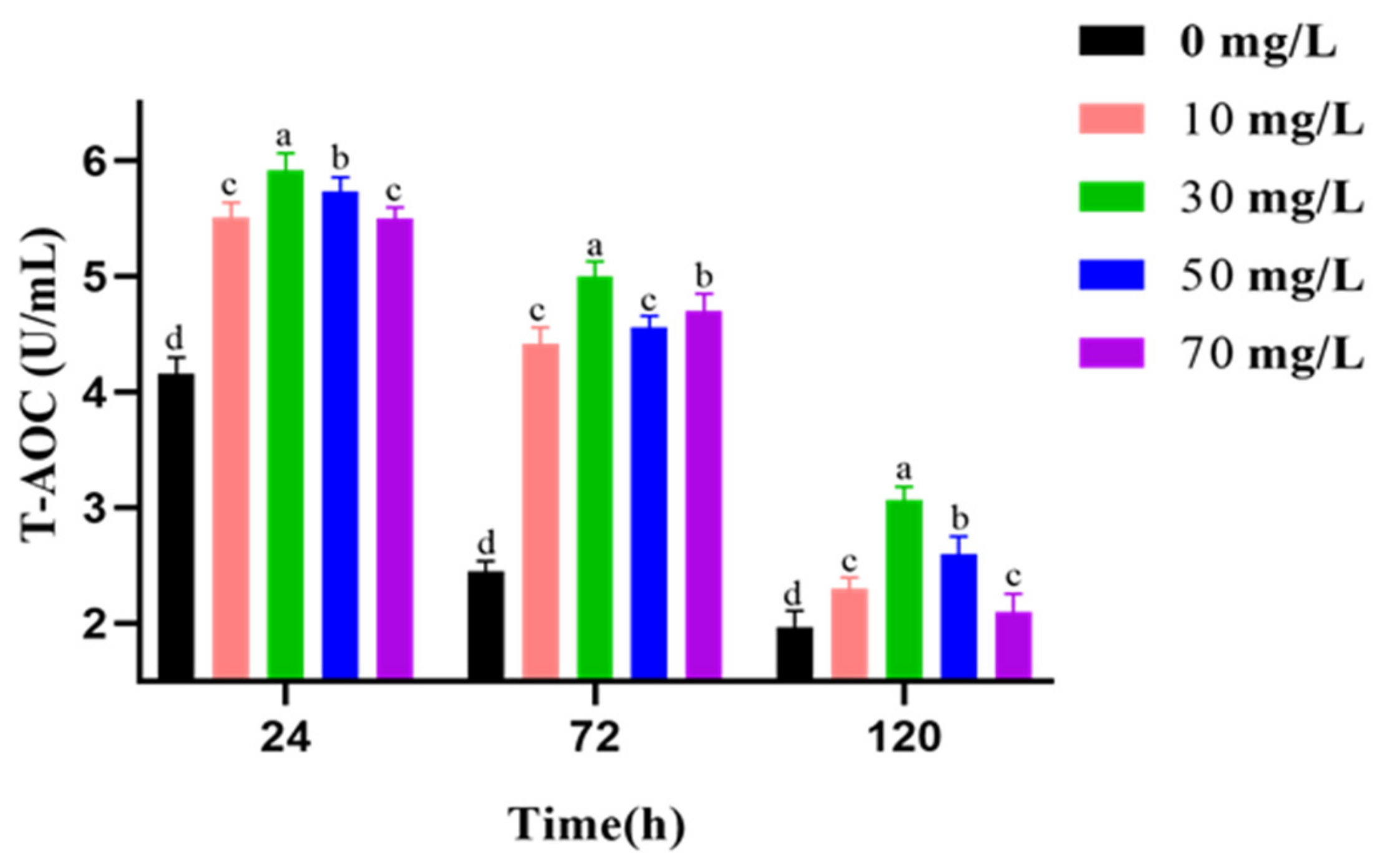
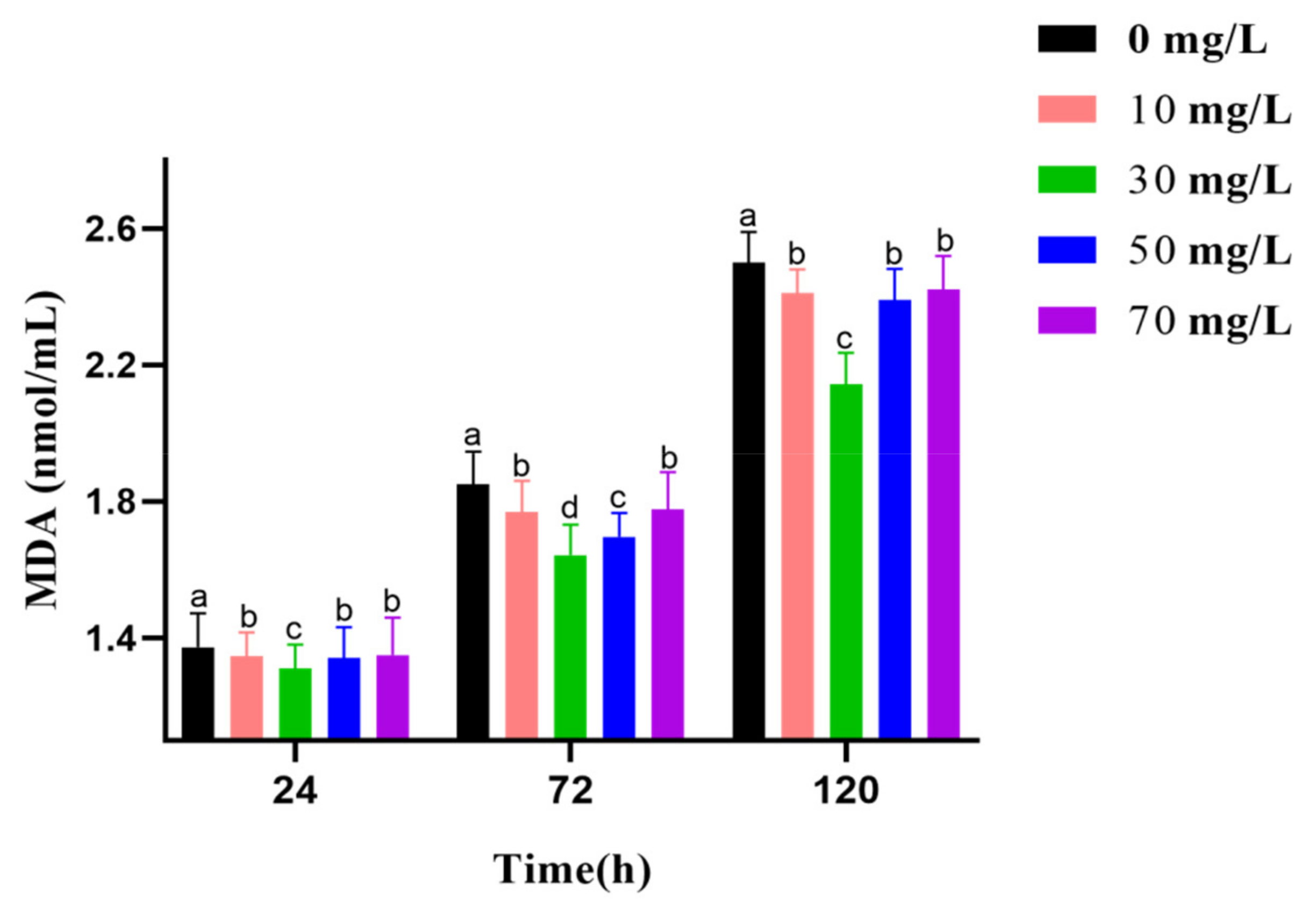
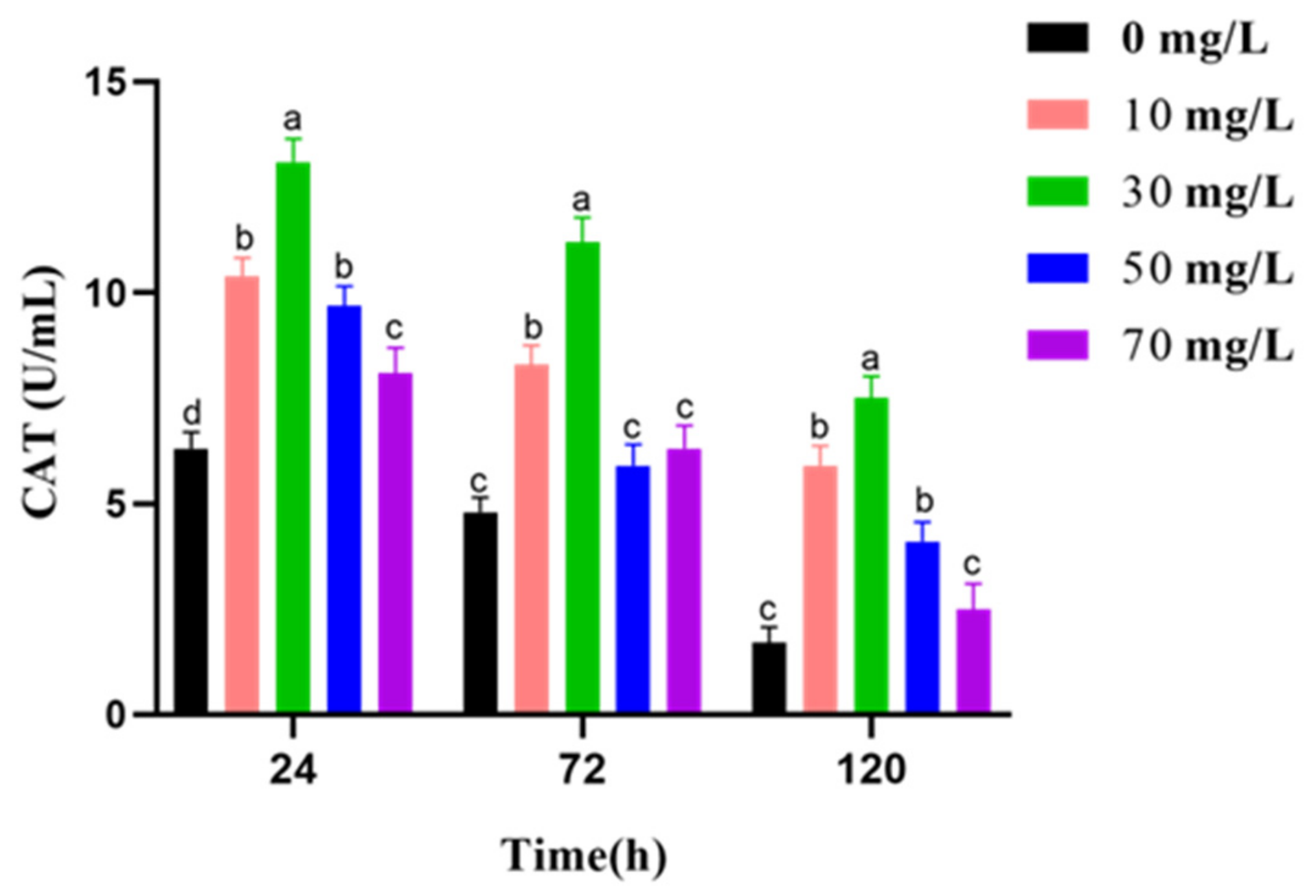
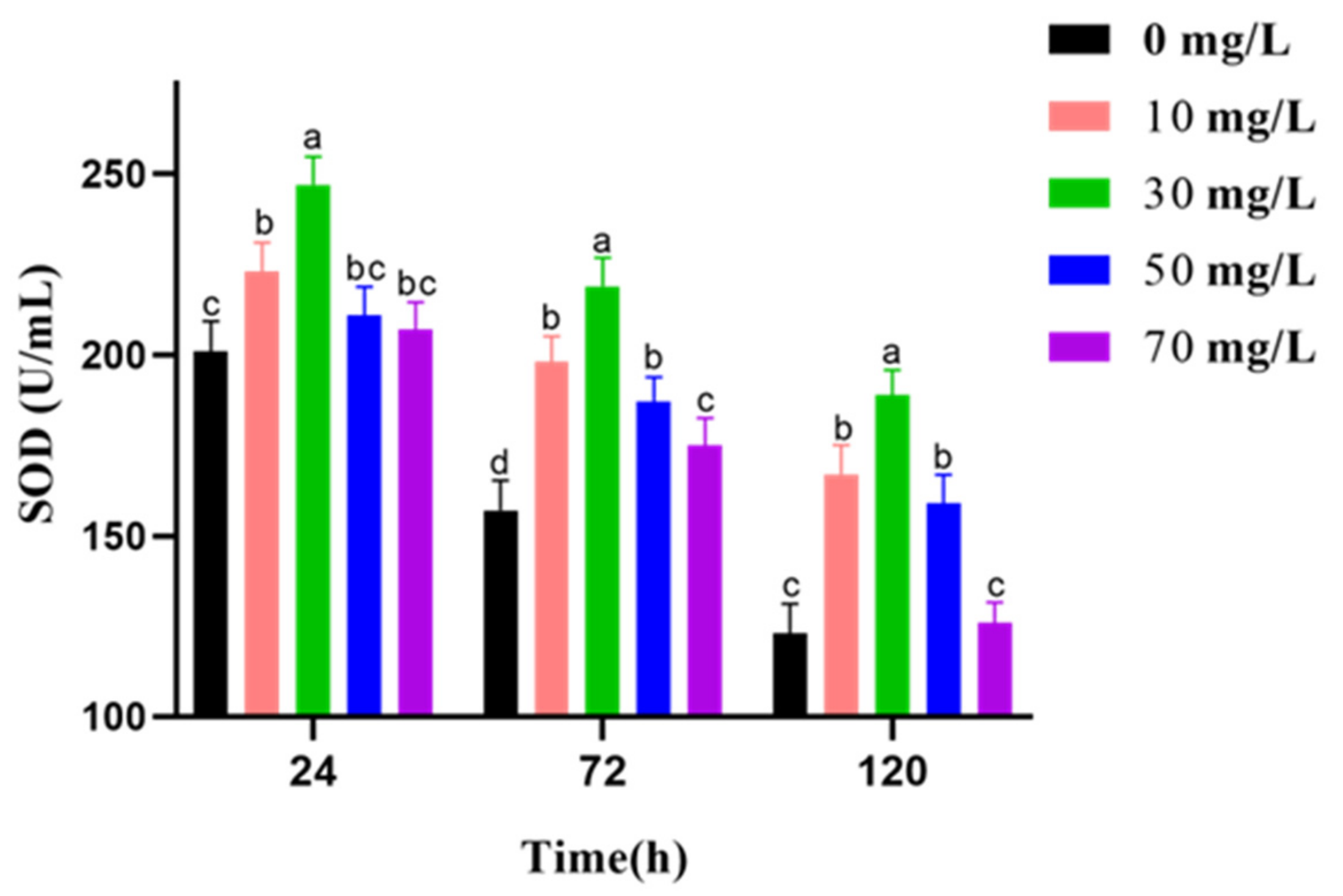
2.8. Measurement of T-AOC, SOD Activities, CAT Activities, and MDA Level
2.9. Fertility Trials
2.10. Statistical Analysis
3. Results
3.1. Sperm Motility
3.2. Functional Plasma Membrane Integrity
3.3. Acrosome Integrity
3.4. Mitochondrial Activity
3.5. Analysis of Antioxidant Activities
3.6. Pregnancy Rates and Litter Sizes
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ren, F.; Fang, Q.; Feng, T.Y.; Li, Y.; Wang, Y.H.; Zhu, H.J.; Hu, J.H. Lycium barbarum and Laminaria japonica polysaccharides improve Cashmere goat sperm quality and fertility rate after cryopreservation. Theriogenology 2019, 129, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Casali, R.; Pinczak, A.; Cuadro, F.; Guillen-Munoz, J.M.; Mezzalira, A.; Menchaca, A. Semen deposition by cervical, transcervical and intrauterine route for fixed-time artificial insemination (FTAI) in the ewe. Theriogenology 2017, 103, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.T.; Han, D.; Xu, C.L.; Luo, M.J.; Chang, Z.L.; Tan, J.H. Protocol optimization for long-term liquid storage of goat semen in a chemically defined extender. Reprod. Domest. Anim. 2009, 44, 865–872. [Google Scholar] [CrossRef] [PubMed]
- O’Hara, L.; Hanrahan, J.P.; Richardson, L.; Donovan, A.; Fair, S.; Evans, A.C.O.; Lonergan, P. Effect of storage duration, storage temperature, and diluent on the viability and fertility of fresh ram sperm. Theriogenology 2010, 73, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Paul, R.K.; Balaganur, K.; Bahire, S.V.; Kumar, D.; Singh, R. Supplementation of cauda epididymal plasma improves sperm characteristics following liquid preservation of ram semen at 3–5 °C. Reprod. Fertil. Dev. 2018, 30, 1389–1401. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, W.M.; Salamon, S. Liquid storage of ram semen: A review. Reprod. Fertil. Dev. 1993, 5, 613–638. [Google Scholar] [CrossRef]
- Fang, Q.; Wang, J.; Hao, Y.Y.; Li, H.; Hu, J.X.; Yang, G.S. Effects of iodine methionine on boar sperm quality during liquid storage at 17 °C. Reprod. Domest. Anim. 2017, 52, 1061–1066. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.G.; Fang, Q.; Liu, Q.; Du, R.R.; Yang, G.S. Supplemental effect of different levels of taurine in modena on boar semen quality during liquid preservation at 17 °C. Anim. Sci. J. 2017, 88, 1692–1699. [Google Scholar] [CrossRef]
- Barrios, B.; Pérez-Pé, R.; Gallego, M.; Tato, A.; Osada, J.; Muino-Blanco, T.; Cebrian-Perez, J.A. Seminal plasma proteins revert the cold-shock damage on ram sperm membrane. Biol. Reprod. 2000, 63, 1531–1537. [Google Scholar] [CrossRef]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Bilodeau, J.F.; Blanchette, S.; Gagnon, C.; Sirard, M.A. Thiols prevent h2o2-mediated loss of sperm motility in cryopreserved bull semen. Theriogenology 2001, 56, 275–286. [Google Scholar] [CrossRef]
- Domosławska, A.; Zdunczyk, S.; Franczyk, M.; Kankofer, M.; Janowski, T. Selenium and vitamin e supplementation enhances the antioxidant status of spermatozoa and improves semen quality in male dogs with lowered fertility. Andrologia 2018, 50, e13023. [Google Scholar] [CrossRef] [PubMed]
- Llópiz, N.; Puiggròs, F.; Céspedes, E.; Arola, L.; Ardévol, A.; Bladé, C.; Salvado, M.J. Antigenotoxic effect of grape seed procyanidin extract in fao cells submitted to oxidative stress. J. Agric. Food Chem. 2004, 52, 1083–1087. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Garg, A.; Krohn, R.L.; Bagchi, M.; Tran, M.X.; Stohs, S.J. Oxygen free radical scavenging abilities of vitamins c and e, and a grape seed proanthocyanidin extract in vitro. Res. Commun. Mol. Pathol. Pharmacol. 1997, 95, 179–189. [Google Scholar]
- Décordé, K.; Teissèdre, P.L.; Sutra, T.; Ventura, E.; Cristol, J.P.; Rouanet, J.M. Chardonnay grape seed procyanidin extract supplementation prevents high-fat diet-induced obesity in hamsters by improving adipokine imbalance and oxidative stress markers. Mol. Nutr. Food Res. 2009, 53, 659–666. [Google Scholar] [CrossRef]
- Guo, L.; Wang, L.H.; Sun, B.; Yang, J.Y.; Zhao, Y.Q.; Dong, Y.X. Direct in vivo evidence of protective effects of grape seed procyanidin fractions and other antioxidants against ethanol-induced oxidative dna damage in mouse brain cells. J. Agric. Food Chem. 2007, 55, 5881–5891. [Google Scholar] [CrossRef]
- Su, L.; Deng, Y.; Zhang, Y.; Li, C.; Zhang, R.; Sun, Y. Protective effects of grape seed procyanidin extract against nickel sulfate-induced apoptosis and oxidative stress in rat testes. Toxicol. Mech. Methods 2011, 21, 487–494. [Google Scholar] [CrossRef]
- Mata-Campuzano, M.; Soleilhavoup, C.; Tsikis, G.; Martinez-Pastor, F.; de Graaf, S.P.; Druart, X. Motility of liquid stored ram spermatozoa is altered by dilution rate independent of seminal plasma concentration. Anim. Reprod. Sci. 2015, 162, 31–36. [Google Scholar] [CrossRef]
- Lodhi, L.A.; Zubair, M.; Qureshi, Z.I.; Ahmad, I.; Jamil, H. Correlation between hypo-osmotic swelling test and various conventional semen evaluation parameters in fresh Nili-Ravi buffalo and Sahiwal cow bull semen. Pak. Vet. J. 2008, 28, 186–188. [Google Scholar]
- Olivera-Muzante, J.; Fierro, S.; Gil, J. Conception rates in ewes after ai with ram semen preserved in milk-egg yolk extenders supplemented with glycerol. Reprod. Domest. Anim. 2011, 46, 508–512. [Google Scholar] [CrossRef]
- Rodrigues, M.A.M.; Souza, C.E.A.; Martins, J.A.M.; Rego, J.P.A.; Oliveira, J.T.A.; Domont, G. Seminal plasma proteins and their relationship with sperm motility in santa ines rams. Small Rumin. Res. 2013, 109, 94–100. [Google Scholar] [CrossRef]
- Güvenç, M.; Cellat, M.; Gökçek, İ.; Yavaş, İ.; Yurdagül Özsoy, Ş. Effects of thymol and carvacrol on sperm quality and oxidant/antioxidant balance in rats. Arch. Physiol. Biochem. 2019, 125, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, F.; Liang, M.; Chen, S.; Zhu, Y.; Zou, Z. Grape seed proanthocyanidin extract attenuates varicocele-induced testicular oxidative injury in rats by activating the nrf2-antioxidant system. Mol. Med. Rep. 2018, 17, 1799–1806. [Google Scholar] [PubMed]
- Fernandes, G.H.; De Carvalho, P.T.; Serra, A.J.; Crespilho, A.M.; Peron, J.P.; Rossato, C. The effect of low-level laser irradiation on sperm motility, and integrity of the plasma membrane and acrosome in cryopreserved bovine sperm. PLoS ONE 2015, 10, e0121487. [Google Scholar] [CrossRef]
- Sutovsky, P.; Navara, C.S.; Schatten, G. Fate of the sperm mitochondria, and the incorporation, conversion, and disassembly of the sperm tail structures during bovine fertilization. Biol. Reprod. 1996, 55, 1195–1205. [Google Scholar] [CrossRef]
- Ruiz-Pesini, E.; Díez-Sánchez, C.; López-Pérez, M.J.; Enriquez, J.A. The role of the mitochondrion in sperm function: Is there a place for oxidative phosphorylation or is this a purely glycolytic process? Curr. Top. Dev. Biol. 2007, 77, 3–19. [Google Scholar]
- Zhao, Y.M.; Gao, L.P.; Zhang, H.L.; Guo, J.X.; Guo, P.P. Grape seed proanthocyanidin extract prevents ddp-induced testicular toxicity in rats. Food Funct. 2014, 5, 605–611. [Google Scholar] [CrossRef]
- Foo, L.Y. Proanthocyanidins: Gross chemical structures by infrared spectra. Phytochemistry 1981, 20, 1397–1402. [Google Scholar] [CrossRef]
- Qiu, J.H.; Li, Y.W.; Xie, H.L.; Li, Q.; Dong, H.B.; Sun, M.J. Effects of glucose metabolism pathways on sperm motility and oxidative status during long-term liquid storage of goat semen. Theriogenology 2016, 86, 839–849. [Google Scholar] [CrossRef]
- Gadea, J.; Sellés, E.; Marco, M.A.; Coy, P.; Matás, C.; Romar, R.; Ruiz, S. Decrease in glutathione content in boar sperm after cryopreservation: Effect of the addition of reduced glutathione to the freezing and thawing extenders. Theriogenology 2004, 62, 690–701. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, R.; Li, W.M.; Niu, Y.J.; Guo, H.C.; Liu, X.H.; Zhao, L.J. The protective effect of grape seed procyanidin extract against cadmium-induced renal oxidative damage in mice. Environ. Toxicol. Pharmacol. 2013, 36, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.; Szkutnik, D.; Sanocka, D.; Kurpisz, M. peroxidation components of sperm lipid membranes in male infertility. Ginekol. Pol. 2001, 72, 73–79. [Google Scholar] [PubMed]
- Wood, J.E.; Senthilmohan, S.T.; Peskin, A.V. Antioxidant activity of procyanidin-containing plant extracts at different phs. Food Chem. 2002, 77, 155–161. [Google Scholar] [CrossRef]
- Fracassetti, D.; Costa, C.; Moulay, L.; Tomás-Barberán, F.A. Ellagic acid derivatives, ellagitannins, proanthocyanidins and other phenolics, vitamin c and antioxidant capacity of two powder products from camu-camu fruit (myrciaria dubia). Food Chem. 2013, 139, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Bagchi, M.; Stohs, S.J.; Das, D.K.; Ray, S.D.; Kuszynski, C.A.; Joshi, S.S.; Pruess, H.G. Free radicals and grape seed proanthocyanidin extract: Importance in human health and disease prevention. Toxicology 2000, 148, 187–197. [Google Scholar] [CrossRef]
- Okazaki, T.; Mihara, T.; Fujita, Y.; Yoshida, S.; Teshima, H.; Shimada, M. Polymyxin b neutralizes bacteria-released endotoxin and improves the quality of boar sperm during liquid storage and cryopreservation. Theriogenology 2010, 74, 1691–1700. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (h) | GSPE (mg/L) | ||||
|---|---|---|---|---|---|
| 0 | 10 | 30 | 50 | 70 | |
| 0 | 85.29 ± 1.22 | 84.79 ± 1.42 | 85.00 ± 1.31 | 84.65 ± 1.23 | 84.50 ± 1.32 |
| 24 | 78.31 ± 1.31 | 78.60 ± 1.19 | 78.98 ± 1.40 | 78.29 ± 1.33 | 78.94 ± 1.60 |
| 48 | 70.12 ± 1.44 | 70.13 ± 1.53 | 70.73 ± 0.69 | 70.51 ± 1.55 | 70.45 ± 1.12 |
| 72 | 66.80 ± 1.62 c | 68.59 ± 1.12 b | 69.45 ± 0.77 a | 68.31 ± 1.04 b | 66.21 ± 1.34 c |
| 96 | 54.30 ± 1.29 e | 58.32 ± 1.23 c | 65.34 ± 1.04 a | 61.58 ± 1.17 b | 56.44 ± 1.69 d |
| 120 | 45.32 ± 1.11 d | 46.89 ± 1.64 c | 51.46 ± 0.97 a | 49.41 ± 1.20 b | 45.77 ± 1.31 d |
| Time (h) | GSPE (mg/L) | ||||
|---|---|---|---|---|---|
| 0 | 10 | 30 | 50 | 70 | |
| 0 | 74.32 ± 1.27 | 74.32 ± 0.79 | 75.54 ± 1.31 | 74.67 ± 1.25 | 74.51 ± 1.77 |
| 24 | 70.69 ± 1.63 c | 71.84 ± 1.55 c | 73.82 ± 1.28 a | 72.71 ± 1.98 b | 71.43 ± 1.11 c |
| 48 | 64.25 ± 1.42 e | 77.72 ± 1.03 a | 70.43 ± 0.77 b | 68.08 ± 1.23 c | 67.01 ± 1.03 d |
| 72 | 60.20 ± 1.51 d | 65.11 ± 1.01 b | 67.13 ± 1.51 a | 65.85 ± 1.01 ab | 63.02 ± 1.20 c |
| 96 | 53.05 ± 0.88 e | 56.31 ± 1.22 b | 58.74 ± 1.01 a | 56.88 ± 1.44 b | 55.71 ± 1.25 b |
| 120 | 44.32 ± 1.16 | 47.43 ± 1.13 c | 50.97 ± 0.75 a | 48.42 ± 1.02 b | 46.33 ± 1.34 c |
| Time (h) | GSPE (mg/L) | ||||
|---|---|---|---|---|---|
| 0 | 10 | 30 | 50 | 70 | |
| 0 | 85.26 ± 1.35 | 84.83 ± 1.34 | 85.16 ± 1.42 | 85.34 ± 1.24 | 84.76 ± 1.43 |
| 24 | 81.85 ± 1.61 b | 81.73 ± 1.22 b | 83.19 ± 1.33 a | 82.54 ± 1.63 ab | 80.55 ± 1.21 c |
| 48 | 78.94 ± 1.48 c | 79.45 ± 1.12 bc | 81.49 ± 1.41 a | 80.79 ± 1.52 ab | 79.43 ± 1.22 bc |
| 72 | 72.50 ± 1.62 d | 75.13 ± 1.32 c | 78.46 ± 1.04 a | 76.11 ± 1.02 b | 74.40 ± 1.04 c |
| 96 | 65.69 ± 1.31 e | 69.45 ± 0.88 c | 74.00 ± 1.11 a | 70.33 ± 1.17 b | 67.72 ± 1.31 d |
| 120 | 62.05 ± 1.22 c | 65.02 ± 1.26 b | 67.55 ± 0.65 a | 65.33 ± 0.51 b | 64.75 ± 1.06 b |
| Time (h) | GSPE (mg/L) | ||||
|---|---|---|---|---|---|
| 0 | 10 | 30 | 50 | 70 | |
| 0 | 87.60 ± 1.23 | 87.27 ± 1.14 | 87.24 ± 1.51 | 87.34 ± 1.23 | 87.53 ± 1.86 |
| 24 | 77.11 ± 1.31 | 78.80 ± 1.13 bc | 81.12 ± 1.23 a | 80.12 ± 1.21 ab | 78.34 ± 1.64 bc |
| 72 | 62.79 ± 1.45 c | 64.99 ± 1.01 b | 70.36 ± 1.02 a | 65.20 ± 1.32 b | 63.79 ± 1.11 c |
| 120 | 43.19 ± 1.44 d | 47.37 ± 1.04 bc | 50.78 ± 1.13 a | 48.03 ± 0.62 b | 46.72 ± 1.20 c |
| Groups | Hybridization Number | Conception Number | Conception Rate (%) | Litter Sizes |
|---|---|---|---|---|
| Control | 83 | 57 | 68.7% | 1.47 ± 0.21 b |
| 30 Mg/L GSPE Group | 80 | 58 | 72.5% | 1.50 ± 0.30 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, F.; Li, Y.; Feng, T.; Du, Y.; Ren, F.; Zhang, L.; Han, N.; Ma, S.; Li, F.; Wang, P.; et al. Grape Seed Procyanidin Extract (GSPE) Improves Goat Sperm Quality When Preserved at 4 °C. Animals 2019, 9, 810. https://doi.org/10.3390/ani9100810
Wen F, Li Y, Feng T, Du Y, Ren F, Zhang L, Han N, Ma S, Li F, Wang P, et al. Grape Seed Procyanidin Extract (GSPE) Improves Goat Sperm Quality When Preserved at 4 °C. Animals. 2019; 9(10):810. https://doi.org/10.3390/ani9100810
Chicago/Turabian StyleWen, Fei, Yu Li, Tianyu Feng, Yeqing Du, Fa Ren, Likun Zhang, Ning Han, Shulan Ma, Fangzhou Li, Peng Wang, and et al. 2019. "Grape Seed Procyanidin Extract (GSPE) Improves Goat Sperm Quality When Preserved at 4 °C" Animals 9, no. 10: 810. https://doi.org/10.3390/ani9100810
APA StyleWen, F., Li, Y., Feng, T., Du, Y., Ren, F., Zhang, L., Han, N., Ma, S., Li, F., Wang, P., & Hu, J. (2019). Grape Seed Procyanidin Extract (GSPE) Improves Goat Sperm Quality When Preserved at 4 °C. Animals, 9(10), 810. https://doi.org/10.3390/ani9100810