Protective Effect of Resveratrol Improves Systemic Inflammation Responses in LPS-Injected Lambs


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment 1
2.2. Experiment 2
2.3. Enzyme Linked Immunosorbent Assays
2.4. Quantitative Real-Time PCR
2.5. Statistical Analysis
3. Results
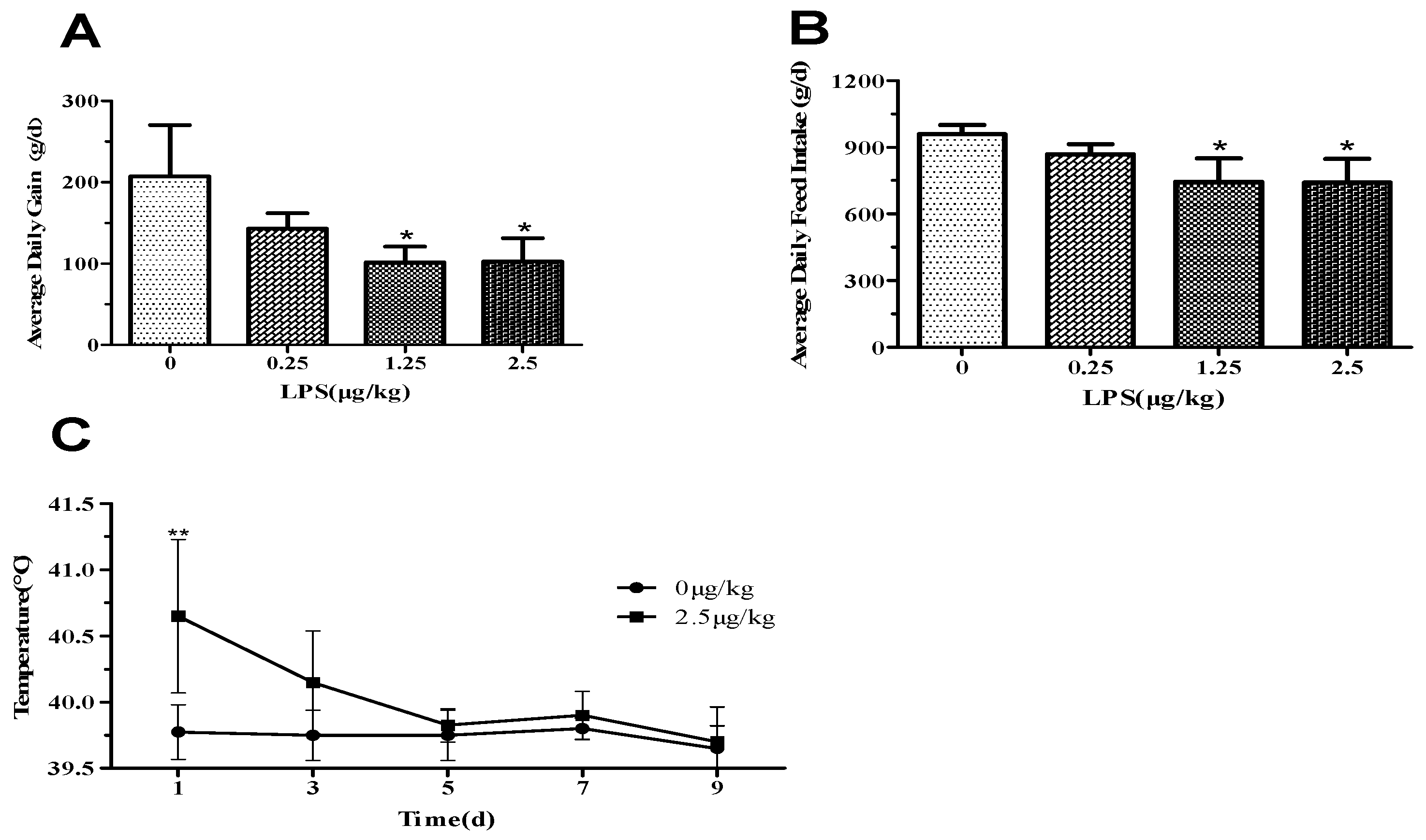
3.1. Average Daily Gain, Feed Intake, and Clinical Symptoms after LPS-Challenge
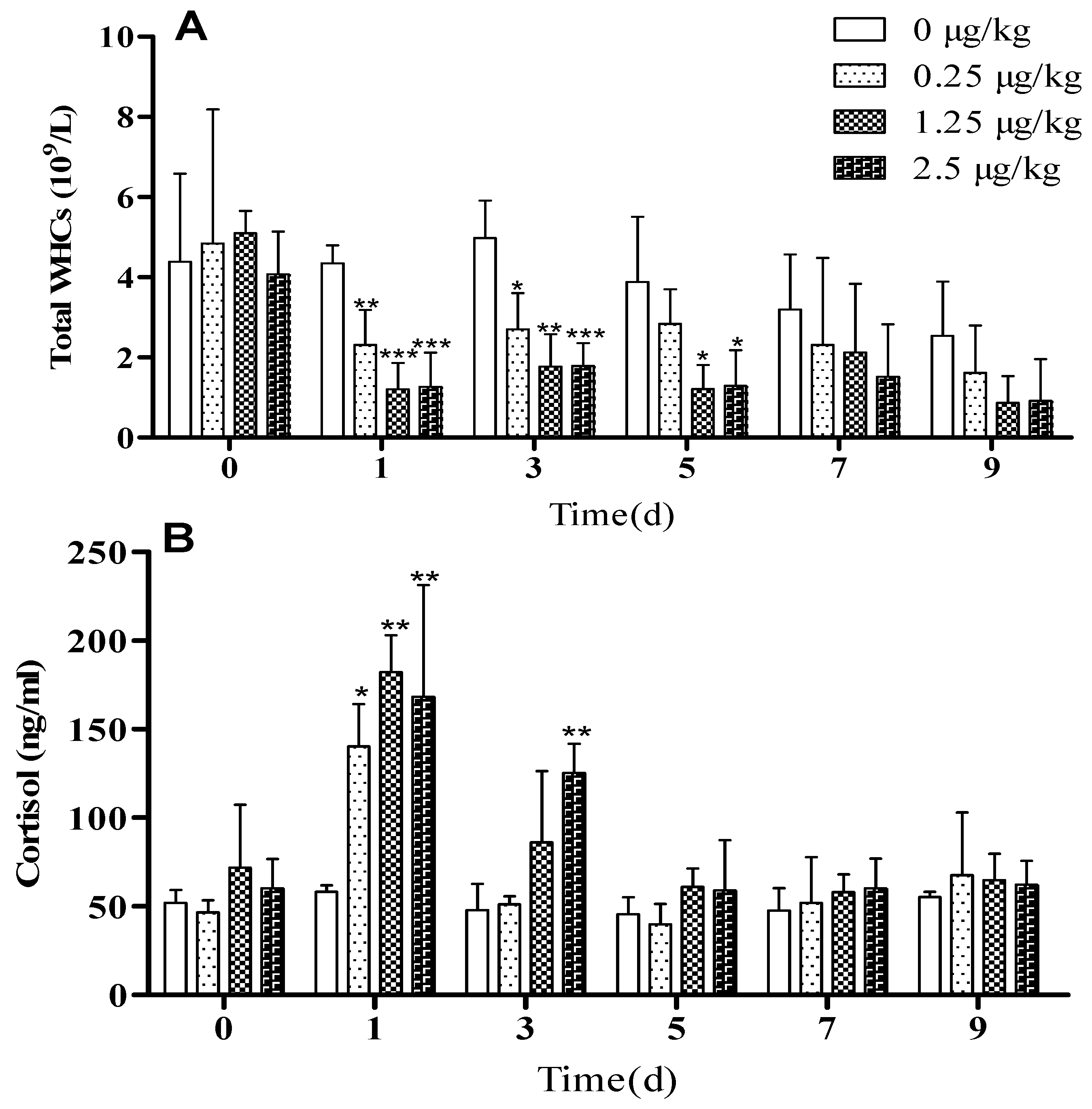
3.2. LPS Challenge Induced Systemic Inflammation in Lambs
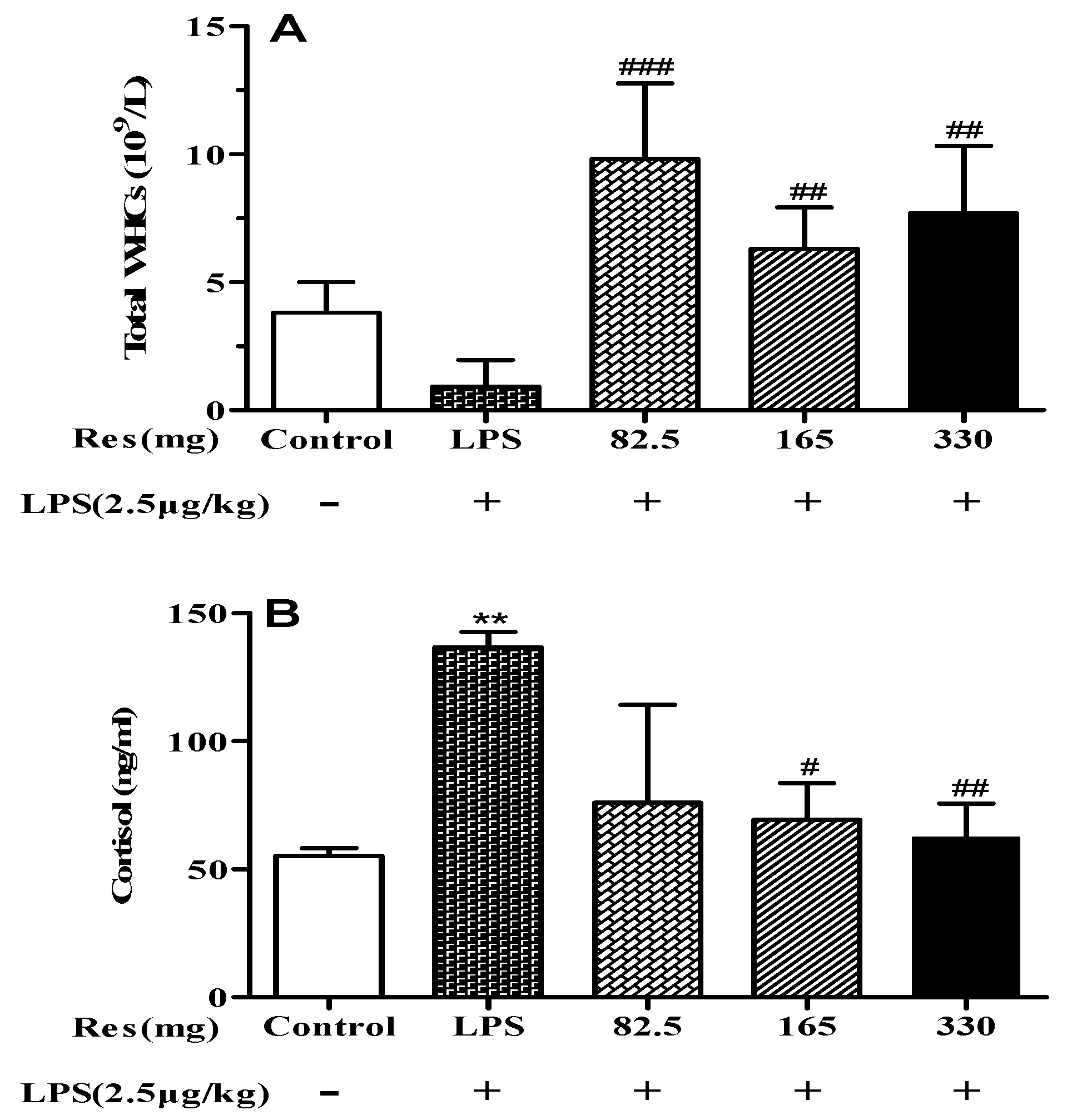
3.3. Effect of Resveratrol on Blood Parameters in LPS-Challenged Lambs
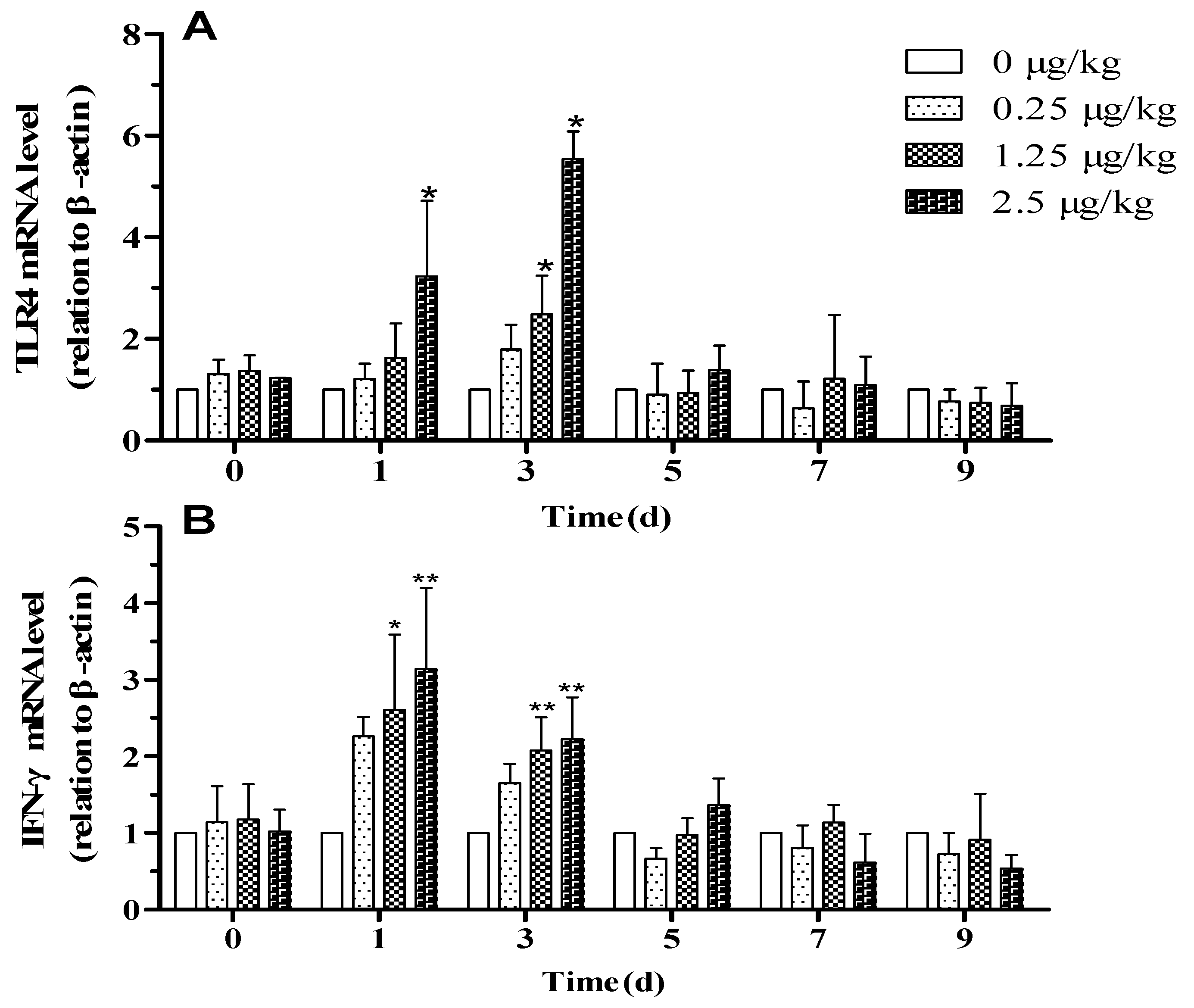
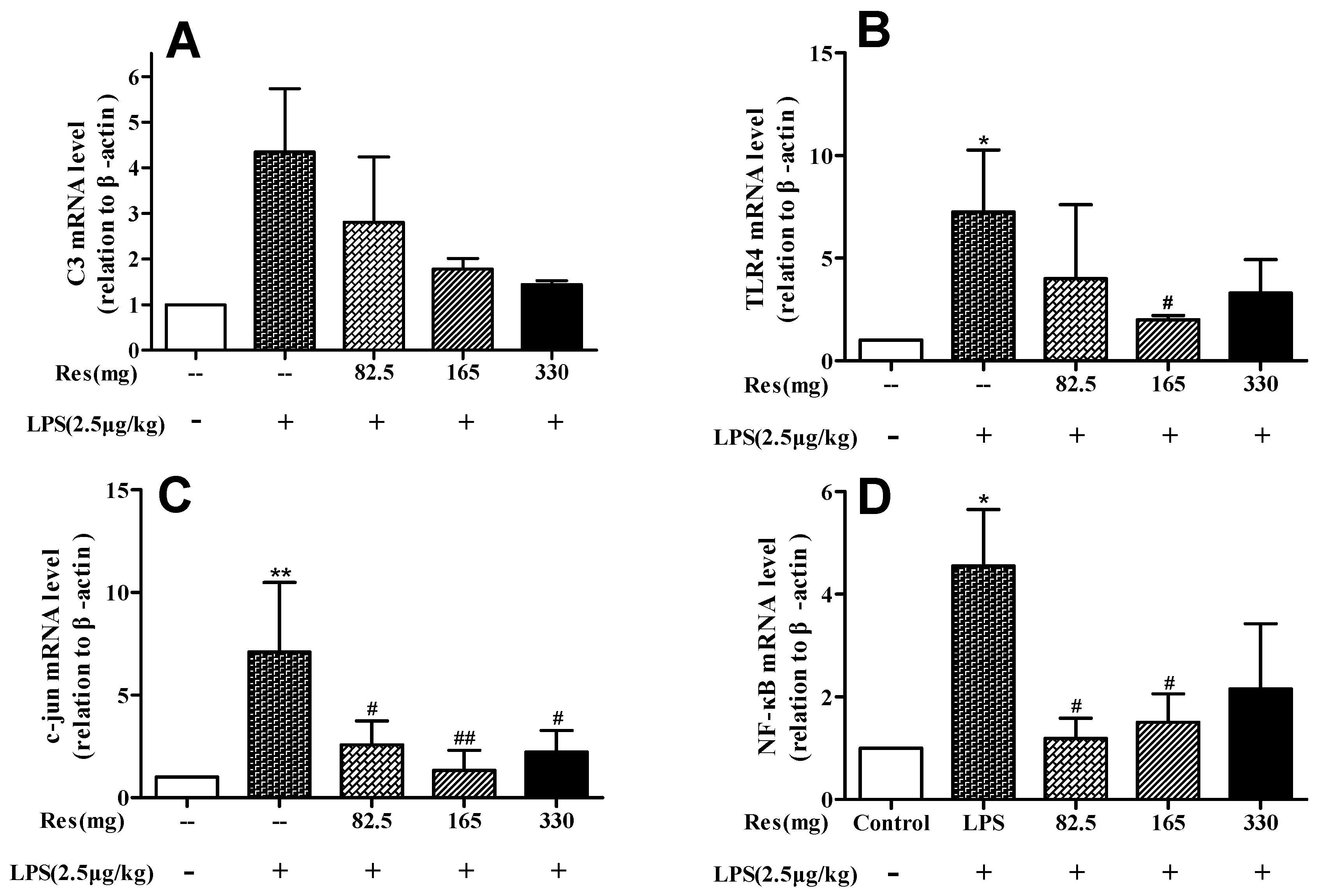
3.4. Effect of Resveratrol on Expression of Genes Following the LPS Challenge
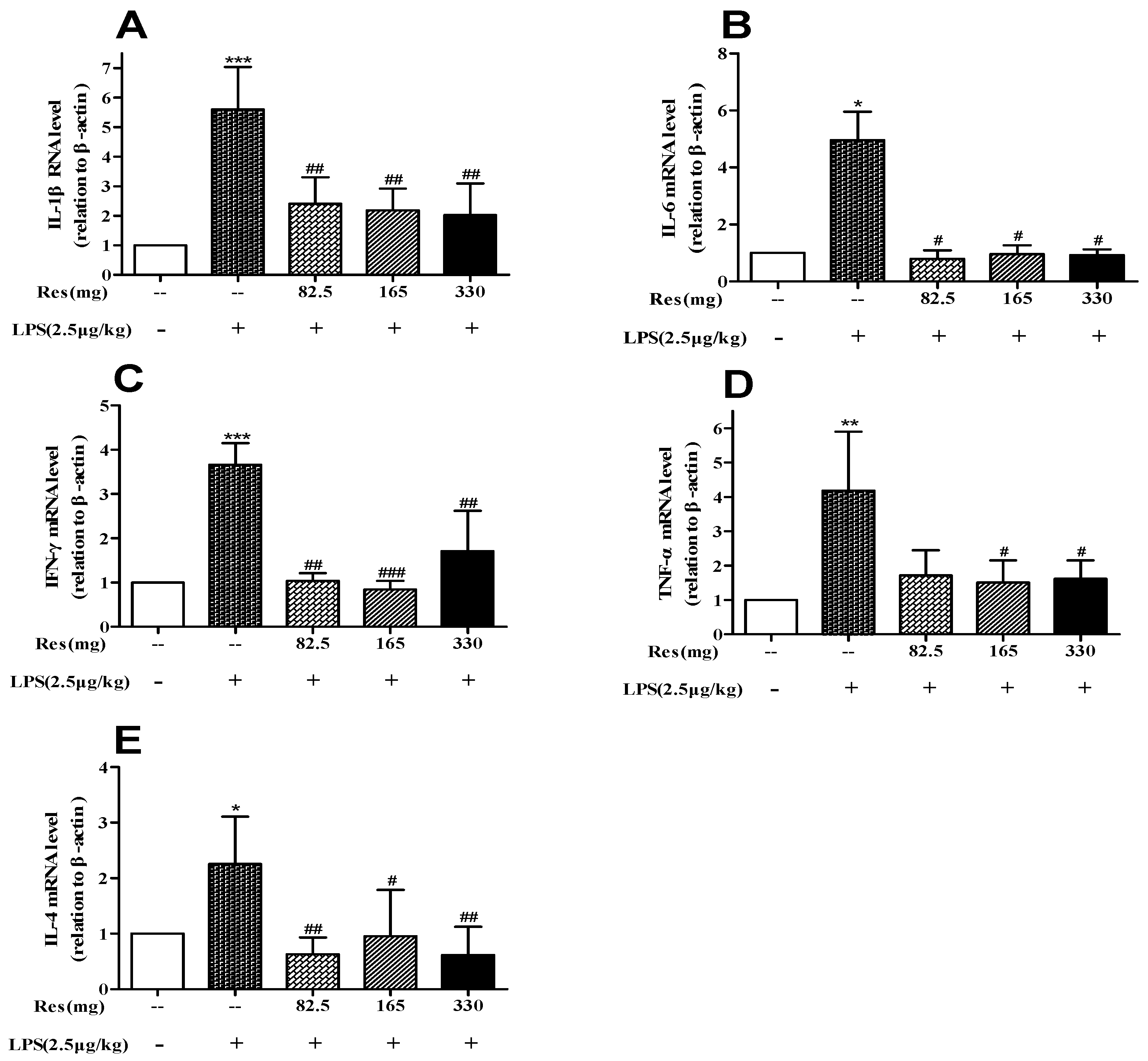
3.5. Effect of Resveratrol on Expression of Inflammatory Cytokines Following LPS Challenge
4. Discussion
4.1. LPS Challenge Induced Systemic Inflammation in Lambs
4.2. Resveratrol Improves Systemic Inflammation Response in LPS-Injected Lambs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kumar, B. Stress and its impact on farm animals. Front. Biosci. 2012, 4, 1759–1767. [Google Scholar] [CrossRef]
- Hadfield, J.M.; Bowdridge, E.C.; Holásková, I.; Elsasser, T.H.; Dailey, R.A. Breed-specific differences in the immune response to lipopolysaccharide in ewes. J. Anim. Sci. 2018, 96, 4220–4228. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, A.; Korkmaz, F.; Elsasser, T.; Kerr, D. Neonatal lipopolysaccharide exposure does not diminish the innate immune response to a subsequent lipopolysaccharide challenge in Holstein bull calves. J. Dairy Sci. 2016, 99, 5750–5763. [Google Scholar] [CrossRef] [PubMed]
- Campos, P.H.; Merlot, E.; Damon, M.; Noblet, J.; Le Floc’h, N. High ambient temperature alleviates the inflammatory response and growth depression in pigs challenged with Escherichia coli lipopolysaccharide. Vet. J. 2014, 200, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Kallapura, G.; Pumford, N.; Hernandez-Velasco, X.; Hargisand, B.M.; Tellez, G. Mechanisms involved in lipopolysaccharide derived ROS and RNS oxidative stress and septic shock. J. Microbiol. Res. Rev. 2014, 2, 6–11. [Google Scholar]
- Plessers, E.; Watteyn, A.; Wyns, H.; Pardon, B.; De Backer, P.; Croubels, S. Study of the immunomodulatory properties of gamithromycin and dexamethasone in a lipopolysaccharide inflammation model in calves. Res. Vet. Sci. 2015, 103, 218–223. [Google Scholar] [CrossRef]
- Harris, R.E. Inflammation in the Pathogenesis of Chronic Diseases: The Cox-2 Controversy; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Singh, A.P.; Singh, R.; Verma, S.S.; Rai, V.; Kaschula, C.H.; Maiti, P.; Gupta, S.C. Health benefits of resveratrol: Evidence from clinical studies. Med. Res. Rev. 2019, 39, 1851–1891. [Google Scholar] [CrossRef]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The therapeutic potential of resveratrol: A review of clinical trials. NPJ Precis. Oncol. 2017, 1, 35. [Google Scholar] [CrossRef]
- Bobermin, L.D.; Roppa, R.H.A.; Quincozes-Santos, A. Adenosine receptors as a new target for resveratrol-mediated glioprotection. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 634–647. [Google Scholar] [CrossRef]
- Bigagli, E.; Cinci, L.; Paccosi, S.; Parenti, A.; D’Ambrosio, M.; Luceri, C. Nutritionally relevant concentrations of resveratrol and hydroxytyrosol mitigate oxidative burst of human granulocytes and monocytes and the production of pro-inflammatory mediators in LPS-stimulated RAW 264.7 macrophages. Int. Immunopharmacol. 2017, 43, 147–155. [Google Scholar] [CrossRef]
- Mathew, L.M.; Woode, R.A.; Axiak-Bechtel, S.M.; Amorim, J.R.; DeClue, A.E. Resveratrol administration increases phagocytosis, decreases oxidative burst, and promotes pro-inflammatory cytokine production in healthy dogs. Vet. Immunol. Immunopathol. 2018, 203, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Li, Y.; Cao, L.; Du, J.; Zheng, T.; Qian, H.; Gu, Z.; Jeney, G.; Xu, P.; Yin, G. Antioxidative, anti-inflammatory and hepatoprotective effects of resveratrol on oxidative stress-induced liver damage in tilapia (Oreochromis niloticus). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 215, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.K.; Ma, Q.; Lei, Z.; Sha, H. Resveratrol ameliorates the deleterious effect of severe acute pancreatitis. Cell Biochem. Biophys. 2012, 62, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Ma, J.; Wang, W.; Zhang, L.; Xu, J.; Wang, K.; Li, D. Resveratrol supplement inhibited the NF-κB inflammation pathway through activating AMPKα-SIRT1 pathway in mice with fatty liver. Mol. Cell. Biochem. 2016, 422, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Muñoz, M.; Fischer, A.; Plickert, R.; Haag, L.M.; Otto, B.; Kühl, A.A.; Loddenkemper, C.; Göbel, U.B.; Heimesaat, M.M. Anti-Inflammatory Effects of Resveratrol, Curcumin and Simvastatin in Acute Small Intestinal Inflammation. PLoS ONE 2010, 5, e15099. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cui, H.; Niu, F.; Liu, S.L.; Li, Y.; Zhang, L.M.; Du, H.B.; Zhao, Z.G.; Niu, C.Y. Effect of Resveratrol on Blood Rheological Properties in LPS-Challenged Rats. Front. Physiol. 2018, 9, 1202. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, E.J.; Mattson, M.P.; Calabrese, V. Resveratrol commonly displays hormesis: Occurrence and biomedical significance. Hum. Exp. Toxicol. 2010, 29, 980–1015. [Google Scholar] [CrossRef]
- Salvesen, Ø.; Reiten, M.R.; Espenes, A.; Bakkebø, M.K.; Tranulis, M.A.; Ersdal, C. LPS-induced systemic inflammation reveals an immunomodulatory role for the prion protein at the blood-brain interface. J. Neuroinflamm. 2017, 14, 106. [Google Scholar] [CrossRef] [Green Version]
- Gupta, Y.; Briyal, S.; Chaudhary, G. Protective effect of trans-resveratrol against kainic acid-induced seizures and oxidative stress in rats. Pharmacol. Biochem. Behav. 2002, 71, 245–249. [Google Scholar] [CrossRef]
- Narayanan, B.A.; Narayanan, N.K.; Nargi, D. Natural anti-inflammatory agent resveratrol inhibit mediators of inflammation and cancer. Proc. Am. Assoc. Cancer Res. 2006, 47, 920–921. [Google Scholar]
- Shen, M.Y.; Hsiao, G.; Liu, C.L.; Fong, T.H.; Lin, K.H.; Chou, D.S.; Sheu, J.R. Inhibitory mechanisms of resveratrol in platelet activation: Pivotal roles of p38 MAPK and NO/cyclic GMP. Br. J. Haematol. 2007, 139, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Olas, B.; Wachowicz, B. Resveratrol, a phenolic antioxidant with effects on blood platelet functions. Platelets 2005, 16, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Sha, H.; Ma, Q.; Jha, R.K.; Wu, Z.; Qingyuan, Z.; Wang, Z.; Ma, Z.; Luo, X.; Liu, C. Resveratrol suppresses microcirculatory disturbance in a rat model of severe acute pancreatitis. Cell Biophys. 2013, 67, 1059–1065. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Qian, L.H.; Deng, B.; Liu, Z.M.; Zhao, Y.; Le, Y.Y. Resveratrol protects vascular endothelial cells from high glucose–Induced apoptosis through Inhibition of NADPH oxidase activation–driven oxidative stress. CNS Neurosci. Ther. 2013, 19, 675–681. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Bandyopadhyay, M. Neuroprotective mode of action of resveratrol in central nervous system. Pharma. Nutrition 2013, 1, 90–97. [Google Scholar] [CrossRef]
- Shah, Z.; Ali, T. Inflammation the kingpin of chronic diseases naturopathy as an alternative source of treatment: An updated overview. J. Drug Deliv. Ther. 2019, 9, 478–483. [Google Scholar] [CrossRef]
- Yates, D.T.; Loest, C.A.; Ross, T.T.; Hallford, D.M.; Carter, B.H.; Limesand, S.W. Effects of bacterial lipopolysaccharide injection on white blood cell counts, hematological variables, and serum glucose, insulin, and cortisol concentrations in ewes fed low- or high-protein diets. J. Anim. Sci. 2011, 89, 4286–4293. [Google Scholar] [CrossRef]
- Beishuizen, A.; Thijs, L.G. Endotoxin and the hypothalamo-pituitary-adrenal (HPA) axis. J. Endotoxin Res. 2003, 9, 3–24. [Google Scholar]
- Wu, C.C.; Huang, Y.S.; Chen, J.S.; Huang, C.F.; Su, S.L.; Lu, K.C.; Lin, Y.F.; Chu, P.; Lin, S.H.; Sytwu, H.K. Resveratrol Ameliorates renal damage, increases expression of heme oxygenase-1, and has anti-complement, anti-oxidative, and anti-apoptotic effects in a murine model of membranous nephropathy. PLoS ONE 2015, 10, e0125726. [Google Scholar] [CrossRef]
- Elvington, M.; Liszewski, M.K.; Bertram, P.; Kulkarni, H.S.; Atkinson, J.P. A C3(H2O) recycling pathway is a component of the intracellular complement system. J. Clin. Investig. 2017, 127, 970–981. [Google Scholar] [CrossRef]
- Chen, M.; Wang, C.; Li, Z.; Chen, J. Involvement of JNK signaling pathway in lipopolysaccharide-induced complement C3 transcriptional activation from amphioxus Branchiostoma belcheri. Fish Shellfish. Immunol. 2019, 86, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Karin, M. The Regulation of AP-1 Activity by mitogen-activated protein kinases. J. Biol. Chem. 1995, 270, 16483–16486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Xiao, C.; Wei, Z.; Wang, J.; Yang, Z.; Fu, Y. Resveratrol inhibits LPS-induced mice mastitis through attenuating the MAPK and NF-κB signaling pathway. Microb. Pathog. 2017, 107, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Hu, Z.; Fu, Q.; Song, X.; Cui, Q.; Jia, R.; Zou, Y.; He, C.; Li, L.; Yin, Z. Resveratrol mitigates lipopolysaccharide-mediated acute inflammation in rats by inhibiting the TLR4/NF-κBp65/MAPKs signaling cascade. Sci. Rep. 2017, 7, 45006. [Google Scholar] [CrossRef] [PubMed]
- Moynagh, P.N. The NF-kappaB pathway. J. Cell Sci. 2005, 118, 4589–4592. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2. [Google Scholar] [CrossRef]
- Gorina, R.; Font-Nieves, M.; Márquez-Kisinousky, L.; Santalucia, T.; Planas, A.M. Astrocyte TLR4 activation induces a proinflammatory environment through the interplay between MyD88-dependent NFκB signaling, MAPK, and Jak1/Stat1 pathways. Glia 2011, 59, 242–255. [Google Scholar] [CrossRef]
- Deng, Y.; Gong, W.; Li, Q.; Wu, X.; Wu, L.; Zheng, X.; Chen, W.; Huang, H. Resveratrol inhibits high glucose-induced activation of AP-1 and NF-κB via SphK1/S1P2 pathway to attenuate mesangial cells proliferation and inflammation. J. Funct. Foods 2019, 55, 86–94. [Google Scholar] [CrossRef]
- Nielsen, O.H.; Køppen, T.; Rüdiger, N.; Horn, T.; Eriksen, J.; Kirman, I.; Nielsen, D.O.H. Involvement of interleukin-4 and-10 in inflammatory bowel disease. Dig. Dis. Sci. 1996, 41, 1786–1793. [Google Scholar] [CrossRef]
- Laflamme, N.; Lacroix, S.; Rivest, S. An essential role of interleukin-1beta in mediating NF-kappaB activity and COX-2 transcription in cells of the blood-brain barrier in response to a systemic and localized inflammation but not during endotoxemia. J. Neurosci. 1999, 19, 10923–10930. [Google Scholar] [CrossRef]
- Barnes, T.C.; Anderson, M.E.; Moots, R.J. The Many Faces of Interleukin-6: The Role of IL-6 in Inflammation, Vasculopathy, and Fibrosis in Systemic Sclerosis. Int. J. Rheumatol. 2011, 2011, 721608. [Google Scholar] [CrossRef]
- Zhang, J. Yin and yang interplay of IFN-gamma in inflammation and autoimmune disease. J. Clin. Investig. 2007, 117, 871–873. [Google Scholar] [CrossRef]
- Kim, K.S.; Jung, H.; Shin, I.K.; Choi, B.R.; Kim, D.H. Induction of interleukin-1 beta (IL-1β) is a critical component of lung inflammation during influenza A (H1N1) virus infection. J. Med. Virol. 2015, 87, 1104–1112. [Google Scholar] [CrossRef]
- Rubinstein, A.I.; Rubinstein, D.B. Interleukin-4 (IL-4) inhibits secretion of IL-1 beta, tumor necrosis factor aipha, and IL-6 by human monocytes. Blood 1990, 76, 1392–1397. [Google Scholar] [CrossRef]
- Paul, W.E. Interleukin-4 a prototypic immunoregulatory lymphokine. Blood 1991, 77, 1859–1870. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Nutrient 2 | ||
|---|---|---|---|
| Ingredient, % of DM | |||
| Oat hay | 25.00 | ME (MJ/kg) | 9.50 |
| Corn stalk | 15.00 | Crude protein (%) | 10.89 |
| Corn | 31.50 | Neutral detergent fiber (%) | 43.83 |
| Soybean meal | 4.00 | Acid detergent fiber (%) | 16.71 |
| Wheat bran | 4.50 | Ca (%) | 0.63 |
| DDGS | 3.00 | P (%) | 0.45 |
| Molasses | 3.00 | ||
| Barley grain | 8.50 | ||
| NaCl | 0.50 | ||
| CaHPO4 | 0.80 | ||
| Limestone | 0.70 | ||
| Soybean oil | 1.50 | ||
| Sodium bicarbonate | 1.00 | ||
| Premix 1 | 1.00 |
| Gene | Forward Primer 5’-3’ | Reverse Primer 5’-3’ | Product Length | Annealing Temperature (℃) |
|---|---|---|---|---|
| C3 | GCACTGTCCACCAACCTCA | ATCAGGCTTCTGCTTCTCCA | 87 | 58 |
| TLR4 | GGCATCATCTTCATCGTCCT | CCACTCCAGGTAGGTGTTCC | 99 | 58 |
| NF-κB | CATCAGCCAGCGCATCCAGAC | GCACGGCATTCAGGTCGTAGTC | 86 | 61 |
| c-jun | AGCGGATCAAGGCGGAGAGG | CCTGAGCATATTGGCGGTGGAC | 155 | 55 |
| IL-1β | GCAGGCAGTGTCGGTCATCG | CCTCAGGTCATCATCACGGAAGAC | 82 | 58 |
| IL-4 | GCGGACTTGACAGGAATCTCAGC | CAGCGTACTTGTACTCGTCTTGGC | 80 | 63 |
| IL-6 | ACACTGACATGCTGGAGAAGATGC | GCCGCAGCTACTTCATCCGAATAG | 132 | 61 |
| IFN-R | ATGTTTCATTTGCCACCATCC | GGTTACGCTTGCTTTGCCTTATGT | 81 | 60 |
| TNF-α | CTGGCGGAGGAGGTGCTCTC | GGAGGAAGGAGAAGAGGCTGAGG | 85 | 59 |
| β-actin | AGCCTTCCTTCCTGGGCATGGA | GGACAGCACCGTGTTGGCGTAGA | 113 | 60 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, Y.; Zhou, J.; Ji, K.; Liu, H.; Degen, A.; Zhai, M.; Jiao, D.; Guo, J.; Zhao, Z.; Yang, G. Protective Effect of Resveratrol Improves Systemic Inflammation Responses in LPS-Injected Lambs. Animals 2019, 9, 872. https://doi.org/10.3390/ani9110872
Liang Y, Zhou J, Ji K, Liu H, Degen A, Zhai M, Jiao D, Guo J, Zhao Z, Yang G. Protective Effect of Resveratrol Improves Systemic Inflammation Responses in LPS-Injected Lambs. Animals. 2019; 9(11):872. https://doi.org/10.3390/ani9110872
Chicago/Turabian StyleLiang, Yanping, Jianwei Zhou, Kaixi Ji, Hu Liu, Allan Degen, Manjun Zhai, Dan Jiao, Junqiang Guo, Zongsheng Zhao, and Guo Yang. 2019. "Protective Effect of Resveratrol Improves Systemic Inflammation Responses in LPS-Injected Lambs" Animals 9, no. 11: 872. https://doi.org/10.3390/ani9110872
APA StyleLiang, Y., Zhou, J., Ji, K., Liu, H., Degen, A., Zhai, M., Jiao, D., Guo, J., Zhao, Z., & Yang, G. (2019). Protective Effect of Resveratrol Improves Systemic Inflammation Responses in LPS-Injected Lambs. Animals, 9(11), 872. https://doi.org/10.3390/ani9110872