Characteristics of the BMP7 Promoter in Hu Sheep

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Ethics Statement
2.2. Sample Collection
2.3. Bioinformatics Analysis of Proximal Promoter
2.4. Deletion Plasmids Construction
2.5. Transfection and Dual-Luciferase Assay
2.6. Mutant and Overexpression Plasmids Construction and Dual-Luciferase Assays
2.7. Statistical Analysis
3. Results
3.1. Bioinformatics Analysis of the Proximal Promoter Region of Hu Sheep BMP7
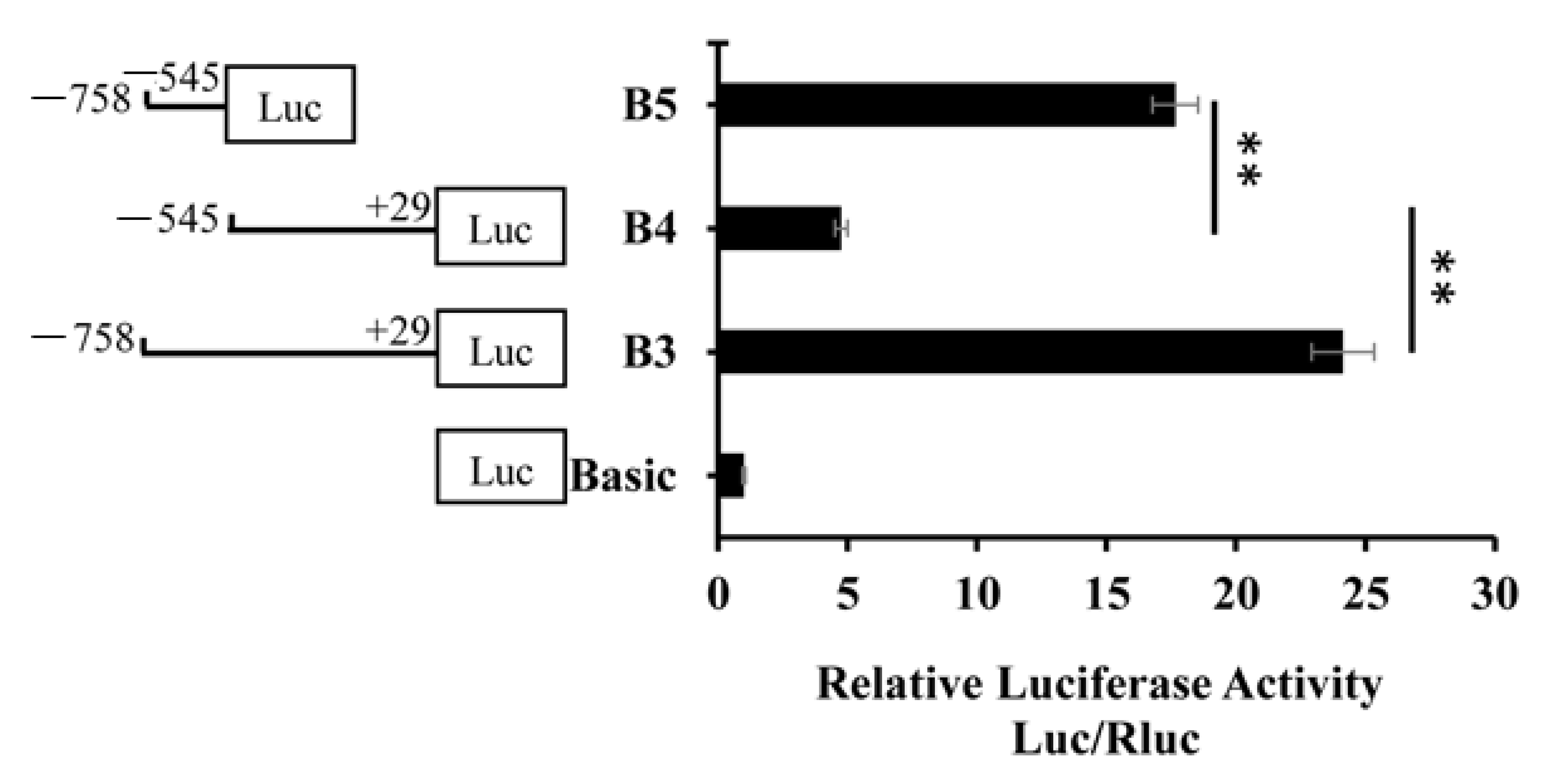
3.2. Deletion Fragment Analysis of the BMP7 Promoter
3.3. Conservative Analysis of Transcription Core and Prediction of Transcription Factors
3.4. Verification of Point Mutation of Transcription Factor
3.5. Overexpression of EGR1 on Transcriptional Activity of the BMP7 Promoter
4. Discussion
4.1. Characteristics of the Hu Sheep BMP7 Promoter
4.2. The Regulatory Elements Involved in Control of Hu Sheep BMP7 Expression
4.3. Functions of EGR1 and SP1
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Termaat, M.F.; Den, B.F.C.; Bakker, F.C.; Bakker, F.C.; Patka, P.; Haarman, H.J. Bone morphogenetic proteins. Development and clinical efficacy in the treatment of fractures and bone defects. J. Bone Jt. Surg. 2005, 87, 1367–1378. [Google Scholar]
- Oshinmori, N.; Fuchs, E. Paracrine TGF-β signaling counterbalances BMP-mediated repression in hair follicle stem cell activation. Cell Stem Cell 2012, 10, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Genander, M.; Cook, P.J.; Ramsköld, D.; Ramskold, D.; Keyes, B.E.; Mertz, A.F.; Sandberg, R.; Fuchs, E. BMP signaling and its pSMAD1/5 target genes differentially regulate hair follicle stem cell lineages. Cell Stem Cell 2014, 15, 619–633. [Google Scholar] [CrossRef] [PubMed]
- Kulessa, H.; Turk, G.; Hogan, B.L. Inhibition of bmp signaling affects growth and differentiation in the anagen hair follicle. EMBO J. 2000, 19, 6664–6674. [Google Scholar] [CrossRef] [PubMed]
- Andl, T.; Ahn, K.; Kairo, A.; Chu, E.Y.; Wine-Lee, L.; Reddy, S.T.; Croft, N.J.; Cebra-Thomas, J.A.; Metzger, D.; Chambon, P.; et al. Epithelial bmprla regulates differentiation and proliferation in postnatal hair follicles and is essential for tooth development. Development 2004, 131, 2257–2268. [Google Scholar] [CrossRef]
- Ma, Y.N.; You, Y.; Shen, H.H.; Sun, Z.Z.; Zeng, L.; Fa, Y.Z. Effect of Noggin silencing on the BMP and Wnt signaling pathways. Acta Lab. Anim. Sci. Sin. 2016, 24, 475–478. (In Chinese) [Google Scholar]
- Noramly, S.; Morgan, B.A. BMPs mediate lateral inhibition at successive stages in feather tract development. Development 1998, 125, 3775–3787. [Google Scholar]
- Song, L.L.; Cui, Y.; Yu, S.J.; Liu, P.G.; Zhang, Q. Expression characteristics of bmp2, bmpr-ia and noggin in different stages of hair follicle in yak skin. Gen. Comp. Endocrinol. 2017, 260, 18–24. [Google Scholar] [CrossRef]
- Adly, M.A.; Assaf, H.A.; Hussein, M.R. Expression of bone morphogenetic protein-7 in human scalp skin and hair follicles. Br. J. Dermatol. 2010, 154, 551–554. [Google Scholar] [CrossRef]
- Knudsen, S. Promoter2.0: for the recognition of PolII promoter sequences. Bioinformatics 1999, 5, 356–361. [Google Scholar] [CrossRef]
- Reese, M.G. Application of a time-delay neural network to promoter annotation in the Drosophila melanogaster genome. Comput. Chem. 2002, 1, 51–56. [Google Scholar]
- Li, L.C.; Dahiya, R. MethPrimer: designing primers for methylation PCRs. Bioinformatics 2002, 11, 1427–1431. [Google Scholar] [CrossRef] [PubMed]
- Wingender, E. The TRANSFAC project as an example of framework technology that supports the analysis of genomic regulation. Brief. Bioinform. 2008, 4, 326–332. [Google Scholar] [CrossRef] [PubMed]
- Aziz, K.; Oriol, F.; Arnaud, S.; Marius, G.; Jaime, A.C.; Robin, V.D.L.; Adrien, B.; Jeanne, C.; Shubhada, R.K.; Ge, T.; et al. JASPAR 2018: update of the open-access database of transcription factor binding profiles and its web framework. Nucleic Acids Res. 2018, D1, D1284. [Google Scholar]
- Lee, W.S.; Otsuka, F.; Moore, R.K.; Shimasaki, S. Effect of Bone Morphogenetic Protein-7 on Folliculogenesis and Ovulation in the Rat. Biol. Reprod. 2014, 65, 994–999. [Google Scholar] [CrossRef]
- Dudley, A.T.; LyonS, K.M.; Robertson, E.J. A requirement for bone morphongenetic protein-7 during development of the mammalian kidney and eye. Genes Dev. 1995, 9, 2795–2807. [Google Scholar] [CrossRef]
- Ross, A.; Munger, S.; Capel, B. Bmp7 Regulates Germ Cell Proliferation in Mouse Fetal Gonads. Sex. Dev. 2007, 1, 127. [Google Scholar] [CrossRef]
- Ma, Q.; Feng, W.R.; Zhuang, Z.M.; Liu, S.F. Cloning, expression profiling and promoter functional analysis of Bone morphogenetic protein 6 and 7 in tongue sole (Cynoglossus semilaevis). Fish Physiol. Biochem. 2017, 43, 435–454. [Google Scholar] [CrossRef]
- Kawai, S.; Sugiura, T. Characterization of Human Bone Morphogenetic Protein(BMP)-4 and -7 Gene Promoters: Activation of BMP Promoters by Gli, a Sonic Hedgehog Mediator. Bone 2001, 29, 54–61. [Google Scholar] [CrossRef]
- Simon, M.; Feliers, D.; Arar, M.; Bhandari, B.; Abboud, H.E. Cloning of the 5′-flanking region of the murine bone morphogenetic protein-7 gene. Mol. Cell. Biochem. 2002, 233, 31–37. [Google Scholar] [CrossRef]
- Li, M.; Chen, Q.X.; Sun, G.R.; Shi, X.M.; Zhao, Q.H.; Zhang, C.; Zhou, J.S.; Qin, N. Characterization and expression of bone morphogenetic protein 4 gene in postnatal pigs. Mol. Biol. Rep. 2010, 37, 2369–2377. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.P.; Wu, C.M.; Miao, J.H. Research Progress of Transcription Factors Sp1 and SP2. Int. J. Lab. Med. 2016, 37, 1094–1096. [Google Scholar]
- Ling, F.; Wang, T.; Wei, L.Q.; Zhu, X.P.; Chen, Y.S.; Li, J.Q.; Zhang, Z.W.; Du, H.L.; Wang, X.N.; Wang, J.F. Cloning and characterization of the 5′-flanking region of the pig AgRP gene. Mol. Boil. Rep. 2009, 381, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Sandelin, A.; Carninci, P.; Lenhard, B.; Ponjavic, J.; Hayashizaki, Y.; Hume, D.A. Mammalian RNA polymerase II core promoters: Insights from genome-wide studies. Nat. Rev. Genet. 2007, 8, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Qin, N. Polymorphism in the promoter region of swine BMP7 gene and their association with reproductive traits. Henan Agric. Univ. 2011. (In Chinese) [Google Scholar] [CrossRef]
- Li, F.; Jiang, Z.; Wang, K.; Guo, J.; Hu, G.; Sun, L.X.; Wang, T.; Tang, X.Z.; He, L.; Yao, J.C.; et al. Transactivation of the human NME5 gene by Sp1 in pancreatic cancer cells. Gene 2012, 503, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Shu, H.K.G. EGFR activation results in enhanced cyclooxygenase-2 expression through p38 mitogen-activated protein kinase-dependent activation of the Sp1/Sp3 transcription factors in buman gliomas. Cancer Res. 2007, 67, 6121–6129. [Google Scholar] [CrossRef]
- Nam, E.H.; Lee, Y.; Zhao, X.F.; Park, Y.K.; Lee, J.W.; Kim, S. ZEB2-Sp1 cooperation induces invasion by upregulating cadherin-11 and integrin α5 expression. Carcinogenesis 2014, 35, 302–314. [Google Scholar] [CrossRef]
- Bai, X.; Deng, H. Research progress on relationship between transcription factor Sp1 and tumor. J. Zhejiang Univ. (Med. Sci.) 2010, 39, 215–220. (In Chinese) [Google Scholar]
- Haugen, T.H.; Lace, M.J.; Ishiji, T.; Sameshima, A.; Anson, J.R.; Turek, L.P. Cellular factors are required to activate bovine papillomavirus-1 early gene transcription and to establish viral plasmid persistence but are not required for cellular transformation. Virology 2009, 389, 82–90. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Meng, A. Sp1-like transcription factors are regulators of embryonic development in vertebrates. Dev. Growth Regen. 2005, 47, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Wu, M.; Fang, F.; Tourtellotte, W.; Feghali-Bostwick, C.; Varga, J. Early growth response transcription factors: Key mediators of fibrosis and novel targets for anti-fibrotic therapy. Matrix Biol. 2011, 30, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Li, T.; Qureshi, H.Y.; Han, D.; Paudel, H.K. Early growth response 1 (Egr-1) regulates phosphorylation of microtubule-associated protein tau in mammalian brain. J. Biol. Chem. 2011, 286, 20569–20581. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.; Soung, N.K.; Thimmegowda, N.R.; Jeong, S.J.; Jang, J.H.; Moon, D.O.; Chung, J.K.; Lee, K.S.; Kwon, Y.T.; Erikson, R.L.; et al. Patulin induces colorectal cancer cells apoptosis through EGR-1 dependent ATF3 up-regulation. Cell. Signal. 2012, 24, 943–950. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.T.; Chen, C. Essential role of KLF5 transcription factor in cell proliferation and differentiation and its implications for human diseases. Cell. Mol. Life Sci. Cmls 2009, 66, 2691–2706. [Google Scholar] [CrossRef]
- Shindo, T.; Manabe, I.; Fukushima, Y.; Tobe, K.; Aizawa, K.; Miyamoto, S.; Kawai-Kowase, K.; Moriyama, N.; Imai, Y.; Kawakami, H.; et al. Kruppel-like zinc-finger transcription factor KLF5/BTEB2 is a target for angiotensin II signaling and and essential regulator of cardiovascular remodeling. Nat. Med. 2002, 8, 856–863. [Google Scholar] [CrossRef]
- Kaczynski, J.; Cook, T.; Urrutia, R. Sp1- and Kruppel-like transcription factors. Genome Biol. 2003, 4, 206. [Google Scholar] [CrossRef]
- Nandan, M.O.; Mcconnell, B.B.; Ghaleb, A.M.; Bialkowska, A.B.; Sheng, H.; Shao, J.; Babbin, B.A.; Robine, S.; Yang, V.W. Krüppel-like factor 5 mediates cellular transformation during oncogenic KRAS-induced intestinal tumorigenesis. Gastroenterology 2008, 134, 120–130. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) | Product Length/bp |
|---|---|---|
| B1 | F: GGGGTACCCCAAGTGGGCAACTCAGTGTC | 1757 |
| R: CCAAGCTTGTACAGGTCCAGCATGAACATGG | ||
| B2 | F: GGGGTACCCAGTACAAAGGCAAACTGGCAACA | 1014 |
| B3 | F: GGGGTACCCGCTCGTATTCCCCTCTCCGCATC | 787 |
| B4 | F: GGGGTACCCCCAGGCCCCAGCGCGTACCA | 574 |
| R: CCAAGCTTCCAGAGCGCCACGAAGCTGT | ||
| B5 | F: GGGGTACCCGAGTCCGGAGAAGGCAGG | 214 |
| R: CCAAGCTTGCTGGGGCCTGGGAGGAGGA |
| Name | Primer Sequence (5′–3′) | Length/bp |
|---|---|---|
| BTF1 | GCCCTTGGAAAGGCCGTCCTCTTCTCCCTCCTCCTCCCAG | 214 |
| BTR1 | CTGGGAGGAGGAGGGAGAAGAGGACGGCCTTTCCAAGGGC | |
| BTF2 | AGGCCGTCCTCCTCCCCCGCCGCCGCCCAGGCCCCAGC | 214 |
| BTR2 | GCTGGGGCCTGGGCGGCGGCGGGGGAGGAGGACGGCCT |
| Region | Transcription Factor | Start Site | Sequence |
|---|---|---|---|
| −758 bp to −545 bp | FOXB1 | −675 (1) | TCAGTAAATAT |
| FOXC1 | −675 (1) | TCAGTAAATAT | |
| FOXC2 | −675 (1) | TCAGTAAATATT | |
| FOXD2 | −672 (1) | GTAAATA | |
| FOXI1 | −672 (1) | GTAAATA | |
| FOXL1 | −672 (1) | GTAAATA | |
| FOXO4 | −672 (1) | GTAAATA | |
| Klf4 | −612 (1) | AGGGCGGGGC | |
| NRF1 | −552 (1) | CCGCGGGCGCG | |
| EGR1 | −552 (1) | CCTCCTCCCCCTCC | |
| KLF5 | −552 (1) | CCTCCTCCCC | |
| SP1 | −552 (1) | CCTCCTCCCCC | |
| EGR1 | −546 (1) | CCCCCTCCTCCTCC | |
| SP1 | −546 (1) | CCCCCTCCTCC | |
| SP2 | −546 (1) | CCCCCTCCTCCTCCC | |
| SP1 | −546 (1) | CCTCCTCCTCC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, X.; Sun, W.; Zou, S.; Chen, L.; Mwacharo, J.M.; Wang, J. Characteristics of the BMP7 Promoter in Hu Sheep. Animals 2019, 9, 874. https://doi.org/10.3390/ani9110874
Lv X, Sun W, Zou S, Chen L, Mwacharo JM, Wang J. Characteristics of the BMP7 Promoter in Hu Sheep. Animals. 2019; 9(11):874. https://doi.org/10.3390/ani9110874
Chicago/Turabian StyleLv, Xiaoyang, Wei Sun, Shuangxia Zou, Ling Chen, Joram M. Mwacharo, and Jinyu Wang. 2019. "Characteristics of the BMP7 Promoter in Hu Sheep" Animals 9, no. 11: 874. https://doi.org/10.3390/ani9110874
APA StyleLv, X., Sun, W., Zou, S., Chen, L., Mwacharo, J. M., & Wang, J. (2019). Characteristics of the BMP7 Promoter in Hu Sheep. Animals, 9(11), 874. https://doi.org/10.3390/ani9110874